Effects of supplementary selenium source on performance, blood measurements, and immune function in beef cows and calves
Selenium (Se) deficiency in grazing and forage-fed cattle is widespread in the United States as well as in other countries (Kubota et al., 1967; Carter et al., 1968; van Vleet, 1980). As an integral part of the enzyme glutathione peroxidase (GSH-Px), selenium functions to prevent oxidative damage to body tissues (Hoekstra, 1974). Also, selenium deficiencies can inhibit the immunoglobulin G (IgG) response to in vivo challenges with sheep red blood cells (Mulhern et al., 1985) and the detoxification of certain xenobiotic compounds (Burk, 1983).
Studies have indicated that calves can be severely depleted of selenium and Se-dependent GSH-Px (Arthur, 1981; Siddons and Mills, 1981; Koller et al., 1984) but exhibit no clinical deficiency unless they are subjected to an oxidant or other type of stress. More recent studies have shown that selenium from sodium selenite is poorly transferred to milk (Ortman and Pehrson, 1999) and that it is unable to maintain the selenium status of nursing calves (Koller et al., 1984; Ortman and Pehrson, 1999). Compared to non-ruminants, ruminants absorb sodium selenite poorly (Wright and Bell, 1966), but recent research with organic selenium from seleno-yeast, has shown that cows supplemented with seleno-yeast are more effective at transferring selenium to calves via placental transfer and milk than cows supplemented with sodium selenite (Pehrson et al., 1989; Ortman and Pehrson, 1999; Pehrson et al., 1999). Studies with beef cattle in areas that are considered marginal or deficient in selenium are limited (Hidiroglou and Jenkins, 1975; Spears et al., 1986; Ortman and Pehrson, 1999).
In light of these facts, we designed two experiments. The first was to examine selenium source and supplementation level with gestating and lactating beef cows on calf growth, reproduction, and blood parameters when fed grass hay and pastured on forages that were marginally deficient in selenium. The second experiment was designed to examine the effects of selenium source and supplementation level of nursing cows on whole blood selenium (WBSe), GSH-Px and immune function of calves at weaning.
Materials and Methods
EXPERIMENT 1
On 1 October 1999, at the Southwest Research & Extension Center 120 pregnant crossbred beef cows (average BW = 528 ± 10.2 kg) of mostly British (Angus and Hereford) and Continental (Simmental) breeding and bred to begin calving on February 1 were fed bermudagrass (Cynodon dactylon [L.] Pers.)/dallisgrass (Paspalum dilatatum Poir.) hay with ad libitum access to salt blocks (NaCL; Morton Salt Co., Salt Lake City, UT) as their only supplemental mineral source for two months. This preliminary feeding period was an attempt to adjust the selenium status of the cows to the selenium level expected for cows maintained in the local environment without selenium supplementation.
On 2 December 1999, cows were weighed and body condition scored (Wagner et al., 1988). Cows were also injected with a seven-way Clostridial antigen (Vision 7 Somnus; Bayer Corp., Shawnee Mission, KS) to increase Clostridial antibodies in the colostrum (Clarkson et al., 1985) and dewormed with fenbendazole (Safeguard; Intervet, Inc., Millsboro, DE). Cows were sorted into six groups of 20 stratified by BCS, BW, breed, and age, and assigned randomly to one of six 5.1-ha common bermudagrass pastures for two years. Beginning on December 2, cows had ad libitum access to a bermudagrass/dallisgrass hay (Table 1) plus they were allowed limited access to a 2.4-ha winter annual paddock planted in half of each pasture for a protein and energy supplement (Gunter et al., 2002). Treatments were randomly assigned to two pastures; cows had ad libitum access to one of the following three free choice mineral supplements which contained (Table 1): 1) no supplemental Se, 2) 26 mg of supplemental Se/kg of free-choice mineral from sodium selenite (Selenium Premix; Southeastern Minerals, Inc., Bainbridge, GA), and 3) 26 mg of supplemental Se/kg of a free choice mineral from seleno-yeast (Sel-PlexTM; Alltech Inc., Nicholasville, KY). A commercial feedmill (Sunbelt Custom Minerals, Inc., Sulphur Springs, TX) manufactured and delivered the mineral supplements in November of each year. Free choice minerals were formulated for consumption at a rate of 113 g/cow daily; minerals were re-supplied weekly by putting 1,000 g/cow in a weather vanestyle mineral feeder. On alternate weeks, mineral feeders were cleaned and orts were weighed to determine mineral intake. After orts were weighed, if they appeared contaminated with feces or water, they were subsampled for DM determination and the remainder was discarded.
