Organic trace mineral supplementation
Organic trace mineral supplementation: can success in animal nutrition be extrapolated to humans?
Published: October 25, 2011
By: RONAN POWER - Alltech Inc.
There can be little doubt that dietary habits in western populations have changed dramatically since the middle of the last century. Current trends are driven by convenience and changing domestic practices. Vegetarianism is on the increase, especially in younger females, and red meat consumption has fallen in response to BSE awareness and its possible link with vCJD. For example, vegetarianism in the UK currently stands at 5% of the population, with the figures doubled over a 15 year period (RealEat Gallup surveys, 1984-1999).
Likewise, over the last 60 years, an overall reduction in the daily energy intake of the UK population has occurred; decreasing from approximately 12.6 MJ per day in 1940 to 7.6 MJ in 1999 (MAFF, 1999; Wardle, 1977). These changes are further evidence of changing eating patterns and habits in many western countries and are a cause for some concern (Harvey, 2001). Decreases in energy intakes, coupled with the changing pattern of meat consumption, mean that mineral intakes are falling. Consequently, certain population groups are at an increased risk of suffering from mineral deficiencies. Of course, in developing countries, the situation is much worse. For example, the prevalence of iron deficiency in industrialized countries is less than 10%. In contrast, it is a very serious problem in the developing world, principally affecting infants, children and women (Pizarro et al., 2002). One of the last estimates released by the World Health Organization reported that at least 1.32 billion people, living mainly in less industrialized nations, suffered from iron deficiency anaemia (DeMayer, 1989).
Education programmes, and the wealth of literature pertaining to diet and health in developed countries, have increased awareness of the essential role of minerals in maintaining health and preventing serious disease. In more affluent societies, sales of human mineral supplements continue to rise. In the UK alone, sales of such products reach £400 million annually (Euromonitor, 2000). In underdeveloped countries, however, such interventions remain unaffordable and the best chance for improving the mineral status of at-risk populations is restricted to sporadic programmes of diet fortification with single essential trace elements, mainly iron. Allied with these shortcomings is the fact that concrete recommendations for mineral intake in humans are very limited (Harvey, 2001). For example, in the case of copper, several international expert committees are unable to define the levels of intake at which health is impaired. These difficulties are compounded by the fact that, with the possible exception of iron, little research has been conducted to determine the most biologically available chemical forms of trace elements for human use. The majority of over-the-counter mineral supplements contain (where present) inorganic forms of trace elements instead of complexes or chelates with organic ligands such as amino acids or oligopeptides.
Evidence from the field of animal nutrition suggests that trace elements, when administered in the form of ‘organic complexes’ are more biologically available than their inorganic counterparts.
From the foregoing, therefore, it makes sense to review the knowledge base that has been developed over the last several decades in the area of organic mineral supplementation in animals to see if meaningful benefits, relative to traditional inorganic mineral supplementation, have been attained. Given the near identical nature of physiological processes across the animal kingdom, benefits observed in animal nutrition may have a bearing on the choice of trace mineral supplementation for human use. The role of trace minerals in biological systems Although the essentiality of some trace elements is still a matter of contention, it is generally accepted that the trace elements now considered to be essential or beneficial to mammalian and avian species are: arsenic (As), boron (B), chromium (Cr), copper (Cu), fluorine (F), iodine (I), iron (Fe), manganese (Mn), molybdenum (Mo), nickel (Ni), selenium (Se), silicon (Si), vanadium (V) and zinc (Zn). Trace element supplementation of animal diets has long been practiced in order to ensure rapid growth rates, boost reproductive performance, avoid rious mineral deficiency syndromes and generally prevent infection and disease. In general, inorganic salts of trace elements, such as sulphates and chlorides, have been used to fortify animal diets.
However, because a large variety of physicochemical and biological factors are known to affect the biological availability of inorganic trace elements in animal nutrition, efforts to improve their uptake, utilization and retention have been ongoing for decades (Tables 1 and 2).
For most of the essential trace elements mentioned above, it is now well established that their provision in ‘organic’ form, e.g. transition metals such as Cu2+, Zn2+ and Mn2+ chelated to amino acids or peptides, can enhance the bioavailability of these trace elements, relative to inorganic forms. This has led to further benefits being noted in general health status, improved growth rates and reproductive performance. Mineral supplements for human use are, of course, not a recent innovation, but it is fair to say that many of the products available in the human health food market were conceived on the basis of beneficial effects being noted in animals.
Table 1. Common interactions and antagonisms affecting mineral utilization1.

1from Rucker et al., 1994
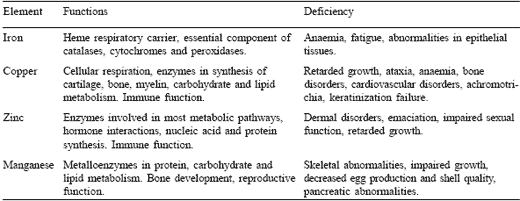
Table 2. Nutritional aspects of iron, copper, zinc and manganese.
IRON
Iron is involved in a multitude of physiological functions, ranging from its role as a redox cofactor to serving as a major constituent of heme, which contributes to the oxygen-carrying capacity of haemoglobin and muscle myoglobin. Consequences of iron deficiency can range from impaired behavioural and cognitive functions, and central nervous system disorders, to lethargy, anaemia, and reduced work capacity in humans (Cook, 1990; Finch and Cook, 1984). It should not be forgotten that while iron is an essential nutrient, an overload can cause cell and tissue damage, brought on by increased production of free oxygen radicals.
As with other essential trace elements, mechanisms exist in the gastrointestinal tract that can modulate the influx of iron across the mucosal barrier, thereby compensating to some degree during low and high intakes of iron. Endogenous losses of iron in the gut are very small, which implies that iron homeostasis is mediated primarily through absorption. A thorough understanding of the dietary factors affecting availability of iron for absorption and the mechanisms which regulate iron absorption are key requirements if the iron nutrition of animals (and humans) is to be optimized.
In the case of iron supplementation in animals, a major goal has been to improve the iron status of newborn animals, such as piglets, through the use of more bioavailable iron sources. For example, iron chelated to amino acids has been reported to lead to increased transfer of the metal across the placenta and into the embryo (Ashmead and Graff, 1982). When provided at 200 ppm in the gestation diet, greater quantities of iron were incorporated into the fetuses resulting in significantly reduced mortality and heavier piglets at birth and weaning (Ashmead, 1996). When administered over eight parities, there were fewer stillborn piglets and more piglets weaned in each parity, as well as a shorter interval between weaning and oestrus. Similar effects have been reported for iron proteinate incorporated into a normal lactation diet fed from seven days pre-farrowing and throughout a twenty six day lactation. This led to improved feed intake of the sows and increased weaning weights of the piglets (Close, 1999). In other studies, piglets from sows whose diets were supplemented with iron proteinate 21 days pre-farrowing, had a significantly higher erythrocyte count (P<0.05) and haemoglobin level (P<0.01) than piglets from sows receiving inorganic iron sources. In addition, liver iron levels were notably higher in piglets from the proteinatesupplemented group.
COPPER
Copper is a critical element for animals as it is an essential component of physiologically important metalloenzymes such as cytochrome oxidase, superoxide dismutase, lysyl oxidase, dopamine hydroxylase and tyrosinase. Overall, this metal is involved in cellular respiration, cardiac function, bone formation, connective tissue development, keratinization and pigmentation of tissue, as well as myelination of the spinal cord (McDowell, 1992).
