Health of dairy cattle
Selenium, immune functions and health of dairy cattle
Health of the dairy cow has been the subject of intense research over the years. Several conditions, including pathogens and poor nutrition, invoke diverse mechanisms that facilitate poor growth and lower productivity, and when unrestrained might be implicated in the pathogenesis of diseases.
The impairment of dairy cow immunity is one of the fundamental reasons why these diseases occur (Mehrzad et al., 2001; Burvenich et al., 2003; Paape et al., 2003) and disruption of normal physiological processes is responsible for lowered productivity. For example, an increased vulnerability of dairy cows to intramammary infections (IMIs) has been attributed to a low status of selenium (Smith et al., 1984; 1987; Malbe et al., 1995; 2003; Weiss and Hogan, 2005).
Selenium is an essential trace element that serves several biological purposes as an integral component of a variety of proteins or enzymes known as selenoproteins. Selenium functions in the body as an antioxidant, in thyroid hormone metabolism, redox reactions, improves reproduction efficiencies and immune functions, and has anticarcinogenic properties (Rayman, 2000; Spears, 2000). It is also insulin mimetic (Stapleton, 2000).
Selenium deficiency therefore exerts profound effects on the dairy industry through lowered health status and productivity. Forages for dairy cows often do not contain adequate supplies of selenium. Therefore supplementation of selenium is now a common practice in many countries. Notwithstanding the beneficial effects of adequate amounts of selenium in dairy nutrition, the exact mechanism of this positive influence is not yet fully understood.
In this review, we will examine the influence of selenium on immune functions of the cow, highlight recent advances and discuss the physiological outcomes and consequences of selenium supplementation.
Selenium and selenoproteins
Selenium is a trace element that was first discovered in the late 19th century and considered toxic to plants and animals in the period between the 1930s and 1950s. Biomedical interest in selenium started in the late 1950s with reports that it was an essential trace element for mammals and also a component of the enzyme, glutathione peroxidase (GSH-Px) (Schwarz and Foltz, 1957; Flohé et al., 1973).
GSH-Px is an enzyme that provides antioxidative defense in mammals by converting hydrogen peroxide to water and lipid hydroperoxides to the corresponding alcohols. Selenium is integrated into GSH-Px as a selenocysteine residue, and is responsible for the catalytic efficiency of the enzyme (Forstrom et al., 1978; Wendel et al., 1978; Rocher et al., 1992; Aumann et al., 1997).
Selenium exerts its physiological influences as an integral part of several proteins known as selenoproteins, in the form mostly of selenocysteine, now recognized as the 21st amino acid in the genetic code (Stadtman, 1996; Hatfield and Gladyshev, 2002; Hatfield et al., 2006). The UGA codon, one of the so-called stop codons of the genetic code, now has the added function of signaling the incorporation of selenocysteine into selenoproteins after signaling the termination of normal protein synthesis. Thus, selenium is the raw material for production and smooth functioning of selenoproteins.
Although selenium is specifically incorporated into selenoproteins as selenocysteine, it can unspecifically join proteins as selenomethionine, which acts as a biological pool for selenium (Suzuki and Ogra, 2002). The important and varied physiological functions of selenoproteins now make selenium an essential dietary trace element in human and bovine nutrition.
Since the discovery of the first selenoprotein, about 50 mammalian proteins have been described, out of which about 25 are found in humans and 24 in rodents, and also about 30 in prokaryotes (reviewed by Köhrle et al., 2000; Kryukov et al., 2003). The first selenoprotein reported for bovine (brain) was selenoprotein P12 (SelP 12) (Saijoh et al., 1995). Lindmark- Månsson and Åkesson (2001) later demonstrated the presence of plasma GSH-Px (GSH-Px3) in both human and bovine milk. Recently, the mRNA of five more mammalian selenoproteins has been found in bovine mammary tissue (Bruzelius et al., 2007). Some of the selenoproteins (enzymes) have been assigned physiological functions but the mode of action of a good number of them is yet to be identified (Table 1).
Selenoproteins are found in many mammalian tissues and organs (Table 1) and their availability is regulated by the presence of selenium. Under a low selenium status, expression of some selenoproteins is prioritized whereas some, like GSH-Px isoforms, are strongly influenced (Brigelius-Flohé, 1999; Romanowska et al., 2007). The distribution and concentration of selenoproteins in animal tissues depends on dietary sources of the mineral (WHO, 1987; Gibson, 1990). On the other hand, the selenium content of plants and plant parts like grains and seeds is largely influenced by the selenium content of the soils in which they grow (WHO, 1987; Gibson, 1990). For dairy cattle, dietary selenium is mostly derived from plant sources.
Selenium and disease incidence in cattle
The influence of selenium on disease incidence and on immune functions is multifactorial and particular circumstances determine which systems are affected (Arthur et al., 2003).
Goff (2006) suggested that a strong correlation exists between malnutrition and the immune system, consequently causing disease conditions in cattle. Selenium appears to stimulate the immune system to boost host resistance to infection. In fact, selenium influences both the innate and the acquired immune systems (Spallholz, 1990; McKenzie et al., 2001). Selenium over-availability or under-availability has been shown to alter normal physiological processes, often leading to disease conditions, which implies that selenium as a trace element must be present in biological systems only at a certain threshold for proper functioning.
The first observed ill effects of selenium came when animals and humans alike consumed seleniferous plants and plant parts (grew in areas with high soil selenium content) and produced severe toxic symptoms such as loss of hoofs (in animals), changes in skin (red, swollen, and blistered), brittle, discolored and eventual loss of hair and nails, and nervous system failure (in humans) (Krehl, 1970; Yang et al., 1983).