Each fall, winter annual grasses were sod-seeded into a portion of the 5.1-ha common bermudagrass pastures during the first week of October. Before planting, standing herbage mass was removed from the area by continuously stocking with cattle until the standing herbage mass was visually estimated to be <5 cm in height. Using a Marliss no-till drill (Sukup Manufacturing, Jonesboro, AR), the dormant pastures were seeded with 134 kg of a 1:1 mixture (wt:wt) of ‘Koolgrazer’ rye (James Reneau Seed Co., Shamrock, TX) and ‘Coker 9542’ soft-red wheat (Southeast Research & Extension Center-Rohwer Division, Rohwer, AR)/kg via the small grain box, and 22 kg of ‘Marshall’ annual ryegrass (Wax Seed Co., LLC, Amory, MS)/kg via the grass seed box. Diammonium phosphate (18-46-0) was banded with the seed at a rate of 168 kg/ha via the fertilizer box. Phosphorus rate was selected to fulfill soil test recommendations (Chapman, 1998). In late January and mid-March, sod-seeded portions of pastures were fertilized with an additional 55 kg of N/ha using ammonium nitrate (34-0-0). The entire pastures were fertilized with 56 kg/ha of N, P, and K in late May from a blended fertilizer, and with 55 kg of N/ha using ammonium nitrate in late June and early August.
Cows were restricted from winter annual paddocks on non-grazing days using electric fences.
Cows were allowed to graze winter annual paddocks 1 day/wk (7 hrs/d) in December, 2 days/ wk in January, 3 days/wk from February 1 until March 15, and 4 days/wk from March 15 until May 1. On 1 May, electric fences that divided pastures were removed and cows were allowed continuous access to the entire pasture until weaning. Winter annual paddocks were planted in October, and the forage was stockpiled until grazing began in December. Therefore, forage was not limiting for grazing in January and February when plant DM production was less than cattle demand. Hay was offered in the form of round bales in ring-type hay feeders and technicians maintained records of the quantity of hay fed in each pasture. To calculate estimated daily hay DMI/cow, quantities of hay offered in each pasture were corrected for wastage based on feeder type and the data of Buskirk et al. (2000) then divided by the number of cows in each pasture.
Chemical composition of the hay (core samples from 10% of the bales fed) and winter annual pasture forage (hand plucked in January, March, and April, composited across pastures and times) was determined at a commercial laboratory; DM and CP were determined as described by AOAC (1990), and TDN was determined as described by Weiss et al. (1992). Hay or pasture samples were composited across days and pastures, and analyzed for Ca, P, Mg, K, Na, Fe, Zn, Cu, Mn, and Mo by inductively coupled plasma emission spectroscopy (Thermo Jerrell Ash Corp.; Franklin, MA) as described by Sirois et al. (1991); selenium concentration was conducted at Michigan State University, Animal Health Diagnostic Laboratory by inductively coupled argon plasma emission spectroscopy using the improved fluorometric method (Olsen et al., 1975). On April 25 in both years, an Angus bull that had passed a breeding soundness examination was placed with each group of cows for a 60-day breeding season. Bulls were rotated weekly among cow groups to minimize the effect of individual bulls.
Cows and calves were weighed and body condition scored on February 8 (early February), May 24 (late May), June 29 (late June), and September 27 (late September) in 1999 and February 8 (early February), May 24 (late May), June 28 (late June), and October 16 (late September) in 2000. The morning after calving, calves were weighed, tattooed in both ears with an individual number, and male calves were surgically castrated. Calves were also evaluated for calving ease, agility, and vigor scores. Calving ease scores were as follows: 1 = unassisted birth, 2 = minor assistance, 3 = mechanical assistance, 4 = caesarian section, and 5 = abnormal presentation (Vandervelde et al., 1990). Agility scores were assigned as follows: 1 = moves well and correct posture, 2 = moves showing slight stiffness in legs, 3 = significant stiffness in gait, 4 = significant stiffness in gait and slight arch in the topline, and 5 = significant stiffness in gait and arch in topline. Vigor scores were assigned as follows: 1 = alert and active, 2 = alert, 3 = appears healthy, but somewhat listless, 4 = listless, and 5 = listless and unresponsive. On May 24 in both years, cows were treated for internal and external parasites (Ivomec; Merial Inc., Duluth, GA), vaccinated with a seven-way Clostridial antigen (Vision 7 Somnus), and vaccinated for infectious bovine rhinotracheitis, bovine viral diarrhea, parainfluenza-3, bovine respiratory syncytial virus, plus five strains of Leptospirosis (Triangle 4 + PH-K and TriVib L5; Fort Dodge Animal Health, Overland Park, KS).