Copper has a direct effect on iron metabolism and thus, indirectly, affects haemoglobin biosynthesis.
The first sign of copper deficiency experimentally observed was anaemia in rats. In those early studies, haemoglobin regeneration was used to estimate the bioavailability of various copper compounds (Schultze et al., 1934; 1936) and in later years to assess copper-amino acid and copper-peptide complexes (Kirchgessner and Grassman, 1970). The overall growth-promotional effects of copper are well established and are comprehensively reviewed elsewhere (Underwood, 1997; McDowell, 1992).
Several parallels can be drawn between essential trace elements such as iron, copper, zinc and manganese. This is particularly true for iron and copper. Much of their utility is based on their ability to assume at least two oxidation states so that they can participate in redox reactions. Unfortunately, this same property can lead, via the Fenton reaction (and Haber-Weiss reaction), to the formation of toxic reactive oxygen species. Another consequence is that both ferric iron and cuprous copper are essentially insoluble in aqueous solution at neutral pH and are subject to hydroxy-polymerization under the prevailing conditions in the small intestine. The potential toxicity of metals such as iron and copper and the challenge of maintaining their solubility in vivo are the reasons why these metals are associated with carriers or chaperones at nearly all stages of their assimilation and metabolism. When the capacity of the carrier/chaperone/storage systems is overloaded, the metal becomes toxic (Garrick et al., 2003).
Improving the bioavailability of copper by providing it to animals in such a carrier-bound form has met with a considerable measure of success.
For example, in ruminants bioplexed copper (copper proteinate) was reported to be more bioavailable than copper sulphate in studies involving beef cattle (Hemken et al., 1993). Further studies by the same group in dairy cows demonstrated increased hepatic iron content in copper proteinate- vs copper sulphate- supplemented animals. This suggested that the copper proteinate did not antagonize iron uptake and storage as is commonly observed with inorganic sources of these elements. In addition, it was found that copper proteinate-supplemented cows had lower plasma ceruloplasmin activity than cows fed copper sulphate, even though plasma copper levels were the same for both groups (Du et al., 1995).
These data suggested that copper proteinate may be absorbed via a different mechanism to copper sulphate and may have been transported in blood without binding to ceruloplasmin. However, very recent work has cast some doubt on the suitability of ceruloplasmin as a biomarker for copper status (Araya et al., 2003) meaning that any such assumptions about different transport mechanisms for different forms of copper will have to be verified by other means. A further point to make on copper bioavailability relates to plasma copper levels arising from the use of different copper sources. For example, it is interesting to note that the bioavailability of copper proteinate relative to copper sulphate has been calculated as either 147% or 112%, depending on whether liver copper or plasma copper is used as the response criterion ( Baker and Ammerman, 1995; Kincaid et al., 1986). Such findings are in agreement with recommendations that liver copper levels, as opposed to plasma levels, are a better indicator of copper status and relative bioavailability between sources (Lee et al., 1988; Xin et al., 1991).
ZINC
Zinc is the second most abundant trace element in the mammalian body and is required as a component of over 300 enzymes in different species (Vallee and Falchuk, 1993). These include carbonic anhydrase, alcohol dehydrogenase and alkaline phosphatase. In association with enzymes, zinc plays an active catalytic role, generally as a strong Lewis acid or acts in a regulatory or structural role.
Biologically, it is a trace element of immense importance, significant effects on the production of steroid and peptide hormones, which may offer an explanation for some of the well documented effects of zinc deficiency. These include growth retardation, impaired reproductive development/function, aberrant water and cation balance and parakeratosis. Apart from its wellestablished structural role in key metalloenzymes, zinc performs unique structural functions at a nucleic acid level (Evans and Hollenberg, 1988). Such functions underscore the essentiality of zinc in many crucial biological processes.
MANGANESE
Manganese, like the other essential trace elements, can function both as an enzyme activator and as a constituent of metalloenzymes. Manganesecontaining enzymes include arginase, pyruvate carboxylase and Mn-superoxide dismutase.
Although the number of Mn-metalloenzymes is limited, a large number of enzymes can be activated by manganese. These include hydrolases, kinases, decarboxylases and transferases. Manganese is a vital element for bone growth, carbohydrate and lipid metabolism, immune and nervous system function and reproduction. Indeed, effects on reproduction were among the first signs of Mn deficiency to be noted.
Studies in poultry have revealed notable differences in the bioavailability of zinc from different sources. For example, it has been reported that the bioavailability of zinc from zinc methionine is 206% relative to that of zinc sulphate (Wedekind et al., 1992). Other indicators of the improved bioavailability of organic versus inorganic zinc sources in poultry include studies documenting significantly positive effects on gonad development in roosters receiving zinc proteinate versus zinc sulphate (Suchy et al., 1998). In addition, studies have demonstrated that replacement of inorganic zinc and manganese sources with zinc and manganese proteinate improved egg shell quality (P<0.05; Miles, 1998).
Indeed, many studies on the relative bioavailability of manganese have been conducted in poultry where there is a critical supplemental need for the element. Early studies with poultry, in which growth or leg deformities were measured, were not sufficiently sensitive to detect differences in bioavailability between supplemental sources. However, over the last few decades, tissue deposition of the element has been used to estimate manganese bioavailability. Such studies have revealed that the most available sources of manganese are manganese methionine and manganese proteinate (Henry, 1995).
Organic trace minerals for human use
Not surprisingly, in the field of human nutrition, much attention has also been paid to enhancing the bioavailability of essential trace elements. In many cases, strategies for human mineral supplementation have been developed from observations in animal models. Organic trace mineral complexes, mainly derived from chelation reactions with amino acids or short peptides have, as in animal nutrition, proven to be among the most successful interventions available. In the particular case of iron deficiency anaemia, the concept of using protein-derived material to boost iron absorption is quite an old one (Klavins et al., 1959; Klavins et al., 1962). These workers postulated that animal protein exerts an enhancing effect on the absorption of dietary iron.
Animal and vegetable foods were biosynthetically labeled with different radioisotopes of iron, and the incorporation of these radioiron forms into the circulating haemoglobin were taken as an index of absorption. It was found that veal increased, about 2-fold, the absorption of both maize and black bean iron. Subsequent studies revealed that certain amino acids, particularly cysteine, were responsible for the observed enhancement in iron absorption (Martinez- Torres et al., 1971).
This concept was further developed by further experiments which demonstrated that cysteine-containing peptides, rather than the free amino acid, were the causative agents for enhanced iron absorption. Two types of peptic digestion extracts were prepared from beef, in which the thiol groups were either oxidized (CYS -) or left untreated (CYS +). The absorption of radioiron mixed with maize was more than 2- fold greater when consumed along with the CYS+ extract than with the CYS- extract (P<0.005; Taylor et al., 1986).
In many respects, this seminal work led the way in the development of chelated organic trace minerals for human use, with the main emphasis being on facilitators of iron absorption. Today, several classes of compounds have been identified which assist iron absorption in humans (Table 3). Many studies have been published in recent years which show clear benefits in mineral status from dietary supplementation with chelated, or otherwise complexed trace elements.
