Efficacy of insect larval meal to replace fish meal in juvenile barramundi, Lates calcarifer reared in freshwater
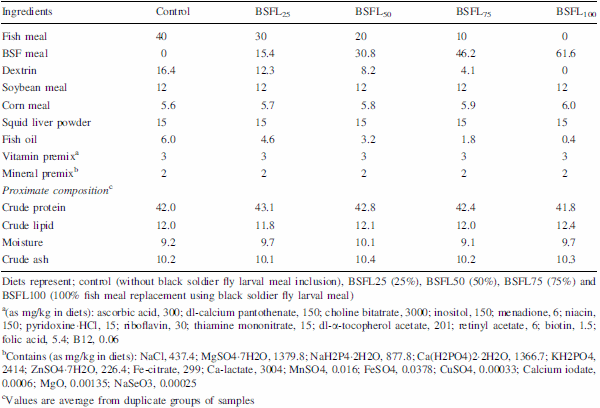
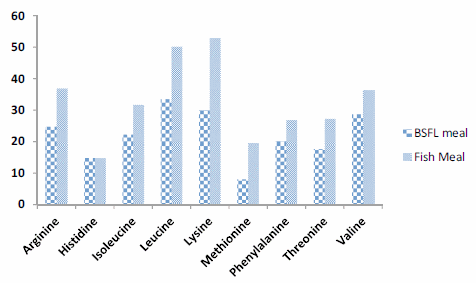
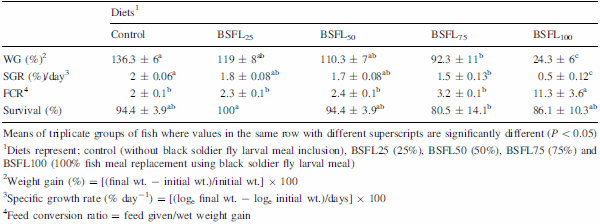
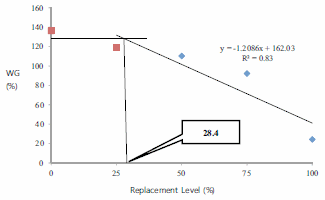
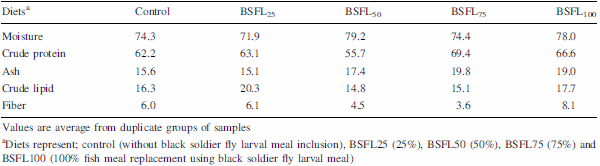
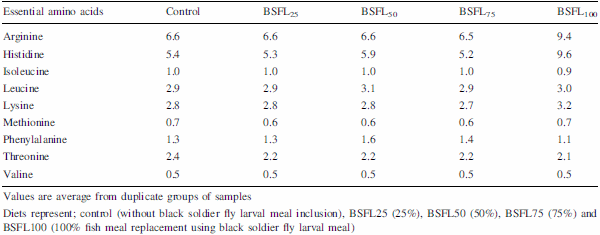
The present experiment was conducted to evaluate the efficacy of dietary protein from black soldier fly, Hermetia illucens, larval meal (BSFL) to replace fish meal (FM) protein in juvenile barramundi, Lates calcarifer. Larvae of black soldier fly were fed with the underutilised crop, sesbania, Sesbania grandiflora. Five isonitrogenous (44% crude protein) and isocaloric (16.0 kJ available energy/g) experimental diets were formulated to replace FM using processed BSFL meal at 0 (control), 25% (BSFL25), 50% (BSFL50), 75% (BSFL75) and 100% (BSFL100). Data for proximate and amino acid analysis suggested BSFL meal as an inferior protein ingredient than FM, but parallel to soybean meal. At the end of 8 weeks of fish feeding trial, there were no significant differences in the average weight gain (WG) and specific growth rate among the group of fish-fed control, BSFL25 and BSFL50 diets (P\0.05). Although numerical differences were recorded in the fish whole-body proximate composition, crude protein and moisture content were not much affected by the different dietary treatments. Essential amino acids including arginine, histidine, lysine and methionine were found to be higher in the whole body of fish-fed BSFL100 diet. Broken line regression analysis of average WG showed an optimum FM replacement level of 28.4% with BSFL meal. Therefore, the present experiment clearly demonstrates that the maximal dietary inclusion level of BSFL meal as FM protein replacer for the optimum growth of juvenile barramundi reared in freshwater could be greater than 28.4% but less than 50%, without any adverse effects on the fish whole-body proximate and amino acid composition.
Keywords Black soldier fly, Fish meal, Growth, Amino acids, Barramundi.