Table 1. Composition of free-choice minerals, hay, and pasture forage offered to beef cows (Exp. 1)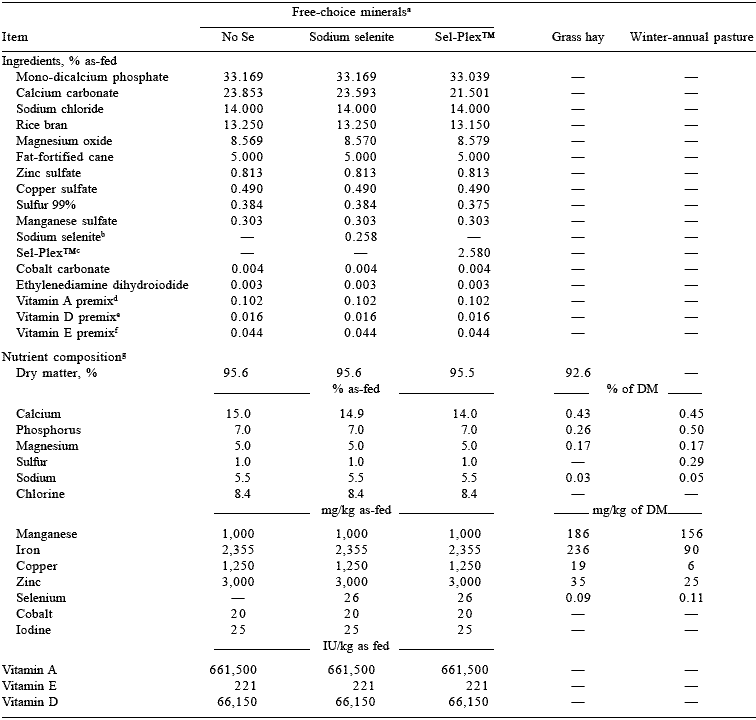
To enlarge the image, click here
aManufactured by Sunbelt Custom Minerals, Inc., Sulphur Springs, TX.
bSelenium Premix, Southeastern Minerals, Inc., Bainbridge, GA; contained 1.0% Se and 36.0% Ca.
c35.0% CP, 6.0% crude fiber, 0.3% crude fat, and 1.0% Se.
dROVIMIX A-650; 650,000 IU/g, Roche Vitamins, Inc., Parsippany, NJ.
eROVIMIX D3-400; 400,000 IU/g, Roche Vitamins, Inc.
fROVIMIX E-50 Absorbate; 50,000 IU/g, Roche Vitamins, Inc.
gValues for mineral supplements are calculated from NRC (1996) and values for grass hay and pasture forage are analyzed values.
Blood samples were collected from four randomly selected cows and their calves within each pasture via jugular venipuncture into a 10 ml tube containing EDTA for determination of WBSe and GSH-Px in the erythrocytes. Cows were bled on December 2 (trial initiation), February 8 (beginning of calving season), and April 19 (beginning of the breeding season) in 1999, and December 7, February 8, and April 19 in 2000; calves were bled at 0730 after birth and on 24 May (near peak lactation) in both years. At bleeding, blood samples were immediately placed on ice, then frozen (-20° C) until overnight shipment on dry ice to the laboratory for analysis.
Blood samples were prepared for analysis and WBSe was determined as described by Reamer and Veillon (1983). Glutathione peroxidase activity per gram of hemoglobin (Hb) in the erythrocytes was determined using the procedure of Paglia and Valentine (1967) as modified by Lawrence et al. (1974). Cows were checked for pregnancy by rectal palpation on 27 September 1999 and 16 October 2000 by a veterinarian (Powell and Perry Veterinary Clinic, Hope, AR). In 1999, cows deemed nonpregnant were replaced by mature pregnant cows (multiparous) in order to maintain 20 cows in each pasture. Postpartum interval was calculated by subtracting 283 days from the calving date of the following year to estimate conception date.
Body weight of cows, cow body condition score, conception rate, postpartum interval, and hay DMI were analyzed using PROC GLM (SAS Inst. Inc.; Cary, NC) as a split plot design with the effects of treatment (main plot), year (sub plot), treatment x year, and the covariates, cow age and calving date, in the model (Steel and Torrie, 1980). Pastures were considered the experimental units, so the treatment effects were tested with pasture within treatment, and year and year x treatment were tested with pasture within year x treatment as the error terms.
Calf BW and calf total gain data were analyzed using PROC GLM (SAS Inst. Inc.) as a split plot design with the effect of treatment (main plot), year (sub plot), and treatment x year and the covariates, cow age, calf gender, birth date, and birth weight in the model (Steel and Torrie, 1980). Models used to analyze birth date (Julian date) and birth weight were analyzed similarly to the preceding model described, except calving date or birth weight, respectively, were excluded as a covariate. Because the experiment required a mixed model for analysis, year was considered a random effect and treatment was considered a fixed effect; all parameters discussed were averaged across years by treatment (Steel and Torrie, 1980). If a cow died during the course of the study, the affected cow and calf were immediately replaced with a spare cow-calf pair to equalize stocking rate; however, the data from replacement cows and calves were not used in the statistical analysis. Least square means were separated using the following contrasts: 1) no supplemental Se vs supplemental Se, and 2) sodium selenite vs Sel-PlexTM with pasture within treatment as the error term (Steel and Torrie, 1980).
EXPERIMENT 2
On 7 November 2001 at the Southwest Research & Extension Center, 18 spring-born Angus sired steers (average BW ± SE = 200 ± 5.2 kg) were selected from the six groups of cow-calf pairs (n = 3 calves/cow group) in Experiment 1. At weaning on October 16, the steers were treated for internal and external parasites (Ivomec; Merial Inc., Duluth, GA), vaccinated with a seven-way Clostridial antigen (Vision 7 plus Somnus; Bayer Corp.