In an effort to boost the mineral status of infants, many preparations are designed for inclusion in powdered or liquid milk formulae. In liquid formulae where the relative solubilities of copper, iron and zinc were examined, it was found that substituting organic forms of copper and iron in the mix resulted in an almost 3-fold increase in solubility while zinc solubilities were not improved. These results were reflected in enhanced absorption of copper and iron from the organic formula vs the inorganic formula, with the result for iron being statistically significant (P<0.05; Hendricks et al., 2001). Ferrous amino acid chelates are increasingly being used to combat Fe-deficiency anaemia through liquid milk fortification. Recent studies have shown that an iron bis-glycinate chelate, administered at a level of 3 mg/liter/day, was successful in restoring normal haemoglobin concentrations in a population of Brazilian children (Iost et al., 1998).
Table 3. Compounds indicated as facilitators of iron absorption1.
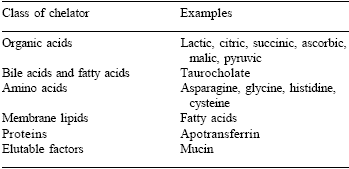
1Cremonesi et al., 2002.
Additional studies with this chelate involved 40 infants with Fe-deficiency anaemia being assigned to two groups, receiving either ferrous sulphate or the ferrous bis-glycinate chelate at a level of 5 mg Fe/kg bodyweight, administered via a syrup.
Significant increases in haemoglobin levels were observed in both groups but only the ferrous bisglycinate group displayed significant increases in plasma ferritin (Pineda and Ashmead, 2001). This material has also been successfully used to enhance iron absorption from foods enriched with phytate and polyphenols. In this case, absorption of iron from the iron glycinate was approximately twice that from ferrous sulphate (P<0.05; Layrisse et al., 2000).
Comparisons in the literature between the ferrous bis-glycinate chelate and other sources of organic iron are rather limited. One study examining the relative bioavailabilities of the iron bis-glycinate and ferrous ascorbate, when administered in drinking water, failed to demonstrate any significant difference between them (Olivares and Pizarro, 2001).
Other organic iron preparations used in the treatment of Fe-deficiency anaemia are designed with both enhanced bioavailability and better tolerability in mind. Unfortunately, orally administered iron compounds, particularly iron sulphate, may cause irritation and damage to the intestinal mucosa. Two types of polymeric ferric compounds have received particular attention in this regard, namely ferric polysaccharide complexes and iron protein succinylate (Raja et al., 2000). Positive therapeutic effects against Fe-deficiency anaemia without gastrointestinal side effects have been achieved with the iron protein succinylate (Cremonesi et al., 1984). A comparative study involving both organic iron forms indicated that both were well tolerated in children, but that the protein succinylate form was more effective in increasing iron stores (Haliotis and Papanastasiou, 1998).
Other organic iron complexes which have been proven to be effective in the treatment of anaemia, without adverse side effects, include ferric trimaltol (Reffitt et al., 2000).
From the available literature, one might be forgiven in thinking that the use of organic trace element supplementation in humans is restricted to the treatment of Fe-deficiency anaemia. Certainly, the majority of published work to date has focused on the treatment of Fe-deficiency anaemia but other interesting studies are beginning to emerge. For example, recent trials examined the therapeutic value of a broad-based nutritional supplement, predominantly composed of chelated trace minerals, on the treatment of bipolar disorder. The overall findings of these studies were that symptom reduction over the six-month duration of the trials ranged from 55% to 66% and the need for psychotropic medicines decreased by more than 50% (Kaplan et al., 2001). Other studies have shown low plasma levels of zinc and other minerals in people with mood and behavior disorders. For example, lower serum zinc levels were recorded in 48 unipolar depressed patients in comparison to 32 volunteers with normal mental health (Maes et al., 1994; Maes et al., 1997). In addition, a study which evaluated the copper and zinc levels in 135 assaultive, incarcerated males in comparison to controls found an inverse relationship between zinc and the severity of the behaviour, ranging from verbal assault to aggravated and violent assault; the lower the zinc, the greater the rate of violent behaviour (Walsh et al., 1997).
Such findings further assert the importance of certain trace minerals in diverse physiological processes and raise interesting possibilities for the amelioration of different ailments, both physical and psychological, through the enhancement of trace mineral status.
Uptake mechanisms for organic minerals in the gastrointestinal tract
There is clear evidence from the literature that differences in bioavailability exist between sources of the same trace metal, with metal proteinates and chelates proving superior to inorganic sources in many cases. If it can be accepted that uptake of metals from the intestine is the predominant factor influencing their bioavailability (Dreosti, 1993), then differences in uptake mechanisms or in the general presentation of the metal in organic versus inorganic form in the intestinal lumen are likely explanations for the differences noted.
The theory that metal amino acid chelates and proteinates utilize amino acid uptake and peptide uptake mechanisms rather than regular inorganic metal ion uptake pathways has become widely accepted (Ashmead et al., 1985; Ashmead, 1993).
The basic theory holds that the metal in question is protected within the complex in a chemically inert form. Consequently, the metal is not susceptible to the range of physicochemical factors which can adversely affect the efficient uptake of ‘unprotected’ mineral ions. Furthermore, it is hypothesized that the organic complex is absorbed intact through the intestinal mucosa, effectively carrying the metal along with it. The complex traverses the mucosal cell membrane, the mucosal cell and basement membrane, surviving, intact into the plasma.
If such an uptake mechanism did exist, it would help to explain numerous practical observations from studies on the relative bioavailability of organic versus inorganic sources in both animals and humans. The reductions noted in negative interactions between elements such as copper, iron and zinc when organic supplements are used agree well with this theory (Lowe et al., 1994; Hemken et al., 1996). For example, if copper in proteinated form utilizes a peptide uptake mechanism, it does not have to compete with zinc for regular metal ion uptake pathways.
Conclusive evidence for the existence of this kind of uptake mechanism has not been published, however. Although there is strong evidence that selenium in the form of selenomethionine enters the enterocyte via an electrogenic Na+- dependent neutral amino acid transport system in a manner which is indistinguishable from that of methionine (Vendeland et al., 1994), selenomethionine cannot be directly compared to structures such as ferrous bis-glycinate or zinc methionine. In selenomethionine, selenium replaces sulphur to become integrated into the molecular structure of methionine. It is inaccurate, therefore to draw an analogy between selenomethionine uptake and coordination complexes such as zinc methionine or iron glycine.
Some of the more recent studies in humans indicate that the processing of inorganic and organic minerals at the point of absorption may share more common elements than previously thought. For example, absorption studies on ferrous bis-glycinate reveal strong evidence that this substance uses the classic non-heme iron uptake pathway rather than the pathway for heme absorption (Pizarro et al., 2002). In this study, it was shown that iron from the bis-glycinate competes directly with ferrous sulphate for the non-heme uptake route in the intestinal lumen. Likewise, the finding that the intestinal absorption of iron (ferric) protein succinylate is inhibited by a ferrous chelator, ferrozine, indicates that reduction of ferric iron in this complex to ferrous iron must occur before absorption can take place.
This indicates a disruption of the complex and argues against uptake as an intact entity. The authors conclude that iron protein succinylate is absorbed by a mechanism involving digestion to release soluble, available ferric species which are reduced at the mucosal surface to provide ferrous iron for membrane transport into the enterocytes (Raja et al., 2000). Another organic iron complex, ferric trimaltol, has been shown to dissociate upon reaching the intestinal mucosa (Reffitt, 2000).