Adesulu EA, Mustapha AK (2000) Use of housefly maggots as a fishmeal replacer in tilapia culture: a recent vogue in Nigeria. In: Ftzimmons K, Filho JC (eds) Proceedings of the fifth international symposium on tilapia aquaculture, pp 138–143
Ajani EK, Nwanna LC, Musa BO (2004) Replacement of fishmeal with maggot meal in the diets of Nile tilapia, Oreochromis niloticus. J World Aquac Soc 35(52–277):54
Akiyama DM, Coelho SR, Lawrence AL, Robinson EH (1989) Apparent digestibility of feedstuffs by the marine shrimp Penaeus vannamei Boone. Nippon Suisan Gakkaishi 55:91–98
Alegbeleye WO, Obasa SO, Olude OO, Otubu K, Jimoh W (2002) Preliminary evaluation of the nutritive value of the variegated grasshopper (Zonocerus variegatus L.) for African catfish Clarias gariepinus (Burchell. 1822) fingerlings. Aquac Res 43:412–420
AOAC (1995) Official methods of analysis, 16th edn. Association of Official Analytical Chemists, Arlington
Bai SC, Kim KW (1997) Effects of dietary animal protein sources on growth and body composition in Korean rockfish, Sebastes schlegelii. J Aquac 10:77–85 (in Korean with English Abstract)
Bai SC, Jang HK, Kim KH (1997) Evaluation of feather meal and meat and bone meal as the fish meal replacer in Israeli carp diets. J Aquac 10:153–161
Bai SC, Jang HK, Cho ES (1998) Possible use of the animal by-product mixture as a dietary fish meal replacer in growing common carp (Cyprinus carpio). J Korean Fish Soc 31:380–385
Bondari K, Sheppard DC (1987) Soldier fly, Hermetia illucens L., larvae as feed for channel catfish, Ictalurus punctatus (Rafinesque), and blue tilapia, Oreochromis aureus (Steindachner). Aquac Res 18(3):209–220
Boonyaratpalin M, Suraneiranat P, Tunpibal T (1998) Replacement of fish meal with various types of soybean products in diets for the Asian sea bass, Lates calcarifer. Aquaculture 161:67–78
Carter CG, Hauler RC (2000) Fish meal replacement by plant meals in extruded feeds for Atlantic salmon, Salmo salar L. Aquaculture 185:299–311
Choi SM, Wang XJ, Lim SR, Kim KW, Bai SC, Shin IS (2004) Dietary dehulled soybean meal as a replacement for fish meal in fingerling and growing olive flounder, Paralichthys olivaceus (Temminck et Schlegel). Aquac Res 35:410–418
Cummins VC, Webster CD, Thompson KR, Velasquez A (2013) Replacement of fish meal with soybean meal, alone or in combination with distiller’s dried grains with solubles, in practical diets for Pacific white shrimp, Litopenaeus vannamei, grown in a clear-water culture system. J World Aquac Soc 44:775–785
Cummins VC Jr, Rawles SD, Thompson KR, Velasquez A, Kobayashi Y, Hager J, Webster CD (2017) Evaluation of black soldier fly (Hermetia illucens) larvae meal as partial or total replacement of marine fish meal in practical diets for Pacific white shrimp (Litopenaeus vannamei). Aquaculture 473:337–344
El-Sayed A-FM (1998) Total replacement of fishmeal with animal protein sources in Nile tilapia (Oreochromis niloticus) feeds. Aquac Res 29:275–280
FAO (Food and Agricultural Organization of the United Nations) (2009) State of world fisheries and aquaculture 2008 (SOFIA).
FAO Fisheries and Aquaculture Department, Rome
Fasakin EA, Balogun AM, Ajayi OO (2003) Evaluation of full-fat and defatted maggot meals in the feeding of clariid catfish Clarias gariepinus fingerlings. Aquac Res 34:733–738
Gatlin DM, Barrows FT, Brown P, Dabrowski K, Gaylord TG, Hardy RW, Herman E, Hu G, Krogdahl A, Nelson R, Overturf K, Rust M, Sealy W, Skonberg D, Souza EJ, Stone D, Wilson R, Wurtele E (2007) Expanding the utilization of sustainable plant products in aquafeeds: a review. Aquac Res 38:551–579
Glencross BD (2003) Can modelling assist with the determination of nutrient requirements in fish diet? In: van Barneveld RJ (ed) FRDC aquaculture nutrition subprogram annual workshop, Fremantle, WA, Australia. 29 May. pp 65–68
Katya K, Yun YH, Park G, Lee JY, Yoo G, Bai SC (2014) Evaluation of the efficacy of fermented by-product of mushroom, Pleurotus ostreatus, as a fish meal replacer in juvenile Amur catfish, Silurus asotus: effects on growth, serological characteristics and immune responses. Asian Australas J Anim Sci 27(10):1478–1486
Kim YC, Yoo GY, Wang X, Lee J, Shin IS, Bai SC (2008) Long term feeding effects of dietary dehulled soybean meal as a fish meal replacer in growing olive flounder, Paralichthys olivaceus. Asian Aust J Anim Sci 21:868–872
Kroeckel S, Harjes A-GE, Roth I, Katz H, Wuertz S, Susenbeth A, Schulz C (2012) When a turbot catches a fly: evaluation of a pre-pupae meal of the black soldier fly (Hermetia illucens) as fish meal substitute—growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 364:345–352