Shawnee Mission, KS) and with a modified live infectious bovine rinotracheitis, bovine viral diarrhea, parainfluenza-3, bovine respiratory syncytial virus vaccine (Cattle Master 4; Pfizer Inc., New York, NY). After weaning, the calves were fed freechoice bermudagrass hay with 1.40 kg/calf daily of a supplement supplying 350 mg chlortetracycline/ day (Aureomycin-90; Alpharma Inc., Fort Lee, NJ) through the end of the trial on November 27. The supplement contained (as-fed basis) 29.8% defatted rice bran, 49.7% ground corn, 9.9% cottonseed hulls, 5% liquid protein supplement, 2.3% ammonium sulfate and 3.3% of the Se-free mineral as a mineral premix. The weaning supplement was calculated to supply 15% CP, 1.74 Mcal NEm/kg, 1.12 Mcal NEg/kg and 0.24 ppm Se (DM basis).
On November 7, (22 days post-weaning) the steers were weighed (unshrunk) beginning at 0730 and blood was collected via jugular venipuncture. Blood was immediately placed on ice, then frozen (-20°C) and sent for analysis of WBSe and GSHPx as described in Experiment 1. Additional blood was collected and placed on ice for transport to University of Arkansas Animal Science Laboratory (374 km, 4 hrs transit time) for analysis of lymphocyte proliferation, macrophage phagocytosis, and interferon-γ (IFN) production.
Blastogenic response of peripheral blood lymphocytes was determined by methods adapted from Blecha et al. (1983). Briefly, peripheral blood mononuclear cells were isolated by density gradient centrifugation using Ficoll gradient (Histopaque 1077, density=1.077g/ml; Sigma Chemical Co., St. Louis, MO). Adding 1 ml of sterile water to the isolated cell pellet for 20 sec lysed any remaining erythrocytes. Cells were re-suspended in RPMI 1640 (Sigma Chemical Co.) at 2 x 106 cells/ml. Cells were plated in triplicate in 96-well round bottom plates in 100 ml aliquots. Cells were then incubated in unstimulated cultures and in cultures stimulated with 25 µg/ml concanavalin A (Con A; Sigma Chemical Co.), 40 µg/ml phytohemagglutinin (PHA; Sigma Chemical), and 15 µg/ml pokeweed mitogen (PWM; Sigma Chemical Co.). Incubation, labeling with tritiated-thymidine, and cell harvesting followed procedures outlined by van Heugten et al. (1994).
To summarize, cells were incubated for 48 hrs at 39°C and 5% CO2, tritiated-thymidine was added to each well, and cultures were incubated for an additional 18 hrs. Cells were harvested on glass fiber mats and the radioactivity was measured as counts/ minute on a liquid scintillation analyzer (TRI-CARB 2200CA, Packard Instrument Co., Downers Grove, IL). Production of IFN by activated immune cells was quantified by incubating whole blood with 5 mg/ml PHA for 24 hrs at 39°C and 5% CO2 followed by harvesting serum by centrifugation. Interferon-γ concentration was then determined by ELISA (Biocor Animal Health, Inc., Omaha, NE).
Monocytes/macrophages were isolated and phagocytic ability was determined by methods adapted from Moneleone et al. (1999) and Nibbering et al. (1987). In short, peripheral blood mononuclear cells were isolated as described above for the lymphocyte blastogenesis assay. Two ml of cells suspended in LM Hahn medium (2 x 106 cells/ ml) were incubated in duplicate in 6-well plates containing a glass cover slip for 16 hrs at 39°C.
Isolation of monocytes/macrophages occurs because of the propensity of the cells to adhere to the glass surface of the coverslip. Following the 16 hr incubation, excess medium was removed, and two ml of a 5% porcine red blood cell (PRBC) suspension was added to the monocytes/ macrophages remaining on the glass coverslips.
Cells were incubated with PRBC at 39ºC and 5% CO2 for 2 hrs, after which coverslips were removed and rinsed with warmed LM Hahn medium to remove non-adherent cells and non-phagocytized PRBC. Cells were then fixed on coverslips and stained (Heme-3®, Fisher Scientific, Pittsburgh, PA) for visualization under a microscope at 1000x. The percentage of phagocytic macrophages and number of PRBC consumed by each phagocytic macrophage were determined by visual assessment of 200 macrophages/monocytes on each duplicate coverslip. On November 13, in vivo cell-mediated immunity was measured by determining the ability of the steers to respond to an intradermal injection of 0.2 mL PHA. The injection site was clipped and skin fold thickness was measured before and 8, 24, 32, and 48 hrs after injection using micrometric calipers (Fisher Scientific). Differences in pre- and post-injection skin fold thickness at the injection site are reported.
Whole blood selenium concentration, GSH-Px, lymphocyte proliferation, macrophage phagocytosis, and IFN concentrations were analyzed by ANOVA as a completely random experimental design using the PROC GLM (SAS Inst. Inc.). Residual error was used as the error term for testing the effect of treatment. Least square means were separated using predicted differences when there was a significant F-test (P<0.05; Steel and Torrie, 1980).
Skin-swelling response to PHA injection over time was analyzed as a split-plot in time. Treatment was included in the main plot and time post-injection was included in the sub plot. The effect of treatment, time post injection, and the treatment by time interaction was tested using residual error.