Today, several models exist for the uptake of trace elements from the gastrointestinal tract. The most popular of these is based upon concepts put forward by Conrad and Umbreit in 1993. As trace elements leave the acidic environment of the stomach and enter the alkaline environment of the small intestine, there is a tendency for certain elements, such as iron, copper, manganese and zinc, to undergo hydroxypolymerization (Powell et al., 1999). This results in the formation of large, insoluble metal hydroxide or metal oxide polymers that remain unavailable for uptake and utilization. For example, it has been estimated that in the pH range from 5.5 to 7.0, an increase of one pH unit causes a 30 to 45-fold decrease in aqueous Zn2+ ion concentration.
Similarly, an increase of one pH unit causes a 100- and 1,000-fold decrease in aqueous Fe2+ and Fe3+ ion concentration, respectively (Frossard et al., 2000). In the model of Conrad and Umbreit, ligands present in the structure of organic mineral products keep the metal soluble at alkaline pH until it can be donated to mucins. Mucins are large metal-binding glycoproteins that predominantly reside in the mucosally-adherent mucus layer. In the case of iron, mucin-bound iron subsequently crosses the mucosal cell membrane in association with integrins. Once inside the cell, a cytoplasmic iron-binding protein, dubbed ‘mobilferrin’, accepts the element and shuttles it to the basolateral surface of the cell where it is delivered to the plasma. The entire absorptive process is hypothesized to be driven by a cascade of differences in the binding constants of proteins so that iron moves from luminal mucin to cytoplasmic mobilferrin to plasma transferrin.
Whatever the mechanism, however, there is no doubt that the provision of minerals in organic form for human and animal use has been beneficial in terms of enhancing the bioavailability of essential trace elements, thereby contributing to improved health and performance. One can only look forward to further developments in this field, in areas such as the development of more effective ligands together with a more complete understanding of the relevant uptake mechanisms involved so that new approaches can be taken to influence them.
References
Likewise, over the last 60 years, an overall reduction in the daily energy intake of the UK population has occurred; decreasing from approximately 12.6 MJ per day in 1940 to 7.6 MJ in 1999 (MAFF, 1999; Wardle, 1977). These changes are further evidence of changing eating patterns and habits in many western countries and are a cause for some concern (Harvey, 2001). Decreases in energy intakes, coupled with the changing pattern of meat consumption, mean that mineral intakes are falling. Consequently, certain population groups are at an increased risk of suffering from mineral deficiencies. Of course, in developing countries, the situation is much worse. For example, the prevalence of iron deficiency in industrialized countries is less than 10%. In contrast, it is a very serious problem in the developing world, principally affecting infants, children and women (Pizarro et al., 2002). One of the last estimates released by the World Health Organization reported that at least 1.32 billion people, living mainly in less industrialized nations, suffered from iron deficiency anaemia (DeMayer, 1989).
Education programmes, and the wealth of literature pertaining to diet and health in developed countries, have increased awareness of the essential role of minerals in maintaining health and preventing serious disease. In more affluent societies, sales of human mineral supplements continue to rise. In the UK alone, sales of such products reach £400 million annually (Euromonitor, 2000). In underdeveloped countries, however, such interventions remain unaffordable and the best chance for improving the mineral status of at-risk populations is restricted to sporadic programmes of diet fortification with single essential trace elements, mainly iron. Allied with these shortcomings is the fact that concrete recommendations for mineral intake in humans are very limited (Harvey, 2001). For example, in the case of copper, several international expert committees are unable to define the levels of intake at which health is impaired. These difficulties are compounded by the fact that, with the possible exception of iron, little research has been conducted to determine the most biologically available chemical forms of trace elements for human use. The majority of over-the-counter mineral supplements contain (where present) inorganic forms of trace elements instead of complexes or chelates with organic ligands such as amino acids or oligopeptides.
Evidence from the field of animal nutrition suggests that trace elements, when administered in the form of ‘organic complexes’ are more biologically available than their inorganic counterparts.
From the foregoing, therefore, it makes sense to review the knowledge base that has been developed over the last several decades in the area of organic mineral supplementation in animals to see if meaningful benefits, relative to traditional inorganic mineral supplementation, have been attained. Given the near identical nature of physiological processes across the animal kingdom, benefits observed in animal nutrition may have a bearing on the choice of trace mineral supplementation for human use. The role of trace minerals in biological systems Although the essentiality of some trace elements is still a matter of contention, it is generally accepted that the trace elements now considered to be essential or beneficial to mammalian and avian species are: arsenic (As), boron (B), chromium (Cr), copper (Cu), fluorine (F), iodine (I), iron (Fe), manganese (Mn), molybdenum (Mo), nickel (Ni), selenium (Se), silicon (Si), vanadium (V) and zinc (Zn). Trace element supplementation of animal diets has long been practiced in order to ensure rapid growth rates, boost reproductive performance, avoid rious mineral deficiency syndromes and generally prevent infection and disease. In general, inorganic salts of trace elements, such as sulphates and chlorides, have been used to fortify animal diets.
However, because a large variety of physicochemical and biological factors are known to affect the biological availability of inorganic trace elements in animal nutrition, efforts to improve their uptake, utilization and retention have been ongoing for decades (Tables 1 and 2).
For most of the essential trace elements mentioned above, it is now well established that their provision in ‘organic’ form, e.g. transition metals such as Cu2+, Zn2+ and Mn2+ chelated to amino acids or peptides, can enhance the bioavailability of these trace elements, relative to inorganic forms. This has led to further benefits being noted in general health status, improved growth rates and reproductive performance. Mineral supplements for human use are, of course, not a recent innovation, but it is fair to say that many of the products available in the human health food market were conceived on the basis of beneficial effects being noted in animals.
Table 1. Common interactions and antagonisms affecting mineral utilization1.
1from Rucker et al., 1994
Table 2. Nutritional aspects of iron, copper, zinc and manganese.
IRON
Iron is involved in a multitude of physiological functions, ranging from its role as a redox cofactor to serving as a major constituent of heme, which contributes to the oxygen-carrying capacity of haemoglobin and muscle myoglobin. Consequences of iron deficiency can range from impaired behavioural and cognitive functions, and central nervous system disorders, to lethargy, anaemia, and reduced work capacity in humans (Cook, 1990; Finch and Cook, 1984). It should not be forgotten that while iron is an essential nutrient, an overload can cause cell and tissue damage, brought on by increased production of free oxygen radicals.
As with other essential trace elements, mechanisms exist in the gastrointestinal tract that can modulate the influx of iron across the mucosal barrier, thereby compensating to some degree during low and high intakes of iron. Endogenous losses of iron in the gut are very small, which implies that iron homeostasis is mediated primarily through absorption. A thorough understanding of the dietary factors affecting availability of iron for absorption and the mechanisms which regulate iron absorption are key requirements if the iron nutrition of animals (and humans) is to be optimized.