Selenium deficiency on the other hand has been implicated in several disease conditions in cattle, such as ill-thrift, reduced growth rate, retained placenta and abortion, impaired fertility, cystic ovaries, metritis, white muscle disease in calves, mastitis and nutritional muscular dystrophy (Smith et al., 1984; Koller and Exon, 1986; Hemingway, 1999; 2003; Pehrson, 1993; Ndiweni and Finch, 1995; Kommisrud et al., 2005; Enjalbert et al., 2006). Augmentation of the selenium status of cattle has been shown to improve, reverse or prevent most of these conditions. Table 2 presents a summary of the effects of level of blood selenium status on cow health parameters.
Table 1. Some mammalian selenoproteins, their functions and tissue distribution.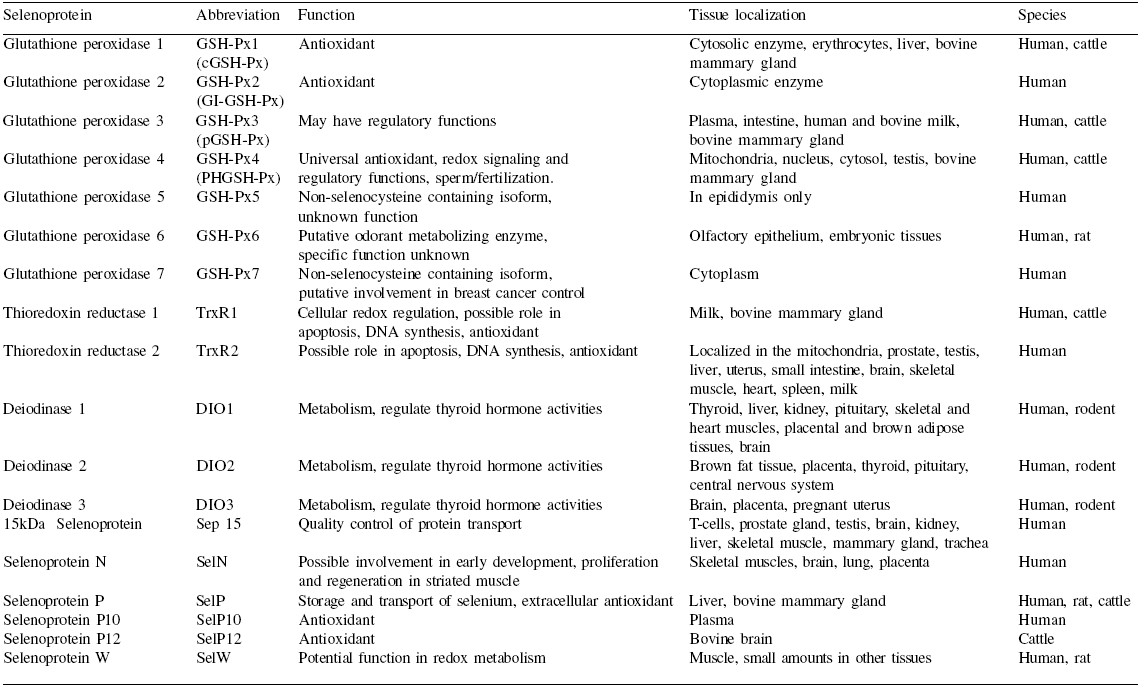
To enlarge the image, click here
References: Holben and Smith, 1999; Köhrle et al., 2000; Gromer et al., 2005; Bruzelius et al., 2007.
Table 2. Blood selenium level and health parameters of cattle.
SELENIUM AND SOMATIC CELL COUNTS
Milk somatic sell counts (SCC) have over the years been used as a primary indicator of mastitis (or disease incidence) and milk quality in dairy herds. Several controlled trials and field studies have associated deficiencies in selenium with increased incidence and severity of IMIs, clinical mastitis cases, and higher SCC in individual cows and bulk tank milk (Erskine et al., 1987; Smith et al., 1987; Weiss et al., 1990; Hemingway, 1999). Smith et al. (1987) supplemented the diets of first lactation heifers with 0.3 mg Se/kg DM and reported a reduction in prevalence of infected quarters by 42%, clinical mastitis cases by 32.1%, and a significant reduction in SCC compared with nonsupplemented heifers.
Studies that examined the relationships among dietary and serum selenium concentration and bulk tank SCC associated high serum selenium concentrations with lower bulk tank SCC and lower rates of clinical mastitis (Erskine et al., 1987; Weiss et al., 1990). In the study by Erskine et al. (1987), higher GSH-Px activity was found in the herd with low SCC. Conversely, Norwegian herds with high selenium status were found to have higher frequencies of treatment for clinical mastitis and higher milk SCC than herds with low selenium status (Ropstad et al., 1987).
On the other hand, Erskine et al. (1990) found no difference in SCC from low selenium status cows (received 0.04 mg Se/kg DM) and high selenium status cows (received 2.04 mg Se/kg DM) after intracisternal challenge with Staphylococcus aureus. They, however, noted that peak SCC was reached much earlier in the low selenium status cows.
SELENIUM AND NEONATE IMMUNITY
At birth, a neonate is rapidly exposed to large numbers of potential pathogens. Hence, the ruminant mammary gland is responsible for providing protective immunity to neonates and for defending itself from invading pathogens (Kehrli and Harp, 2001).
Selenium inadequacy during this period leading up to calving is considered critical, because most disorders are thought to be selenium-responsive (retained placenta, mastitis, neonatal mortality, reduced milk production, impaired reproductive efficiency) (Gerloff, 1992; Hemingway, 2003). As demonstrated by Ndiweni and Finch (1995), low selenium status of cows may affect the levels of important immune cells in colostrum and consequently put calf health at risk.
Furthermore, several authors have confirmed that selenium supplementation resulted in a higher concentration of selenium in milk of suckler cows and also improved the selenium status of their calves (Pehrson et al., 1999; Gunter et al., 2003; Weiss and Hogan, 2005). High GSH-Px activities were also reported in calves from cows fed supplemental selenium (Gunter et al., 2003).