Results and discussion
EXPERIMENT 1
Bi-weekly intake estimates of the free-choice minerals did not differ (P>0.10) on any date between cows supplemented with selenium and cows receiving no selenium (Figure 1); however, on weeks 1 and 18 mineral intake by cows supplemented with Sel-PlexTM was greater (P<0.05) than for cows supplemented with sodium selenite. Each fall when we began to measure mineral intake, it required approximately 6 weeks for cows to become accustomed to the feeder and achieve the target intake of 113 g/day. Also, from week 6 to 28 (January to July), intake seemed to vary more by week than during weeks 30 to 42 (July to October), which probably resulted from inclement weather as a result of frequent thundershowers that normally occur in the spring. The average daily intake per cow for no selenium, sodium selenite, and Sel-PlexTM minerals was 108, 107, and 109 g, respectively.
The bermudagrass/dallisgrass hay fed during the winter averaged 11% CP and 56% TDN (DM basis) over the two year period. Based on the average measured hay DMI of 8.2 kg/day (Table 2) and a daily milk production of 10 kg, supplemental CP and TDN requirements were 101 g/day and 2.7 kg/day, respectively, at peak lactation (NRC, 1996).
Samples of the winter annual pasture collected over the two years averaged 25% CP and 63% TDN. It was estimated that grazing winter annual paddock 3 days/week would meet or exceed the requirements of the cow for supplemental protein and energy based on the research of Gunter et al. (2002). Calcium, P, Mg, S, Mn, and Fe amounts supplied by both hay and winter annual pasture samples exceeded the NRC (1996) requirements.
Regarding the high sulfur concentration in the winter annual pasture, the NRC (1996) points out that sulfur in the diet can become toxic over 0.30% of dietary DM; however, some researchers have reported sulfur toxicity with concentrations as low as 0.22% of dietary DM (Gould et al., 1991). Hay and winter annual pasture samples were deficient in sodium relative to the NRC (1996) requirement of 0.08% of dietary DM. Copper and zinc concentrations in hay samples exceeded NRC (1996) requirements; however, the copper and zinc concentrations in the winter annual pasture were deficient. Selenium concentrations in hay and winter annual pasture samples were marginally deficient as suggested by Puls (1989; 0.10 to 0.25 ppm). However, the NRC (1996) suggests 0.10 ppm in dietary DM meets animal requirements.
Body weight did not differ on any date between cows supplemented with selenium and cows receiving no selenium (Table 2) or between cows supplemented with sodium selenite or Sel-PlexTM.
Other research has reported cows consuming diets that were moderately deficient in selenium (0.05 to 0.09 ppm) and were injected monthly with sodium selenite and vitamin E lost less BW during the winter than non-injected cows. However, the injected cows gained less BW during the summer than non-injected cows (Spears et al., 1986).
Figure 1. Intake (measured every 2 wk) by beef cows of a free-choice mineral differing in selenium content and source. No Se = no supplemental Se in free-choice minerals, sodium selenite = free-choice minerals with 26 mg of Se/kg from sodium selenite, and Sel-Plex™ = free-choice minerals with 26 mg of Se/kg from selenium yeast (* signifies a significant (P<0.05) contrast between mineral supplements with sodium selenite vs Sel-PlexTM).
Table 2. Body weight, body condition score, conception rate, and post-partum interval of beef cows offered no Se, sodium selenite, or Sel-Plex™ in free-choice minerals (Exp. 1).
aContrasts: no Se vs Se = no Se vs sodium selenite and Sel-Plex™; NaSe vs SP = sodium selenite vs Sel-Plex™.
bBody condition score range 1 to 9; 1 = emaciated, 9 = obese (Wagner et al., 1988).
Research in Alaska showed that cows administered an intra-ruminal selenium bolus (sodium selenite) did not differ in BW from cows not given a Se bolus (Bruce, 1997). Body condition score did not differ on any date between cows supplemented with selenium and cows receiving no selenium (Table 2) or between cows supplemented with sodium selenite and Sel-PlexTM. Research has also reported cows supplemented with sodium selenite did not differ in fat thickness compared to non-supplemented cows (Bruce, 1997).
Conception rates and postpartum intervals did not differ between cows supplemented with selenium and cows receiving no selenium (Table 2), or between cows supplemented with sodium selenite and Sel-PlexTM. Research has reported cows consuming diets that were moderately deficient in selenium that were injected with sodium selenite and vitamin E did not differ in conception rate, but they did report a decrease in the postpartum interval compared to cows not injected with selenium and vitamin E (Spears et al., 1986). However, Aréchiga et al. (1994) reported that when Holstein dairy cows were injected with a selenium and vitamin E mixture they had increased conception rates and a decreased postpartum interval. Hay DMI did not differ between cows supplemented with selenium and cows receiving no selenium (Table 2), or between cows supplemented with sodium selenite or Sel- PlexTM. In previous research at our location with cows limit-grazed on winter annual pasture 3 days/ week, hay DMI was similar to the values in the present study (Gunter et al., 2002).