In the case of iron supplementation in animals, a major goal has been to improve the iron status of newborn animals, such as piglets, through the use of more bioavailable iron sources. For example, iron chelated to amino acids has been reported to lead to increased transfer of the metal across the placenta and into the embryo (Ashmead and Graff, 1982). When provided at 200 ppm in the gestation diet, greater quantities of iron were incorporated into the fetuses resulting in significantly reduced mortality and heavier piglets at birth and weaning (Ashmead, 1996). When administered over eight parities, there were fewer stillborn piglets and more piglets weaned in each parity, as well as a shorter interval between weaning and oestrus. Similar effects have been reported for iron proteinate incorporated into a normal lactation diet fed from seven days pre-farrowing and throughout a twenty six day lactation. This led to improved feed intake of the sows and increased weaning weights of the piglets (Close, 1999). In other studies, piglets from sows whose diets were supplemented with iron proteinate 21 days pre-farrowing, had a significantly higher erythrocyte count (P<0.05) and haemoglobin level (P<0.01) than piglets from sows receiving inorganic iron sources. In addition, liver iron levels were notably higher in piglets from the proteinatesupplemented group.
COPPER
Copper is a critical element for animals as it is an essential component of physiologically important metalloenzymes such as cytochrome oxidase, superoxide dismutase, lysyl oxidase, dopamine hydroxylase and tyrosinase. Overall, this metal is involved in cellular respiration, cardiac function, bone formation, connective tissue development, keratinization and pigmentation of tissue, as well as myelination of the spinal cord (McDowell, 1992).
Copper has a direct effect on iron metabolism and thus, indirectly, affects haemoglobin biosynthesis.
The first sign of copper deficiency experimentally observed was anaemia in rats. In those early studies, haemoglobin regeneration was used to estimate the bioavailability of various copper compounds (Schultze et al., 1934; 1936) and in later years to assess copper-amino acid and copper-peptide complexes (Kirchgessner and Grassman, 1970). The overall growth-promotional effects of copper are well established and are comprehensively reviewed elsewhere (Underwood, 1997; McDowell, 1992).
Several parallels can be drawn between essential trace elements such as iron, copper, zinc and manganese. This is particularly true for iron and copper. Much of their utility is based on their ability to assume at least two oxidation states so that they can participate in redox reactions. Unfortunately, this same property can lead, via the Fenton reaction (and Haber-Weiss reaction), to the formation of toxic reactive oxygen species. Another consequence is that both ferric iron and cuprous copper are essentially insoluble in aqueous solution at neutral pH and are subject to hydroxy-polymerization under the prevailing conditions in the small intestine. The potential toxicity of metals such as iron and copper and the challenge of maintaining their solubility in vivo are the reasons why these metals are associated with carriers or chaperones at nearly all stages of their assimilation and metabolism. When the capacity of the carrier/chaperone/storage systems is overloaded, the metal becomes toxic (Garrick et al., 2003).
Improving the bioavailability of copper by providing it to animals in such a carrier-bound form has met with a considerable measure of success.
For example, in ruminants bioplexed copper (copper proteinate) was reported to be more bioavailable than copper sulphate in studies involving beef cattle (Hemken et al., 1993). Further studies by the same group in dairy cows demonstrated increased hepatic iron content in copper proteinate- vs copper sulphate- supplemented animals. This suggested that the copper proteinate did not antagonize iron uptake and storage as is commonly observed with inorganic sources of these elements. In addition, it was found that copper proteinate-supplemented cows had lower plasma ceruloplasmin activity than cows fed copper sulphate, even though plasma copper levels were the same for both groups (Du et al., 1995).
These data suggested that copper proteinate may be absorbed via a different mechanism to copper sulphate and may have been transported in blood without binding to ceruloplasmin. However, very recent work has cast some doubt on the suitability of ceruloplasmin as a biomarker for copper status (Araya et al., 2003) meaning that any such assumptions about different transport mechanisms for different forms of copper will have to be verified by other means. A further point to make on copper bioavailability relates to plasma copper levels arising from the use of different copper sources. For example, it is interesting to note that the bioavailability of copper proteinate relative to copper sulphate has been calculated as either 147% or 112%, depending on whether liver copper or plasma copper is used as the response criterion ( Baker and Ammerman, 1995; Kincaid et al., 1986). Such findings are in agreement with recommendations that liver copper levels, as opposed to plasma levels, are a better indicator of copper status and relative bioavailability between sources (Lee et al., 1988; Xin et al., 1991).
ZINC
Zinc is the second most abundant trace element in the mammalian body and is required as a component of over 300 enzymes in different species (Vallee and Falchuk, 1993). These include carbonic anhydrase, alcohol dehydrogenase and alkaline phosphatase. In association with enzymes, zinc plays an active catalytic role, generally as a strong Lewis acid or acts in a regulatory or structural role.
Biologically, it is a trace element of immense importance, significant effects on the production of steroid and peptide hormones, which may offer an explanation for some of the well documented effects of zinc deficiency. These include growth retardation, impaired reproductive development/function, aberrant water and cation balance and parakeratosis. Apart from its wellestablished structural role in key metalloenzymes, zinc performs unique structural functions at a nucleic acid level (Evans and Hollenberg, 1988). Such functions underscore the essentiality of zinc in many crucial biological processes.
MANGANESE
Manganese, like the other essential trace elements, can function both as an enzyme activator and as a constituent of metalloenzymes. Manganesecontaining enzymes include arginase, pyruvate carboxylase and Mn-superoxide dismutase.
Although the number of Mn-metalloenzymes is limited, a large number of enzymes can be activated by manganese. These include hydrolases, kinases, decarboxylases and transferases. Manganese is a vital element for bone growth, carbohydrate and lipid metabolism, immune and nervous system function and reproduction. Indeed, effects on reproduction were among the first signs of Mn deficiency to be noted.
Studies in poultry have revealed notable differences in the bioavailability of zinc from different sources. For example, it has been reported that the bioavailability of zinc from zinc methionine is 206% relative to that of zinc sulphate (Wedekind et al., 1992). Other indicators of the improved bioavailability of organic versus inorganic zinc sources in poultry include studies documenting significantly positive effects on gonad development in roosters receiving zinc proteinate versus zinc sulphate (Suchy et al., 1998). In addition, studies have demonstrated that replacement of inorganic zinc and manganese sources with zinc and manganese proteinate improved egg shell quality (P<0.05; Miles, 1998).
Indeed, many studies on the relative bioavailability of manganese have been conducted in poultry where there is a critical supplemental need for the element. Early studies with poultry, in which growth or leg deformities were measured, were not sufficiently sensitive to detect differences in bioavailability between supplemental sources. However, over the last few decades, tissue deposition of the element has been used to estimate manganese bioavailability. Such studies have revealed that the most available sources of manganese are manganese methionine and manganese proteinate (Henry, 1995).
Organic trace minerals for human use
Not surprisingly, in the field of human nutrition, much attention has also been paid to enhancing the bioavailability of essential trace elements. In many cases, strategies for human mineral supplementation have been developed from observations in animal models. Organic trace mineral complexes, mainly derived from chelation reactions with amino acids or short peptides have, as in animal nutrition, proven to be among the most successful interventions available. In the particular case of iron deficiency anaemia, the concept of using protein-derived material to boost iron absorption is quite an old one (Klavins et al., 1959; Klavins et al., 1962). These workers postulated that animal protein exerts an enhancing effect on the absorption of dietary iron.
Animal and vegetable foods were biosynthetically labeled with different radioisotopes of iron, and the incorporation of these radioiron forms into the circulating haemoglobin were taken as an index of absorption. It was found that veal increased, about 2-fold, the absorption of both maize and black bean iron. Subsequent studies revealed that certain amino acids, particularly cysteine, were responsible for the observed enhancement in iron absorption (Martinez- Torres et al., 1971).