Potential mechanisms of action of selenium
Available information indicates that selenium consumption at safe levels affects positively the cow’s biological systems leading to improvements in health and reproduction parameters. The mechanisms of action of selenium in achieving these goals are mirrored in studies showing enhanced effects on the general immune system, the antioxidant system and in specific activities such as anti-pathogenic roles and influence on neutrophil functions.
SELENIUM AND THE DAIRY COW IMMUNE SYSTEM
Selenium availability is essential for an optimum response after pathogen attack in cattle, although the exact mechanisms of action are still being investigated. Selenium influences both the acquired and adaptive immune system of cows including antibody production, cell proliferation, cytokine production and neutrophil function (Larsen, 1993; Ndiweni and Finch, 1995; McKenzie et al., 1998).
Studies in humans indicate that selenium deficient lymphocytes were less able to proliferate in response to mitogens and also neutrophil chemotaxis was impaired (Turner and Finch, 1991; Beckett et al., 2003). Selenium deficiency also affects the humoral system whereby, IgG and IgA titers were decreased in rats and IgG and IgM in humans (Arvilommi et al., 1983; Turner and Finch, 1991; Beckett et al., 2003).
In cows suffering from selenium deficiency, Boyne and Arthur (1986) noted a decrease in the ability of neutrophils to kill phagocytosed Candida albicans. Also, a lack of selenium lowered the production of leucotrienes of polymorphonuclear leukocytes, which led to lowered chemotaxis of neutrophils (Aziz and Klesius, 1986).
Several reports associated increased concentration of blood selenium in cows with decreased rate in infections, including subclinical mastitis, clinical mastitis and infections by Actinomyces pyogenes and Corynebacterium spp. (Jukola et al., 1996; Ranjan et al., 2005). Furthermore, many epidemiological studies have also revealed positive association of selenium supplementation with cow udder health (Weiss et al., 1990; Hogan et al., 1993; Malbe et al., 1995; Kommisrud et al., 2005).
In other findings, both the severity and duration of infections as well as somatic cell count are associated with selenium status of animals (Smith et al., 1987; Erskine et al., 1990; Weiss et al., 1990; Hemingway, 1999).
THE ANTIOXIDANT EFFECTS OF SELENIUM
Beneficial health effects have been consistently associated with selenium supplementation in dairy cows (Pehrson et al., 1999; Rayman, 2004; Malbe et al., 2006). It is generally understood that these benefits are achieved through the action of selenium as a component of selenoproteins.
The GSH-Px and thioredoxin reductase groups of selenoproteins (Table 1) catalyze reduction of peroxides that can damage cells and tissues. Being a part of these proteins, selenium has the capacity to affect oxidative processes in the system and is considered an antioxidant nutrient. These antioxidants protect cells from oxidative damage from free radicals and peroxides (Rotruck et al., 1973). The activity of these enzymes is directly related to the concentration of selenium in the diet. In cattle, Bruzelius et al. (2007) demonstrated for the first time the activity of GSH-Px and thioredoxin reductase activity in bovine mammary tissue, which was influenced by selenium status.
This is an indication that selenium status of the mammary gland is an important regulator of selenoprotein expression and activities. Furthermore, several studies have demonstrated that oxidative stress of the cow contributes to lowering immunity and increases the risk of retained fetal membranes as well as mastitis (Hemmingway, 1999). Higher activity of GSH-Px has also been reported in the erythrocytes of cows receiving supplemental selenium compared with non-supplemented individuals (Ortman and Pehrson, 1999). Selenium therefore has a positive antioxidant effect in cattle.
SELENIUM-DEPENDENT ANTI-PATHOGENIC ACTIVITIES
Reports in cattle show that increased activity of GSH-Px, the result of selenium supplementation, inhibited growth of mastitis pathogens in cow’s milk (Ali-Vehmas et al., 1997; Malbe et al., 2003; 2006). Ali-Vehmas et al. (1997) observed that selenium supplementation of dairy cattle strengthens the inflammatory response to IMIs by inducing a change in whey, which either inhibited or restricted the growth of mastitis pathogens.
Similarly, 4 mg of supplemental selenium played an important role in maintaining quarter milks pathogen free (Malbe et al., 2003). A recent finding by Malbe et al. (2006) has established the presence of a selenium-dependent anti-pathogenic activity in cow’s milk.
In their study, selenium supplementation increased GSH-Px activity, which accounted for lower somatic cell counts and inhibited Staphylococcus aureus growth, especially when the activity level exceeded 4 μkat/g Hb. Bruzelius et al. (2007) also showed increased activity of cellular GSH-Px1, GSH-Px3, and selenoprotein P with increased mRNA abundance in the mammary gland, which is in line with the findings of Gierus et al. (2002) who demonstrated increased GSH-Px3 activity with selenium supplementation.
These findings give an insight into the mechanism of selenium action in udder defense, which reiterates the importance of adequate amounts of selenium in the diet of the dairy cow.
ROLE OF SELENIUM IN BOVINE NEUTROPHIL FUNCTION
Neutrophils are the first line of cellular defense against the aggression of invading pathogens. Thus, their recruitment to the site of pathogen attack will support defense mechanisms that favor reduction of the incidence of disease. The positive effect of selenium supplementation on the health of dairy cows has necessitated the elucidation of the role of selenium in neutrophil function. Neutrophils release free radicals when they reach sites of infection to take part in the killing of pathogens.
Oxygen free radicals such as superoxide anion, hydrogen peroxide and hydroxyl radical are generated in a process known as oxidative or respiratory burst. Although selenium deficiency has not been shown to affect neutrophil numbers in a range of species, it is clear that certain aspects of their function are affected (Turner et al., 1991).
In cattle, neutrophils collected from cows fed supplemental selenium diets had increased intracellular kill of bacteria, enhanced viability and reduced extracellular hydrogen peroxide concentration when compared with neutrophils from cows fed unsupplemented diets (Gyang et al., 1984; Hogan et al., 1990; Grasso et al., 1990).