Birth date, birth weight, BW of calves, total BW gain, and ADG did not differ between calves nursing cows supplemented with selenium and calves nursing cows supplemented with no selenium, or between calves nursing cows supplemented with sodium selenite or Sel-PlexTM (Table 3).
Researchers have reported significant increases in ADG and/or total BW gain of calves with injectable sodium selenite and vitamin E (Hidiroglou and Jenkins, 1975; Spears et al., 1986). However, research conducted at this research center showed that when nursing calves were supplemented with selenium from sodium selenite via constant release bolus (Dura-Se, Schering-Plough Animal Health; Union, NJ), their ADG was similar to non-supplemented controls (Phillips et al., 1989). No differences (P>0.16) were noted in mortality among treatments in the present study (data not shown).
Whole blood Se concentrations in cows at the beginning of the experiment in December did not differ between cows supplemented with no selenium and cows supplemented with selenium (Table 4). Initial WBSe was also unaffected by selenium sources. The WBSe values reported in December are suggested to be marginal (60 to 150 ng Se/ml) for optimal GSH-Px and immune function (Puls, 1989). At the beginning of the calving season (early February), cows fed Se-fortified minerals for approximately 64 days had higher (P=0.003) WBSe than cows fed minerals with no selenium. At this point in time (early February), the WBSe of the Se-supplemented cows (average = 158 ng/ml) had been increased to a point (160 to 1,200 ng/ml), which is considered to be adequate for optimal GSH-Px and immune function (Puls, 1989). In early February, cows supplemented with sodium selenite had lower (P=0.01) WBSe than cows supplemented with Sel- PlexTM. Whole blood selenium concentrations in cows supplemented with Sel-PlexTM were 23% greater than cows supplemented with sodium selenite, which suggests that Sel-PlexTM was more available. Research by Pehrson et al. (1989) showed that Sel-PlexTM was 1.8 times more available than sodium selenite in heifers. At the beginning of the breeding season in April, cows fed Se-fortified minerals had higher (P=0.003) WBSe than cows fed minerals with no selenium and cows supplemented with Sel-PlexTM had higher (P=0.03) WBSe than cows supplemented with sodium selenite. As noted in early February, the difference between WBSe in cows supplemented with Sel- PlexTM and sodium selenite indicates that Sel-PlexTM is more available.
Table 3. Birth date and weight, body weight, total body weight gain, and ADG of calves nursing beef cows offered no Se, sodium selenite, or Sel-Plex™ in free-choice minerals (Exp. 1).
aContrasts: no Se vs Se = no Se vs sodium selenite and Sel-Plex™; NaSe vs SP = sodium selenite vs Sel-Plex™.
bBirth weight used as a covariate.
cBirth date used as a covariate.
dAgility score: 1.0 = moves well and correct posture, 2.0 = moves showing slight stiffness in legs, and 3.0 = significant stiffness in gait.
eCalving ease: 1.0 = unassisted birth (Vandervelde et al., 1990).
fVigor score: 1 = alert and active, 2 = alert, and 3 = appears healthy, but somewhat listless.
Whole blood selenium concentrations at birth in calves from cows fed Se-fortified minerals were higher (P=0.01) than in calves from cows fed minerals with no selenium (Table 4). Calves from cows supplemented with Sel-PlexTM had higher (P=0.02) WBSe at birth than calves from cows supplemented with sodium selenite. Koller et al. (1984) established that fetuses sequester Se from the dam and therefore normally have equal or greater WBSe than the dam. However, it is noteworthy that calves from cows fed Sel-PlexTM had greater WBSe at birth than calves from cows fed no selenium or sodium selenite. Pehrson et al.
(1989) also reported milk selenium concentration was increased and WBSe and GSH-Px of suckling calves was increased by feeding organic selenium to the dam. Ortman and Pehrson (1999) reported that the selenium concentrations in milk were increased by 190% in cows fed Sel-PlexTM compared to cows fed inorganic selenium. Therefore, increasing concentrations of selenium in milk by supplementing cows with Sel-PlexTM should improve the selenium status of suckling calves. Koller et al. (1984) reported that cows supplemented with sodium selenite gave birth to calves with elevated WBSe compared to non-supplemented cows. These researchers reported that the WBSe in calves nursing cows supplemented with sodium selenite decreased as they aged, and it was concluded that selenium from sodium selenite was inadequately transferred to calves via the milk.
Table 4. Whole blood Se concentration and glutathione peroxidase activity in beef cows offered no Se, sodium selenite, or Sel- PlexTM in free-choice minerals (Exp 1).
aContrasts: no Se vs Se = no Se vs sodium selenite and Sel-PlexTM; NaSe vs SP = sodium selenite vs Sel-PlexTM.
bBeginning of mineral feeding.
cStart of calving season.
dInitiation of the breeding season.
eEU/g Hb = nanomoles of NADPH oxidized per minute per gram of hemoglobin.
Glutathione peroxidase activity in erythrocytes from cows at the beginning of the experiment in December did not differ between cows supplemented with no selenium and cows supplemented with selenium (Table 4). Also, GSH-Px did not differ between selenium sources.