This concept was further developed by further experiments which demonstrated that cysteine-containing peptides, rather than the free amino acid, were the causative agents for enhanced iron absorption. Two types of peptic digestion extracts were prepared from beef, in which the thiol groups were either oxidized (CYS -) or left untreated (CYS +). The absorption of radioiron mixed with maize was more than 2- fold greater when consumed along with the CYS+ extract than with the CYS- extract (P<0.005; Taylor et al., 1986).
In many respects, this seminal work led the way in the development of chelated organic trace minerals for human use, with the main emphasis being on facilitators of iron absorption. Today, several classes of compounds have been identified which assist iron absorption in humans (Table 3). Many studies have been published in recent years which show clear benefits in mineral status from dietary supplementation with chelated, or otherwise complexed trace elements.
In an effort to boost the mineral status of infants, many preparations are designed for inclusion in powdered or liquid milk formulae. In liquid formulae where the relative solubilities of copper, iron and zinc were examined, it was found that substituting organic forms of copper and iron in the mix resulted in an almost 3-fold increase in solubility while zinc solubilities were not improved. These results were reflected in enhanced absorption of copper and iron from the organic formula vs the inorganic formula, with the result for iron being statistically significant (P<0.05; Hendricks et al., 2001). Ferrous amino acid chelates are increasingly being used to combat Fe-deficiency anaemia through liquid milk fortification. Recent studies have shown that an iron bis-glycinate chelate, administered at a level of 3 mg/liter/day, was successful in restoring normal haemoglobin concentrations in a population of Brazilian children (Iost et al., 1998).
Table 3. Compounds indicated as facilitators of iron absorption1.
1Cremonesi et al., 2002.
Additional studies with this chelate involved 40 infants with Fe-deficiency anaemia being assigned to two groups, receiving either ferrous sulphate or the ferrous bis-glycinate chelate at a level of 5 mg Fe/kg bodyweight, administered via a syrup.
Significant increases in haemoglobin levels were observed in both groups but only the ferrous bisglycinate group displayed significant increases in plasma ferritin (Pineda and Ashmead, 2001). This material has also been successfully used to enhance iron absorption from foods enriched with phytate and polyphenols. In this case, absorption of iron from the iron glycinate was approximately twice that from ferrous sulphate (P<0.05; Layrisse et al., 2000).
Comparisons in the literature between the ferrous bis-glycinate chelate and other sources of organic iron are rather limited. One study examining the relative bioavailabilities of the iron bis-glycinate and ferrous ascorbate, when administered in drinking water, failed to demonstrate any significant difference between them (Olivares and Pizarro, 2001).
Other organic iron preparations used in the treatment of Fe-deficiency anaemia are designed with both enhanced bioavailability and better tolerability in mind. Unfortunately, orally administered iron compounds, particularly iron sulphate, may cause irritation and damage to the intestinal mucosa. Two types of polymeric ferric compounds have received particular attention in this regard, namely ferric polysaccharide complexes and iron protein succinylate (Raja et al., 2000). Positive therapeutic effects against Fe-deficiency anaemia without gastrointestinal side effects have been achieved with the iron protein succinylate (Cremonesi et al., 1984). A comparative study involving both organic iron forms indicated that both were well tolerated in children, but that the protein succinylate form was more effective in increasing iron stores (Haliotis and Papanastasiou, 1998).
Other organic iron complexes which have been proven to be effective in the treatment of anaemia, without adverse side effects, include ferric trimaltol (Reffitt et al., 2000).
From the available literature, one might be forgiven in thinking that the use of organic trace element supplementation in humans is restricted to the treatment of Fe-deficiency anaemia. Certainly, the majority of published work to date has focused on the treatment of Fe-deficiency anaemia but other interesting studies are beginning to emerge. For example, recent trials examined the therapeutic value of a broad-based nutritional supplement, predominantly composed of chelated trace minerals, on the treatment of bipolar disorder. The overall findings of these studies were that symptom reduction over the six-month duration of the trials ranged from 55% to 66% and the need for psychotropic medicines decreased by more than 50% (Kaplan et al., 2001). Other studies have shown low plasma levels of zinc and other minerals in people with mood and behavior disorders. For example, lower serum zinc levels were recorded in 48 unipolar depressed patients in comparison to 32 volunteers with normal mental health (Maes et al., 1994; Maes et al., 1997). In addition, a study which evaluated the copper and zinc levels in 135 assaultive, incarcerated males in comparison to controls found an inverse relationship between zinc and the severity of the behaviour, ranging from verbal assault to aggravated and violent assault; the lower the zinc, the greater the rate of violent behaviour (Walsh et al., 1997).
Such findings further assert the importance of certain trace minerals in diverse physiological processes and raise interesting possibilities for the amelioration of different ailments, both physical and psychological, through the enhancement of trace mineral status.
Uptake mechanisms for organic minerals in the gastrointestinal tract
There is clear evidence from the literature that differences in bioavailability exist between sources of the same trace metal, with metal proteinates and chelates proving superior to inorganic sources in many cases. If it can be accepted that uptake of metals from the intestine is the predominant factor influencing their bioavailability (Dreosti, 1993), then differences in uptake mechanisms or in the general presentation of the metal in organic versus inorganic form in the intestinal lumen are likely explanations for the differences noted.
The theory that metal amino acid chelates and proteinates utilize amino acid uptake and peptide uptake mechanisms rather than regular inorganic metal ion uptake pathways has become widely accepted (Ashmead et al., 1985; Ashmead, 1993).
The basic theory holds that the metal in question is protected within the complex in a chemically inert form. Consequently, the metal is not susceptible to the range of physicochemical factors which can adversely affect the efficient uptake of ‘unprotected’ mineral ions. Furthermore, it is hypothesized that the organic complex is absorbed intact through the intestinal mucosa, effectively carrying the metal along with it. The complex traverses the mucosal cell membrane, the mucosal cell and basement membrane, surviving, intact into the plasma.
If such an uptake mechanism did exist, it would help to explain numerous practical observations from studies on the relative bioavailability of organic versus inorganic sources in both animals and humans. The reductions noted in negative interactions between elements such as copper, iron and zinc when organic supplements are used agree well with this theory (Lowe et al., 1994; Hemken et al., 1996). For example, if copper in proteinated form utilizes a peptide uptake mechanism, it does not have to compete with zinc for regular metal ion uptake pathways.
Conclusive evidence for the existence of this kind of uptake mechanism has not been published, however. Although there is strong evidence that selenium in the form of selenomethionine enters the enterocyte via an electrogenic Na+- dependent neutral amino acid transport system in a manner which is indistinguishable from that of methionine (Vendeland et al., 1994), selenomethionine cannot be directly compared to structures such as ferrous bis-glycinate or zinc methionine. In selenomethionine, selenium replaces sulphur to become integrated into the molecular structure of methionine. It is inaccurate, therefore to draw an analogy between selenomethionine uptake and coordination complexes such as zinc methionine or iron glycine.
Some of the more recent studies in humans indicate that the processing of inorganic and organic minerals at the point of absorption may share more common elements than previously thought. For example, absorption studies on ferrous bis-glycinate reveal strong evidence that this substance uses the classic non-heme iron uptake pathway rather than the pathway for heme absorption (Pizarro et al., 2002). In this study, it was shown that iron from the bis-glycinate competes directly with ferrous sulphate for the non-heme uptake route in the intestinal lumen. Likewise, the finding that the intestinal absorption of iron (ferric) protein succinylate is inhibited by a ferrous chelator, ferrozine, indicates that reduction of ferric iron in this complex to ferrous iron must occur before absorption can take place.