The role of selenium in these studies was associated with the activity of GSH-Px in the neutrophils. Under deficient conditions, a lack of/or decreased GSH-Px activity seems to reduce the ability to kill bacteria. On the other hand, increased GSH-Px activity in the presence of selenium enabled the continued production of radicals and bacterial clearance. Furthermore, Cebra and co-workers (2003) reported that neutrophils from postparturient dairy cows with higher blood concentrations of selenium appear to have greater potential to kill microbes. Reported positive correlation between GSH-Px activity in the bovine mammary gland and milk and health benefits supports the positive role of selenium on neutrophil function in dairy cattle health.
Considering the potential mechanisms of action of selenium, we hypothesize that selenium adequacy leads to production and increased abundance of certain selenoproteins (depending on the system and conditions) that increase the ability of neutrophils to kill pathogens.
In addition, selenium adequacy will prolong the lifespan of neutrophils, thus enhancing their function. The antioxidant role of most selenoproteins already provides suitable conditions for neutrophil function. However, other unknown mechanisms may also be involved. Although several mammalian selenoproteins are known, their specific roles in cattle have not been investigated. Experiments are on-going or planned in our laboratory to test our hypotheses and characterize and determine the role of several selenoproteins in the bovine system.
Selenium and vitamin E
The positive action of selenium in biological systems is thought to be supported by vitamin E. Vitamin E is a fat soluble vitamin located in membranes where it protects polyunsaturated fatty acids from peroxidation. Like selenium, vitamin E also plays an oxidative role by protecting neutrophils from the destructive action of toxic oxygen molecules. Deficiencies in both nutrients have been implicated in impaired bactericidal activity of neutrophils and increased mastitis (Hogan et al., 1993).
Consequently, several reports in the past decades have examined the role of vitamin E and selenium on bovine health (Hogan et al., 1990; 1993; Ndiweni and Finch, 1995; Jukola et al., 1996; Sivertsen et al., 2005). The general consensus is that adequate amounts of these nutrients enhance the ability of bovine neutrophils to phagocytize pathogens and also enhance the functions of mammary gland macrophages and peripheral blood lymphocytes and consequently improve health, while their deficiency is associated with lower productivity and increased incidence of diseases.
Although the separate supplementation of these nutrients has resulted in the same positive effects, Sergerson et al. (1981) are of the opinion that, when used together, their effects are even more efficacious. However, Ndiweni and Finch (1995; 1996) found no evidence of additive or synergistic effects of vitamin E and selenium supplementation in health parameters in cattle.
Manipulating the selenium status of the cow and its milk, and bioavailability
Selenium deficiency can be corrected by supplementation. As a consequence of several analyses of selenium deficiency in livestock, humans and agricultural land, selenium supplementation of livestock feed or use of mineral fertilizers on crop lands is now a common practice in many parts of the world. In particular, the positive effects of selenium supplementation on the immune status and reproductive parameters of dairy cows have made the mineral a normal constituent of their rations.
Selenium supplementation is therefore necessary to maintain the selenium status of dairy cows and also increase the selenium concentration of milk (Knowles et al., 1999; Pehrson et al., 1999; Givens et al. 2004; Heard et al. 2004; Juniper et al., 2006). Selenium supplementation of dairy cows is also of significance in human nutrition since the main sources of human selenium supplies are from products of animal origin like meat and milk.
Selenium supplementation in cattle feed is usually in the form of organic (selenized yeast) or inorganic (sodium selenite or sodium selenate) selenium. Several studies have examined the effects of selenium sources on bioavailability and demonstrated varying capacities of the two forms at affecting dairy cow selenium status (in blood) and selenium content of milk (Knowles, et al., 1999; Weiss and Hogan, 2005; Juniper et al., 2006).
In general, the organic form of selenium shows higher capacity of assimilation and bioavailability, and in increasing the blood and milk selenium content when compared with the inorganic form (Ortman and Pehrson, 1999; Knowles et al., 1999; Mahan, 2000; Givens et al. 2004; Heard et al. 2004; Weiss and Hogan, 2005; Juniper et al., 2006). In a review by Weiss (2003) and subsequent studies (Givens et al., 2004; Heard et al., 2004; Juniper et al., 2006), the relative effect of organic selenium on the increase of milk selenium lies in the range of 34 to 90%.
Optimal dosage and toxic effects of selenium supplementation
The level of selenium supplementation in dairy cow diets varies from country to country and is mainly determined by the selenium content of feedstuffs, as influenced by soil selenium status. In Canada and the United States, both organic and inorganic sources of selenium can be supplemented at 0.3 mg/kg of DM. In the European Union, inorganic selenium is fed to cows at a recommended level of 0.5 mg/kg of DM (Ministry of Agriculture, Fisheries and Food, 2000) and was initially the only authorized form of selenium.
Recently, one source of organic selenium, Sel-Plex® (Alltech Inc.) has been approved for use in all livestock species in the European Union (EU regulation 1750/ 2006). In the US, Sel-Plex® first received approval for use in chicken diets in the year 2000, followed by turkey, swine, beef, dairy and equine feeds and most recently dog diets.
Since selenium was originally known to be toxic, it is imperative that care be taken not to reach toxic levels during supplementation. Rate of supplementation in most trials does not consider the selenium concentration in the other feed ingredients, implying that the final selenium intakes of animals is usually more than current recommended levels. However, little data exist to suggest toxic effects even when selenium was included at levels as high as 50 mg/d for 90 days or 100 mg/day for 28 days in diets of Holstein cows (Ellis et al., 1997).