In early February and April, cows fed Se-fortified minerals for approximately 64 days had higher (P=0.05) GSH-Px than cows fed minerals with no selenium; however, the GSH-Px in cows supplemented with sodium selenite did not differ from cows supplemented with Sel-PlexTM. Glutathione peroxidase activity in erythrocytes from calves at birth from cows fed Se-fortified minerals was higher (P=0.11) than in calves from cows fed minerals with no selenium (Table 4); however, calves from cows supplemented with Sel- PlexTM had higher (P=0.05) GSH-Px at birth than calves from cows supplemented with sodium selenite. In late May, GSH-Px in calves nursing cows fed Se-fortified minerals did not differ from calves nursing cows fed minerals with no selenium; however, calves from cows supplemented with Sel- PlexTM had higher (P=0.10) GSH-Px than calves nursing cows supplemented with sodium selenite. Glutathione peroxidase activity is expected to be between 45 and 85 nmol of NADPH oxidized per minute (EU)/g Hb for cattle with adequate selenium status, and a critically low activity is considered to be <15 EU/g Hb (Erksine, 1993). Glutathione peroxidase activity was within the expected range for all cows and calves at all times during the study.
Research comparing selenium sources (Pehrson et al., 1999) demonstrated that Se from Sel-PlexTM is secreted via the milk in lactating beef cows, and cows supplemented with Sel-PlexTM were better able to maintain the GSH-Px in their nursing calves than cows supplemented with sodium selenite.
Selenium supplementation of gestating beef cows via fortified free-choice minerals benefited cows and calves by increasing WBSe and GSH-Px compared to no selenium supplementation. Blood variables from cows supplemented with no selenium indicated a risk of clinical selenium deficiency. The use of Sel-PlexTM as a selenium supplement in free-choice minerals compared to sodium selenite increased the WBSe of both cows and their calves and increased GSH-Px. Also, Sel-PlexTM supplementation of nursing calves maintained WBSe concentration and GSH-Px, which sodium selenite failed to accomplish. No improvement was noted in performance of cows and calves.
EXPERIMENT 2
Whole blood selenium concentrations were increased (P<0.01) by selenium supplementation (Table 5). Supplementation with Sel-PlexTM increased (P<0.01) WBSe compared to calves that had access to mineral fortified with sodium selenite.
Calves from the non-supplemented control treatment had low WBSe (54.7 ng/ml) and GSHPx (40.2 EU/g Hb) indicating a selenium deficiency (Puls, 1989; Erksine, 1993). It is unknown if the improvement in selenium status and immune function observed in these Se-supplemented steers directly resulted from maternal selenium transfer in the milk or through mineral intake by the calves that might have occurred before weaning. By the time of weaning, these calves would be expected to consume some of the mineral, but mineral intake was not differentiated between cows and calves in Experiment 1. Supplementation with Sel-PlexTM increased WBSe by 3.1 times compared to no selenium supplementation and 1.6 times compared to calves supplemented with sodium selenite. This better bioavailability with Sel-PlexTM compared to sodium selenite agrees with blood analysis with calves reported in Experiment 1. Similar results have been reported in cows (Pehrson et al., 1999; Koller et al., 1984), suckling calves (Pehrson et al., 1999), and growing calves (Nicholson et al., 1999). Koller et al. (1984) supplemented pregnant cows with varying concentrations of sodium selenite and reported selenium was transferred across the placenta to the fetus and that there were high selenium concentrations in the colostrum, but deficient selenium concentrations in the milk of cows 8 weeks post-partum. Ortman and Pehrson (1999) reported that milk selenium concentrations were increased by 190% in cows fed Sel-PlexTM compared to cows fed inorganic selenium sources.
The GSH-Px was also increased (P<0.01) by supplementation with selenium compared to no selenium supplementation, but there was only a numerical increase (P=0.38) in GSH-Px in calves from the Sel-PlexTM treatment compared to sodium selenite (Table 5). Glutathione peroxidase activity has been reported to increase in calves with selenium supplementation of their dams. In Experiment 1, it was noted that GSH-Px of calves was increased with selenium supplementation through 3 months of age; the results of the current trial showed that this trend continued through to weaning. Sel-PlexTM increased (P=0.10) GSH-Px compared to sodium selenite in calves up to 3 months of age (Table 5). There are data showing increased GSH-Px early in the life of calves reared by Se-supplemented cows (Pehrson et al., 1999; Koller et al., 1984) and in growing calves (Nicholson et al., 1999), but very little research showing the effect of longer-term supplementation of Sel-PlexTM to older calves. With long term supplementation, there may be adequate availability of sodium selenite to facilitate GSH-Px, while other selenium storage pools (muscle, liver and blood) can still accumulate additional selenium. This supports the observations of increased WBSe in calves from the Sel-PlexTM treatment at weaning, while GSH-Px was not significantly enhanced.