This indicates a disruption of the complex and argues against uptake as an intact entity. The authors conclude that iron protein succinylate is absorbed by a mechanism involving digestion to release soluble, available ferric species which are reduced at the mucosal surface to provide ferrous iron for membrane transport into the enterocytes (Raja et al., 2000). Another organic iron complex, ferric trimaltol, has been shown to dissociate upon reaching the intestinal mucosa (Reffitt, 2000).
Today, several models exist for the uptake of trace elements from the gastrointestinal tract. The most popular of these is based upon concepts put forward by Conrad and Umbreit in 1993. As trace elements leave the acidic environment of the stomach and enter the alkaline environment of the small intestine, there is a tendency for certain elements, such as iron, copper, manganese and zinc, to undergo hydroxypolymerization (Powell et al., 1999). This results in the formation of large, insoluble metal hydroxide or metal oxide polymers that remain unavailable for uptake and utilization. For example, it has been estimated that in the pH range from 5.5 to 7.0, an increase of one pH unit causes a 30 to 45-fold decrease in aqueous Zn2+ ion concentration.
Similarly, an increase of one pH unit causes a 100- and 1,000-fold decrease in aqueous Fe2+ and Fe3+ ion concentration, respectively (Frossard et al., 2000). In the model of Conrad and Umbreit, ligands present in the structure of organic mineral products keep the metal soluble at alkaline pH until it can be donated to mucins. Mucins are large metal-binding glycoproteins that predominantly reside in the mucosally-adherent mucus layer. In the case of iron, mucin-bound iron subsequently crosses the mucosal cell membrane in association with integrins. Once inside the cell, a cytoplasmic iron-binding protein, dubbed ‘mobilferrin’, accepts the element and shuttles it to the basolateral surface of the cell where it is delivered to the plasma. The entire absorptive process is hypothesized to be driven by a cascade of differences in the binding constants of proteins so that iron moves from luminal mucin to cytoplasmic mobilferrin to plasma transferrin.
Whatever the mechanism, however, there is no doubt that the provision of minerals in organic form for human and animal use has been beneficial in terms of enhancing the bioavailability of essential trace elements, thereby contributing to improved health and performance. One can only look forward to further developments in this field, in areas such as the development of more effective ligands together with a more complete understanding of the relevant uptake mechanisms involved so that new approaches can be taken to influence them.
References
Araya, M., M. Olivares, F. Pizzaro, M. Gonzalez, H. Speisky and R. Uauy. 2003. Copper exposure and potential biomarkers of copper metabolism. BioMetals 16: 199-204.
Ashmead, H.D. 1993. Comparative intestinal absorption and subsequent metabolism of metal amino acid chelates and inorganic metal salts. In: The Roles of Amino Acid Chelates in Animal Nutrition. (H.D. Ashmead, ed). Noyes Publishers, New Jersey, pp 306-319.
Ashmead, H.D. 1996. Nutrition of the highproducing first parity sow. Proceedings of the XVII ANAPORC Symposium, Santiago de Compostela, Spain.
Ashmead, H.D. and D.J. Graff. 1982. Placental transfer of chelated iron. Proceedings of the International Pig Veterinary Society Congress, Mexico. p207.
Ashmead, H.D., D.J. Graff and H.H. Ashmead. 1985. Intestinal absorption of metal ions and chelates. Charles C. Thomas, Springfield, Illinois.
Baker, D.H. and C.B. Ammerman. 1995. Copper Bioavailability. In: Bioavailability of Nutrients for Animals. (C.B. Ammerman, D.H. Baker and A.J. Lewis, eds). Academic Press, San Diego. pp 127-156.
Close, B. 1999. Organic Minerals for Pigs: An Update. In: Biotechnology in the Feed Industry. (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, Nottingham, U.K., pp 51-60.
Conrad, M.E. and J.N. Umbreit. 1993. A concise review: iron absorption – the mucin-mobilferrin pathway. A competitive pathway for metal absorption. Am. J. Hematol. 42:67-73.
Cook, J.D. 1990. Adaptation in iron metabolism. Am. J. Clin. Nutr. 51:301-308.
Cremonesi, P., D. Strada, G. Galimberti and G. Sportoletti. 1984. Iron derivatives of modified milk protein. Arzneimittelforsch. Drug Res. 34:948-952.
Cremonesi, P., A. Acebron, K.B. Raja and R.J. Simpson. 2002. Iron absorption: biochemical and molecular insights into the importance of iron species for intestinal uptake. Pharmacol. Toxicol. 91:97-102.
DeMayer, E.M. 1989. Preventing and controlling iron deficiency anemia through primary health care. A guide for health administrators and programme managers. World Health Organization, Geneva.
Dreosti, I.E. 1993. Recommended dietary intakes of iron, zinc and other inorganic nutrients and their chemical form and identity. Nutrition 9:542-545.
Du, Z., R.W. Hemken and T.W. Clark. 1995. Copper proteinate may be absorbed in chelated form by lactating Holstein cows. In: Biotechnology in the Feed Industry. (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, Nottingham, U.K. pp 315-319.
Euromonitor Market Research. 2000. Vitamins and dietary supplements in the U.K.; Value of sales of vitamins and dietary supplements by sector, 1995-1999. Euromonitor International Inc. London.
Evans, R.M. and S.M. Hollenberg. 1988. Zinc fingers: gilt by association. Cell. 52(1):1-3.
Finch, C.A. and J.D. Cook. 1984. Iron deficiency. Am. J. Clin. Nutr. 39:471-477.
Frossard, E., M. Bucher, F. Machler, A. Mozafar and R. Hurrell. 2000. Potential for increasing the content and bioavailability of Fe, Zn and Ca in plants for human nutrition. J. Sci. Food Agric. 80:861-879.
Garrick, M.D., M.T. Nunez, M. Olivares and E.D. Harris. 2003. Parallels and contrasts between iron and copper metabolism. BioMetals 16:1-8.
Haliotis, F.A. and D.A. Papanastasiou. 1998. Comparative study of tolerability and efficacy of iron protein succinylate versus iron hydroxide polymaltose complex in the treatment of iron deficiency in children. Int. J. Clin. Pharmacol. Therap. 36:320-325.
Harvey, L. 2001. Mineral Bioavailability. Nutr.Food Sci. 31:179-182.
Hemken, R.W., T.W. Clark and Z. Du. 1993. Copper; Its role in animal nutrition. In: Biotechnology in the Feed Industry. (T.P. Lyons, ed). Alltech Technical Publications, Nicholasville, Kentucky, USA. pp 35-39.
Hemken, R.W., Z. Du and W. Shi. 1996. Use of proteinates to reduce competition from other trace minerals. In: Biotechnology in the Feed Industry. (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, Nottingham, United Kingdom, pp 91-94.
Hendricks, G.M., M.R. Guo and P.S. Kindstedt. 2001. Solubility and relative absorption of copper, iron and zinc in two milk-based liquid infant formulae. Int. J. Food Sci. Nutr. 52:419-428.