Givens at al. (2004), as in most studies, observed increasing advantage of selenium inclusion levels on milk selenium concentration with the best results at a rate of 1.14 mg Se/kg as Sel-Plex® selenium yeast. These results demonstrate that current recommended levels of inclusion should be revisited in order to set the limits for favorable performance and to avoid toxicity. Since blood selenium concentration is a fairly good variable to measure individual or herd selenium status, Hogan et al. (1993) recommended its use in establishing selenium status. Their recommendation follows that blood selenium concentration should be at least 0.2 μg/ mL, but should not exceed 1 μg/mL.
On the other hand, Braun et al. (1991) considered blood selenium levels of 0.08 to 0.3 μg/mL as normal and 0.03 to 0.07 μg/mL as inadequate for cattle. To ensure that herd selenium intakes are within safe limits, it is imperative that intakes should not exceed the toxic threshold of selenium supplementation for dairy cows. Stating the selenium concentration on feed tags and routinely measuring concentration level in forages, hay and other feed materials may be one way of ensuring safe and favorable levels of inclusion and intakes.
Other unexplored effects of selenium in dairy cattle
A further area where selenium seems to play a role, but not yet examined in dairy cattle, is in apoptosis. The ultimate goal of a cell is to perform its function to the best of its ability in the maintenance of tissue homeostasis. Apoptosis is the process of deliberate life relinquishment by a cell in an organism. It is also commonly referred to as ‘cell suicide.’
In humans, an insufficient amount of apoptosis leads to uncontrolled cell proliferation, such as cancer whereas excessive apoptosis causes cell loss diseases such as ischemic damage (shortage of blood supply to an organism). Data in 1996 indicated that dietary supplementation of 200 μg of selenium in diets resulted in a decreased incidence of human prostate, lung and colorectal cancers (Clark et al., 1996).
The precise role of selenium in modulating cancers is much debated. Some authors are of the opinion that small molecules of selenium metabolites selectively promote apoptosis in transformed prostate epithelium (Jiang et al., 2004; Ip et al., 2000) while others believe that selenium supplementation prevents DNA damage that could lead to cell transformation by increasing levels of antioxidant selenoproteins (Diwadkar- Navsariwala and Diamond, 2004; Lu and Jiang, 2005).
A recent study with transgenic mouse models showed that selenoprotein deficient mice exhibited accelerated development of lesions associated with prostate cancer progression thus suggesting that selenium may function in cancer prevention by modulating the levels of selenoproteins (Diwadkar- Navsariwala et al., 2006). Another finding demonstrates that selenoprotein expression and selenium metabolism are regulated at multiple levels in prostate cells (Rebsch et al., 2006).
Selenium may therefore play a positive role in mediating useful apoptosis necessary for normal functioning of cells. Selenium’s role in this regard, is however, yet to be established in cattle.
| Conclusions Selenium, and consequently selenoproteins, are vital players in dairy cow health and eventually human health through the transfer of selenium into milk and meat. Existing evidence clearly shows a positive influence of adequate dietary selenium supplementation on the general immune system of the cow, which effects are further transferred to calves. Specific aspects of the immune function, like the ability of neutrophils to kill invading pathogens, are enhanced when selenium content of blood is in adequate amounts. The general role of selenium as an antioxidant in ensuring the smooth functioning of the biological system of the cow necessitates adequate supplementation. In supplementation, however, attention needs to be paid to the coordinate effect of selenium and vitamin E plus the form of the selenium. Finally, we suggest the need to characterize and define the roles of bovine selenoproteins, revisit supplementation levels and also explore the role of selenium in mediating apoptosis in the bovine system. Only when all the selenoproteins are identified and characterized can we fully understand the role of selenium in immunity. |
References
Authors: ALOYSIUS E. IBEAGHA1, EVELINE M. IBEAGHA-AWEMU1, JALIL MEHRZAD2 and XIN ZHAO1Ali-Vehmas, T., M. Vikerpuur, W. Fang and M. Sandholm. 1997. Giving selenium supplements to dairy cows strengthens the inflammatory response to intramammary infection and induces a growth-suppressing effect on mastitis pathogens in whey. J. Vet. Med. 44:559-571.
Arthur, J.R., R.C. McKenzie and G.J. Beckett. 2003. Selenium in the immune system. J. Nutr. 133:1457-1459.
Arvilommi, H., K. Poikonen, I. Jokinen, O. Muukkonen, L. Rasanen, J. Foreman and J. Huttunen. 1983. Selenium and immune functions in humans. Infect. Immun. 41:185-189.
Aumann, K.-D., N. Bedorf, R. Brigelius-Flohé, D. Schomburg and L. Flohé. 1997. Glutathione peroxidase revisited. Simulation of the catalytic cycle by computer assisted molecular modeling. Biomed. Environ. Sci. 10:136-155.
Aziz, E. and P.H. Klesius. 1986. Effect of selenium deficiency on caprine polymorphonuclear leucocyte production of leucotriene B4 and its neutrophil chemotactic activity. Am. J. Vet. Res. 47:426-428.
Beckett, G.J., J.R. Arthur, S.M. Miller and R.C. McKenzie. 2003. Selenium, immunity and disease. In: Dietary Enhancement of Human Immune Function (D.A. Hughes, A. Bendich and G. Darlington, eds). Humana Press, Totowa, NJ, USA, pp. 217-240.
Boyne, R. and J. R. Arthur. 1986. The response of selenium deficient mice to Candida albicans infection. J. Nutr. 116:816-822.
Braun, U., R. Forrer, W. Furer and H. Lutz. 1991. Selenium and vitamin D in blood sera of cows from farms with increased incidence of disease. Vet. Record 128:543- 547.
Brigelius-Flohé, R. 1999. Tissue-specific functions of individual glutathione peroxidases. Free Rad. Biol. Med. 27:951-965.
Bruzelius, K., T. Hoac, R. Sundler, G. Önning and B. Åkesson. 2007. Occurrence of selenoprotein enzyme activities and mRNA in bovine mammary tissue. J. Dairy Sci. 90:918-927.