Lee KJ, Bai SC (1997a) Haemoglobin powder as a dietary fish meal replacer in juvenile Japanese eel, Anguilla japonica (Temminck et Schlegel). Aquac Res. 28:509–516
Lee KJ, Bai SC (1997b) Hemoglobin powder as a dietary animal protein source for juvenile Nile tilapia. Prog Fish-Cult. 59:266–271
Lim SR, Choi SM, Wang XJ, Kim KW, Shin IS, Min TS, Bai SC (2004) Effects of dehulled soybean meal as a fish meal replacer in diets for fingerling and growing Korean rockfish, Sebastes schlegelii. Aquaculture 231:457–468
Lock ER, ArisIwala T, Wagbo R (2015) Insect larval meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postmolt. Aquac Nutr 22(6):1–4
Mansour CR (1998) Nutrient requirements of red tilapia fingerlings. MSc. Thesis, Faculty of Science. University of Alexandria, Alexandria, Egypt, p 121
Muros MJS, De Haro C, Sanz A, Villareces S, Barroso FG (2015) Nutritional evaluation of Tenebrio molitor meal as fishmeal substitute for Tilapia (Oreochromis niloticus) diet. Aquac Nutr 22(5):943–955
Ogunji JO, Nimptsch J, Wiegand C, Schultz C (2007) Evaluation of the influence of housefly maggot meal (magmeal) diets on catalase, glutathione S-transferase and glycogen concentration in the liver of Oreochromis niloticus fingerling. Comp Biochem Physiol Part A: Mol Integr Physiol 147:942–947
Ogunji JO, Kloas W, Wirth M, Neumann N, Pietsch C (2008) Effect of housefly maggot meal (magmeal) diets on the performance, concentration of plasma glucose, cortisol and blood characteristics of Oreochromis niloticus fingerlings. J Anim Physiol Anim Nutr (Berlin) 92:511–518
Rana KS, Salam MA, Hashem S, Islam MA (2015) Development of black soldier fly larvae production technique as an alternate fish feed. Int J Res Fish Aquac 5(1):41–47
Refstie S, Storebakken T, Baeverfjord G, Roem A (2001) Long-term protein and lipid growth of Atlantic salmon (Salmo salar) fed diets with partial replacement of fish meal by soy protein products at medium or high lipid levels. Aquaculture 193:91–106
Robbins KR, Norton HW, Baker DH (1979) Estimation of nutrient requirements from growth data. J Nutr. 109(10):1710–1714
Rumpold BA, Schlu¨ter OK (2013) Potential and challenges of insects as an innovative source for food and feed production. Innov Food Sci Emerg Technol 17:1–11
Rust MB (2002) Nutritional physiology. In: Halver JE, Hardy RW (eds) Fish nutrition. Academic Press, New York, pp 368–446
Shiau SY, Yu YP (1999) Dietary supplementation of chitin and chitosan depresses growth in tilapia, Oreochromis niloticus9 O. aureus. Aquaculture 179:439–446
Smith JL, Opekun AR, Larkai E, Graham DY (1989) Sensitivity of the esophageal mucosa to pH in gastroesophageal reflux disease. Gastroenterology 96:683–689
St-Hilaire S, Cranfill K, McGuire MA, Mosley EE, Tomberlin JK, Newton L, Sealey W, Sheppard C, Irving S (2007a) Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J World Aquac Soc 38:309–313
St-Hilaire S, Sheppard C, Tomberlin JK, Irving S, Newton L, McGuire MA, Mosley E, Hardy RW, Sealey W (2007b) Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J World Aquac Soc 38:59–67
Suarez JA, Tudela C, Davis D, Daugherty Z, Taynor M, Glass L, Hoenig R, Buentello A, Beenetti DD (2013) Replacement of fish meal by a novel non-GM variety of soybean meal in cobia, Rachycentron canadum: ingredient nutrient digestibility and growth performance. Aquaculture 416–417:328–333
Trosvik KA, Rawles SD, Thompson KR, Metts LA, Gannam A, Twibell R, Webster CD (2012) Growth and body composition of Nile tilapia, Oreochromis niloticus, fry fed organic diets containing yeast extract and soybean meal as replacement for fish meal, with and without supplemental lysine and methionine. J World Aquac Soc 43:635–647
Van der Spiegel M, Noordam MY Van, der Fels-Klerx HJ (2013) Safety of novel protein sources (insects, microalgae, seaweed, duckweed, and rapeseed) and legislative aspects for their application in food and feed production. Comp Revis Food Sci Food Saf 12:662–678
Van Huis A (2013) Potential of insects as food and feed in assuring food security. Annu Revis Entomol 58:563–583 Wing-Keong N, Liew F-L, Ang LP, Wong KW (2001) Potential of mealworm (Tenebrio molitor) as an alternative protein source in practical diets for African catfish, Clarias gariepinus. Aquac Res 32:273–280
Yu GH, Chen YH (2009) Research progression on the larvae and prepupae of black soldier fly Hermetia illucens used as animal feedstuff. Chin Bull Entomol 46(1):41–45





Has anyone tried improving the results of BSF protein as an FM replacement by adding selected amounts of microalgae which can have a superior amino acid profile and lipid profile? That might answer part of the question whether it's the chitin content of BSF or the lower amino acid content that leads to the observed decrease in WG at a higher dosage?





")