Henry, P.R. 1995. Manganese bioavailability. In: Bioavailability of Nutrients for Animals (C.B. Ammerman, D.H. Baker and A.J. Lewis, eds.). Academic Press, New York. pp 239-256.
Iost, C., J.J. Name, R.B. Jeppsen and H.D. Ashmead. 1998. Repleting hemoglobin in iron deficiency anemia in young children through liquid milk fortification with bioavailable iron amino acid chelate. J. Am. Coll. Nutr. 17:187-194.
Kaplan, B.J., J.S.A. Simpson, R.C. Ferre, C.P. Gorman, D.M. McMullen and S.G. Crawford. 2001. Effective mood stabilization with a chelated mineral supplement: an open-label trial in bipolar disorder. J. Clin. Psychiatry 62:936-944.
Kincaid, R.L., R.M. Blauwiekel and J.D. Conrath. 1986. Supplementation of copper as copper sulfate or copper proteinate for growing calves fed forages containing molybdenum. J. Dairy Sci. 69:160-163.
Kirchgessner, M. and E. Grassman. 1970. Dynamics of copper absorption. In: Trace Element Metabolism in Animals (C.F. Mills, ed.). Livingston, Edinburgh. pp 227-242.
Klavins, J.V., T.D. Kinney and N. Kaufman. 1959. Iron absorption in rats fed a protein-free diet. Am. J. Pathol. 35:690-691.
Klavins, J.V., T.D. Kinney and N. Kaufman. 1962. The influence of dietary protein on iron absorption. Br. J. Exp. Pathol. 43:172-180.
Layrisse, M., M.N. Garcia-Casal, L. Solano, M.A. Baron, F. Arguello, D. Llovera, J. Ramirez, I. Leets and E. Trooper. 2000. Iron bioavailability in humans from breakfasts enriched with iron bisglycine chelate, phytates and polyphenols. J. Nutr. 130:2195-2199.
Lee, D-Y., J. Schroder and P.T. Gordon. 1988. Enhancement of copper bioavailability in the rat by phytic acid. J. Nutr. 118:712-716.
Lowe, J.A., J. Wiseman and D.J.A. Cole. 1994. Absorption and retention of zinc when administered as an amino acid chelate in the dog. J. Nutr. 124:2572s–2574s.
Maes, M., P.C. D’Haese and S. Scharpe. 1994. Hypozincemia in depression. J. Affect. Disord. 31:135-140.
Maes, M., E. Vandoolaeghe and H. Neels. 1997. Lower serum zinc in major depression is a sensitive marker of treatment resistance and of the immune/inflammatory response in that illness. Biol. Psychiatry 42: 349-358.
Martinez-Torres, C. and M. Layrisse. 1971. Iron absorption from veal muscle. Am. J. Clin. Nutr. 21:1175-1183.
MAFF. 1999. National food survey. Annual Report on Food Expenditure, Consumption and Nutrient Intakes. The Stationery Office, London, p. 153.
McDowell, L.R. 1992. Copper and molybdenum. In: Minerals in Animal and Human Nutrition (T.J. Cunha ed). Academic Press, San Diego. pp 176-204.
Miles, R.D. 1998. The influence of Eggshell 49 on shell quality of hens grouped by their shell quality. Poult. Sci. 77(Supplement 1):43.
Olivares, M. and F. Pizarro. 2001. Bioavailability of iron bis-glycinate chelate in water. Arch.Latinoam Nutr. 51:22-25.
Pineda, O. and H.D. Ashmead. 2001. Effectiveness of treatment of iron deficiency anemia in infants and young children with ferrous bis-glycinate chelate. Nutrition 17:381-384.
Pizarro, F., M. Olivares, E. Hertrampf, D.I. Mazariegos, M. Arredondo, A. Letelier and V. Gidi. 2002. Iron bis-glycine chelate competes for the nonheme-iron absorption pathway. Am. J. Clin. Nutr. 76:577-581.
Powell, J.D., R. Jugdaohsingh and R.P.H. Thompson. 1999. The regulation of mineral absorption in the gastrointestinal tract. Proc. Nutr. Soc. 59:147-153.
Raja, K.B., S.E. Jafri, D. Dickson, A. Acebron, P. Cremonesi, G. Fossati and R.J. Simpson. 2000. Involvement of iron (ferric) reduction in the iron absorption mechanism of a trivalent iron-protein complex (iron protein succinylate). Pharmacol. Toxicol. 87:108-115.
RealEat surveys conducted by Gallup 1984-1989.
Poll summaries at www.vegsoc.org/info/realeat. html Reffitt, D.M., T.J. Burden, P.T. Seed, J.Wood, R.P.H. Thompson and J.J. Powell. 2000. Assessment of iron absorption from ferric trimaltol. Ann. Clin. Biochem. 37:457-466.
Rucker, R.B., B. Lonnerdal and C.L. Keen. 1994. Intestinal absorption of nutritionally important trace elements. In: Physiology of the Gastrointestinal Tract Third Edition. (L.R. Johnson ed). Raven Press, New York. pp 2183-2202.
Schultze, M.O., C.A. Elvehjem and E.B. Hart. 1934. The availability of copper in various compounds as a supplment to iron in hemoglobin formation. J. Biol. Chem.106:735-740.
Schultze, M.O., C.A. Elvehjem and E.B. Hart. 1936. Further studies on the availability of copper from various sources as a supplement to iron in hemoglobin formation. J. Biol. Chem. 115:453-457.
Suchy, P.E., P. Strakova and P. Simon. 1998. Effect of applications of various forms of zinc on gonad development in breeding cocks. Czech J. Anim. Sci. 43:343-348.
Taylor, P.G., M.A.C. Martinez-Torres, E.L. Romano and M. Layrisse. 1986. The effect of cysteinecontaining peptides released during meat digestion on iron absorption in humans. J. Clin. Nutr. 43:68- 71.
Underwood, E.J. 1997. Trace elements in human and animal nutrition. Academic Press, New York. Vallee, B.L. and K.H. Falchuk. 1993. The biological basis of zinc physiology. Physiol. Rev. 73:79-118.
Vendeland, S.C., J.T. Deagan, J.A. Butler and P.D. Whanger. 1994. Uptake of selenite, selenomethionine and selenate by brush border membrane vesicles isolated from rat small intestine. BioMetals 7:305-312.
Walsh, W.J., H.R. Isaacson and F. Rehman.1997. Elevated blood copper/zinc ratios in assaultive young males. Physiol.Behav. 62:327-329.
Wardle, C. 1977. Changing food habits in the UK: an assessment of the social, technological, economic and political factors which influence dietary patterns. Earth Resources Research, London, pp 71-90.
Wedekind, K.J., A.E. Hortin and D.H. Baker. 1992. Methodology for assessing zinc bioavailability: efficacy estimates for zinc methionine, zinc sulphate and zinc oxide. J. Anim. Sci. 70:178-184.
Xin, Z., D.F. Waterman, R.W. Hemken and R.J. Harmon. 1991. Effects of copper status on neutrophil function, superoxide dismutase and copper distribution in steers. J. Dairy Sci. 74:3078- 3082.
Author: RONAN POWER North American Biosciences Center, Alltech Inc., Nicholasville, KY, USA
Related topics:
Authors:

Alltech
Recommend
Comment
Share

Would you like to discuss another topic? Create a new post to engage with experts in the community.
Featured users in Pig Industry