Burvenich, C., V. Van Merris, J. Mehrzad, A. Diez-Fraile and L. Duchateau. 2003. Severity of E. coli mastitis is mainly determined by cow factors. Vet. Res. 34:521- 562.
Cebra, C.K., J.R. Heidel, R.O. Crisman and B.V. Stang. 2003. The relationship between endogenous cortisol, blood micronutrients, and neutrophil function in postparturient Holstein cows. J. Vet. Intern. Med. 17:902-907.
Clark, L.C., G.F. Combs Jr, B.W. Turnbull, E.H. Slate, D.K. Chalker, J. Chow, L.S. Davis, R.A. Glover, G.F. Graham, E.G. Gross, A. Krongrad, J.L. Lesher, Jr., H.K. Park, B.B. Sanders, Jr., C.L. Smith and J.R. Taylor. 1996. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention of Cancer Study Group. JAMA 276:1957-1963.
Diwadkar-Navsariwala, V. and A.M. Diamond. 2004. The link between selenium and chemoprevention: a case for selenoproteins. J. Nutr. 134:2899-2902.
Diwadkar-Navsariwala, V., G.S. Prins, S.M. Swanson, L.A. Birch, V.H. Ray, S. Hedayat, D.L. Lantvit and A.M. Diamond. 2006. Selenoprotein deficiency accelerates prostate carcinogenesis in a transgenic model. PANS 103:8179-8184.
Ellis, R.G., T.H. Herdt and H.D. Stowe. 1997. Physical, hematological, biochemical, and immunological effects of supranutrional supplementation with dietary selenium in Holstein cows. Am. J. Vet. Res. 58:760-764.
Enjalbert, F., P. Lebreton and O. Salat. 2006. Effects of copper, zinc and selenium status on performance and health in commercial dairy and beef herds: retrospective study. J. Anim. Physiol. Anim. Nutri. 90:459-466.
Erskine R.J., R.J. Eberhart, L.J. Hutchinson and R.W. Scholz. 1987. Blood selenium concentrations and glutathione peroxidase activities in dairy herds with high and low somatic cell counts. J. Am. Vet. Med. Assoc. 190:1417-1421.
Erskine R.J., R.J. Eberhart and R.W. Scholz. 1990. Experimentally induced Staphylococcus aureus mastitis in selenium-deficient and selenium-supplemented dairy cows. Am. J. Vet. Res. 51:1107-1111.
Flohé, L., W. Günzler and H.H. Schock. 1973. Glutathione peroxidase: a selenoenzyme. FEBS Lett. 32: 132-34.
Forstrom J.W., J.J. Hachowski and A.L. Tappel. 1978. Identification of the catalytic site of rat liver glutathione peroxidase as selenocysteine. Biochemistry 17:2639-2644.
Gerloff, B.J. 1992. Effect of selenium supplementation on dairy cattle. J. Anim. Sci. 70:3934-3940.
Gibson, R.S. 1990. Principles of Nutritional Assessment. Oxford University Press, New York, NY, USA.
Gierus, M., F.J. Schwarz and M. Kirchgessner. 2002. Selenium supplementation and selenium status of dairy cows fed diets based on grass, grass silage or maize silage. J. Anim. Physiol. Anim. Nutr. 86:74-82.
Givens, D.I., A. Allison, B. Cottrill and J.S. Blake. 2004. Enhancing the selenium content of bovine milk through alteration of the form and concentration of selenium in the diet of the dairy cow. J. Sci. Food Agric. 84:811-817.
Goff, J.P. 2006. Major advances in our understanding of nutritional influences on bovine health. J. Dairy Sci. 89:1292-3011.
Grasso, P., R.W. Scholz, R.J. Erskine and R.J. Eberhart. 1990. Phagocytosis, bacterial activity and oxidative metabolism of mammary neutrophils from dairy cows fed selenium-adequate and selenium-deficient diets. Am. J. Vet. Res. 51:269-275.
Gromer, S., J.K. Eubel, B.L. Lee and J. Jacob. 2005. Human selenoproteins at a glance. Cell. Mol. Life Sci. 62:2414-2437.
Gunter, S.A., P.A. Beck and J.M. Phillips. 2003. Effects of supplementary selenium source on the performance and blood measurements in beef cows and their calves. J. Anim. Sci. 81:856-864.
Gyang, E.O., J.B. Stevens, W.G. Olsen, S.D. Tsitsamis and E.A. Usenik. 1984. Effects of selenium-vitamin E injection on the bovine polymorphonucleated leukocytes phagocytosis and killing of Staphylococcus aureus. Am. J. Vet. Res. 45:175-179.
Hatfield, D.L. and V.N. Gladyshev. 2002. How selenium has altered our understanding of the genetic code. Mol. Cell Biol. 22:3565-3576.
Hatfield, D.L., B.A. Carlson, X.M. Xu, H. Mix and V.N. Gladyshev. 2006. Selenocysteine incorporation machinery and the role of selenoproteins in development and health. Prog. Nucleic Acid Res. Mol. Biol. 81:97-142.
Heard, J.W., G.P. Walker, P.J. Royle, G.H. McIntosh and P.T. Doyle. 2004. Effects of short term supplementation with selenized yeast on milk production and composition of lactating cows. Aust. J. Dairy Technol. 59:199-203.
Hemingway, R.G. 1999. The influences of dietary selenium and vitamin E intakes on milk somatic counts and mastitis in cows. Vet. Res. Commun. 23:481-499.
Hemingway, R.G. 2003. The influences of dietary intakes and supplementation with selenium and vitamin E on reproduction, diseases and reproductive efficiency in cattle and sheep. Vet. Res. Commun. 27:159-174.
Hogan, J.S., K.L. Smith, W.P. Weiss, D.A. Todhunter and W.L. Shockey. 1990. Relationships among vitamin E, selenium, and bovine blood neutrophils. J. Dairy Sci. 73:2372-2378.
Hogan, J.S., W.P. Weiss and K.L. Smith. 1993. Role of vitamin E and selenium in host defense against mastitis. J. Dairy Sci. 76:2795-2803.
Holben, D.H. and A.M. Smith. 1999. The diverse role of selenium within selenoproteins: a review. J. Am. Dietetic Assoc. 99:836-843.
Ip, C., H.J. Thompson, Z. Zhu and H.E. Ganther. 2000. In vitro and in vivo studies of methylselenic acid: evidence that a monomethylated selenium metabolite is critical for cancer chemoprevention. Cancer Res. 2000:2882-2886.
Jiang, C., H. Hu, B. Malewicz, Z. Wang and J. Lu. 2004. Selenite-induced p53 Ser-15 phosphorylation and caspase-mediated apoptosis in LNCaP human prostate cancer cells. Mol. Cancer Ther. 3:877-884.
Jukola, E., J. Hakkarainen, H. Saloniemi and S. Sankari. 1996. Blood selenium, vitamin E, vitamin A, and ß-carotene concentrations and udder health, fertility treatments, and fertility. J. Dairy Sci. 79:838-845.
Juniper, D.T, R.H. Phipps, A.K. Jones and G. Bertin. 2006. Selenium supplementation of lactating dairy cows: effect of selenium concentration on blood, milk, urine and feces. J. Dairy Sci. 89:3544-3551.
Kehrli, M.E. Jr. and J.A. Harp. 2001. Immunity in the mammary gland. Vet. Clin. North Am. Food Anim. Pract. 17:495-516.
Knowles, S.O., N.D. Grace, K. Wurms and J. Lee. 1999. Significance of amount and form of dietary selenium on blood, milk and casein selenium concentrations in grazing cows. J. Dairy Sci. 82:429-437.
Köhrle, J., R. Brigelius-Flohé, A. Böck, R. Gärtner, O. Meyer and L. Flohé. 2000. Selenium in biology: facts and medical perspectives (a review). Biol. Chem. 381:849- 864.
Koller, L.D. and J.H. Exon. 1986. The two faces of selenium-deficiency and toxicityare similar in animals and man. Can. J. Vet. Res. 50:297-306.
Kommisrud, E., O. Østerås and T. Vatn. 2005. Blood selenium associated with health and fertility in Norwegian dairy herds. Acta. Vet. Scand. 46:229-240.
Krehl, W.A. 1970. Selenium - the maddening mineral. Nutr. Today 1970(Winter):26- 32.
Kryukov, G.V., S. Castellano, S.V. Novoselov, A.V. Lobanov, O. Zehtab, R. Guigo and V.N. Gladyshev. 2003. Characterization of mammalian selenoproteins. Science 300:1439-1440.
Larsen, H.J.S. 1993. Relations between selenium and immunity. Nor. J. Agric. Sci. 11(Suppl. 1):105-119.
Lindmark-Månsson, H. and B. Åkesson. 2001. Purification and immunochemical assay of bovine extracellular glutathione peroxidase. Int. Dairy. J. 11:649-655.
Lu, J. and C. Jiang. 2005. Selenium and cancer chemoprevention: hypothesis integrating the actions of selenoproteins and selenium metabolites in epithelial and non-epithelial target cells. Antioxid. Redox Signal. 7:1715-1727.
Mahan, D.C. 2000. Effect of organic and inorganic selenium sources and levels on sow colostrum and milk selenium content. J. Anim. Sci. 78:100-105.
Malbe, M., M. Klaassen, W. Fang, V. Myllys, M. Vikerpuur, K. Nyholm, S. Sankari, K. Suoranta and M. Sandholm. 1995. Comparison of selenite and selenium yeast feed supplements on Se-incorporation, mastitis and leukocyte function in Se-deficient dairy cows. J. Vet. Med. 42:111-121.
Malbe, M., E. Klaassen, L. Kaartinen, M. Attila and F. Atroshi. 2003. Effects of oral selenium supplementation on mastitis markers and pathogens in Estonian cows. Vet. Therapeutics 4:145-154.
Malbe, M., M. Attila and F. Atroshi. 2006. Possible involvement of selenium in Staphylococcus aureus inhibition in cow’s whey. J. Anim. Physiol. Anim. Nutr. 90:159- 164.
McKenzie, R.C., T.S. Rafferty and G.J. Beckett. 1998. Selenium: an essential element for immune function. Immunol. Today 19:342-345.
McKenzie, R.C., T.S. Rafferty, J.R. Arthur and G.J. Beckett. 2001. Effects of selenium on immunity and aging. In: Selenium: Its Molecular Biology and Role in Human Health (D.L. Hatfield, ed). Kluwer Academic Publishers, Boston, MA, USA, pp. 258-272.
Mehrzad, J., H. Dosogne, E. Meyer, R. Heyneman and C. Burnenich. 2001. Respiratory burst activity of blood and milk neutrophils in dairy cows during different stages of lactation. J. Dairy Res. 68: 399-415.
Ministry of Agriculture, Fisheries and Food. 2000. The Feedstuffs Regulations. Publication of her Majesty’s Stationery Office, London, UK.
Ndiweni, N. and J.M. Finch. 1995. Effects of in vitro supplementation of bovine mammary gland macrophages and peripheral blood lymphocytes with α-tocopherol and sodium selenite: implications for udder defences. Vet. Immunol. Immunopathol. 47:111-121.
Ndiweni, N. and J.M. Finch. 1996. Effects of in vitro supplementation with α-tocopherol and selenium on bovine neutrophil functions: implications for resistance to mastitis. Vet. Immunol. Immunopathol. 51:67-78.
Ortman, K. and B. Pehrson. 1999. Effect of selenate as a feed supplement to dairy cows in comparison to selenite and selenium yeast. J Anim. Sci. 77: 3365-3370.
Paape, M. J., D.B. Bannerman, X. Zhao and J.-W. Lee. 2003. The bovine neutrophil: structure and function in blood and milk. Vet. Res. 34:597-627.
Pehrson, B. 1993. Diseases and diffuse disorders related to selenium deficiency in ruminants. Nor. J. Agric. Sci. Suppl. 11:79-93.
Pehrson, B., K. Ortman, N. Madjid and U. Trafikowska. 1999. The influence of dietary selenium as selenium yeast or sodium selenite on the concentration of selenium in the milk of suckler cows and on the selenium status of their calves. J. Anim. Sci. 77:3371- 3376.
Ranjan, R., D. Swarup, R. Naresh and R.C. Partra. 2005. Enhance erythrocytic lipid peroxides and reduced plasma ascorbic acid, and alteration in blood trace elements level in dairy cow mastitis. Vet. Res. Commun. 29:27-34.
Rayman, M.P. 2000. The importance of selenium to human health. The Lancet 356:233- 241.
Rayman, M.P. 2004. The use of high-selenium yeast to raise selenium status: how does it measure up? Brit. J. Nutr. 92:557-573.
Rebsch, C.M., F.J. Penna III and P.R. Copeland. 2006. Selenoprotein expression is regulated at multiple levels in prostate cells. Cell Res. 16:940-948.
Rocher, C., J.-L. Lalanne and J. Chaudière. 1992. Purification and properties of a recombinant sulfur analog of purine selenium-glutathione peroxidase. Eur. J. Biochem. 205:955-960.
Romanowska, M., K.D. Kikawa, J.R. Fields, A. Maciag, S.L. North, Y.-H. Shiao, K.S. Kasprzak and L.M. Anderson. 2007. Effects of selenium supplementation on expression of glutathione peroxidase isoforms in cultured human lung adenocarcinoma cell lines. Lung Cancer 55:35-42.
Ropstad, E.G., G. Overnes and A. Refsdal. 1987. Selenium levels in Norwegian dairy herds related to reproduction and health performance. Acta Agric. Scand. 37:397- 405.
Rotruck, J.T., A.L. Pope, H.E. Ganther, A.B. Swanson, D.G. Hafeman and W.G. Hoekstra. 1973. Selenium: biochemical role as a component of glutathione peroxidase. Science 179:588-590.
Saijoh, K., N. Saito, M.J. Lee, M. Fujii, T. Kobayashi and K. Sumino. 1995. Molecular cloning of cDNA encoding a bovine selenoprotein P-like protein containing 12 selenocysteines and a (His-Pro) rich domain insertion, and its regional expression. Mol. Brain Res. 30:301-311.
Schwarz, K. and C.M. Foltz. 1957. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 79:3292-3293.
Segerson, E.C., G.J. Riviere, H.L. Dalton and M.D. Whitacre. 1981. Retained placenta of Holstein cows treated with selenium and vitamin E. J. Dairy Sci. 64:1833-1836.
Sivertsen, T., G. Øvernes, O. Østerås, U. Nymoen and T. Lunder. 2005. Plasma vitamin E and blood selenium concentrations in Norwegian dairy cows: regional differences and relations to feeding and health. Acta Vet. Scand. 46:177-191.
Smith, K.L., J.H. Harrison, D.D. Hancock, D.A. Todhunter and H.R. Conrad. 1984. Effect of vitamin E and selenium supplementation on incidence of clinical mastitis and duration of clinical symptoms. J. Dairy Sci. 67:1293-1300.
Smith, K.L., J.S. Hogan and H.R. Conrad. 1987. Vitamin E, selenium levels may be linked to mastitis in dairy cows. Feedstuffs 8(June):10.
Spallholz, J.E. 1990. Selenium and glutathione peroxidase: essential nutrient and antioxidant component of the immune system. Adv. Exp. Med. Biol. 262:145-158.
Spears, J.W. 2000. Micronutrients and immune function in cattle. Proc. Nutr. Society 59: 587-594.
Stadtman, T.C. 1996. Selenocysteine. Annu. Rev. Biochem. 65:83-100.
Stapleton, S.R. 2000. Introduction: the selenium conundrum. Cell Mol. Life Sci. 57:1823- 1824.
Suzuki, K.T. and Y. Ogra. 2002. Metabolic pathway for selenium in the body: speciation by HPLC-ICP MS with enriched selenium. Food Addit. Contam. 19:974-983.
Turner, R.J. and J.M. Finch. 1991. Selenium and the immune response. Proc. Nutri. Soc. 50:275-285.
Weiss, W.P. 2003. Selenium nutrition of dairy cows: comparing responses to organic and inorganic forms. In: Nutritional Biotechnology in the Feed and Food Industries. Proceedings of Alltech’s 19th Annual Symposium (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, Nottingham, UK, pp. 333-343.
Weiss, W.P. and J.S. Hogan. 2005. Effect of selenium source on selenium status, neutrophil function, and response to intramammary endotoxin challenge of dairy cows. J. Dairy Sci. 88:4366-4374.
Weiss, W.P., J.S. Hogan, K.L. Smith and K.H. Hoblet. 1990. Relationships among selenium, vitamin E, and mammary gland health in commercial dairy herds. J. Dairy Sci. 73:381-390
Wendel A., B. Kerner and K. Graupe. 1978. The selenium moiety of glutathione peroxidase. Hoppe-Seyler’s Z. Physiol. Chem. 359:1035-1036.
WHO (World Health Organization ).1987. Selenium. In: Environmental Health Criteria 58. A Report of the International Programme on Chemical Safety. WHO, Geneva, Switzerland.
Yang, G., Wang S., R. Zhou and S. Sun. 1983. Endemic selenium intoxication of humans in China. Am. J. Clin. Nutr. 37:872-881.
1 McGill University, Ste. Anne de Bellevue, Quebec, Canada
2 Ferdowsi University of Mashhad, Mashhad, Iran













United States