No differences were found in lymphocyte proliferation with either unstimulated cultures or cultures stimulated with Con A, PWM, or PHA (Table 5). Selenium supplementation has had variable effects on lymphocyte proliferation in ruminants. Cows supplemented with selenium had increased lymphocyte proliferation in cultures stimulated with Con A compared to cows that were deficient in selenium (Cao et al., 1992). Pollock et al. (1994) reported that lymphocyte proliferation in response to keyhole limpet haemocyanin stimulation was increased in calves supplemented with selenium compared to no selenium supplementation and calves supplemented with selenium and vitamin E; however, no treatment effects were observed in cultures stimulated with Con A or PWM. Turner et al. (1985) reported no difference in lymphocyte proliferation in response to PHA or PWM stimulation of sera from selenium-supplemented and non-supplemented lambs. In vitro selenium repletion of lymphocyte cultures has been shown to increase proliferation responses of Se-adequate dairy cows (Stabel et al., 1990) and lambs that were both Seadequate and deficient (Turner et al., 1985). This indicates an essential role of selenium on lymphocyte proliferation, which may be difficult to statistically separate with the current immunological techniques. Because the mineral mixture contained 221 IU/kg vitamin E and the predominantly forage diet would naturally contain high levels of tocopherols and tocotrienols (McDowell, 1997), the cattle in the current study could also be considered to have adequate dietary vitamin E. Vitamin E has a sparing effect on selenium (McDowell, 1997), which may mask some of the immunological effects of a marginal selenium deficiency.
Table 5. Effect of supplemental selenium level and source on whole blood selenium, GSH-Px, lymphocyte proliferation, macrophage age phagocytosis, and interferon-γ concentrations of weaned beef calves (Exp. 2).
aEU/g Hb = nanomoles of NADPH oxidized per minute per gram of hemoglobin.
bUnstimulated cultures received no mitogen to elicit proliferation response.
cLymphocytes cultured in the presence of concanavalin A to stimulate proliferation of thymic derived T lymphocytes associated with cellmediated immunity.
dLymphocytes cultured in the presence of phytohemagglutinin to stimulate proliferation of thymic derived T lymphocytes associated with cell-mediated immunity.
eLymphocytes cultured in the presence of pokeweed mitogen to stimulate proliferation of B lymphocytes associated with humoral immunity.
fThe percentage of macrophages that phagocytized at least one porcine red blood cell.
gPorcine red blood cells consumed per macrophage.
xyzLS means within rows with differing superscripts differ (P<0.01).
The average number of PRBC consumed by each macrophage was unaffected (P>0.30) by treatment (Table 5), although there was an increase (P<0.01) in the percentage of phagocytic macrophages with the Sel-PlexTM compared to non-supplemented or sodium selenite-supplemented calves. Combined vitamin E and selenium deficiency has been found to impair phagocytosis of sheep red blood cells by chicken macrophages (Dietert et al., 1990). Stabel et al. (1990) reported that blood macrophages from dairy cows have been shown to benefit from vitamin E supplementation with increases in interleukin-1 and expression of class II major histocompatability complex. Boyne and Arthur (1979) found neutrophil phagocytic ability was not changed with sodium selenite supplementation of semi-synthetic diets fed to Holstein steers, while the ability to kill phagocytized bacteria was increased 3-fold. The results of the current trial indicate that membrane-dependent functions of macrophages are affected by supplementation with Sel-PlexTM.
Macrophage function is essential for the initiation of the immune response to pathogenic invasion. The dramatic improvement in the percentage of phagocytic macrophages may have an impact on disease resistance of weaned calves.
Production of IFN by stimulated cultures was not significantly different among dietary treatments. Cytokines such as interleukin-12 and interleukin-6 are released by macrophages to activate T-cells, which then release IFN to further activate macrophages. Because of the lack of impact that selenium treatment had on IFN production by activated lymphocytes in vitro, it does not appear that the change in macrophage function with Sel- PlexTM supplementation is because of changes in activation of macrophages by T-cells.
Swelling reaction to intradermal PHA injections was affected (P<0.01) by time after injection (Figure 2). Selenium supplementation also tended to increase in vivo cell mediated immune response (P=0.12).
There has been little research conducted to determine the effect of selenium supplementation on in vivo cell-mediated immunity, but it appears selenium deficiency impairs ability of the immune system to respond to mitogens.
Implications
Nursing calves in southwest Arkansas are at risk for selenium deficiency if dams are not supplemented with selenium. Even when sodium selenite is used in a free-choice mineral supplement designed to deliver 2 mg Se daily, calves are still at risk for selenium deficiency after nursing for approximately 90 days. However, based on WBSe, when cows are supplemented with Sel-PlexTM at the same rate, the risk of selenium deficiency is decreased. Supplementation with Sel-PlexTM before weaning has the ability to increase the percentage of phagocytic macrophages in calves where conditions of selenium deficiency are prevalent.
These changes in immune function occurred even with no differences in GSH-Px between selenium sources. However, large differences in WBSe were observed between selenium sources. The change in macrophage function with supplementation with Sel-PlexTM seems not to be effected through modulation of macrophage activity by the adaptive immune system, because no differences were found in lymphocyte proliferation or in vivo cell mediated immunity. With the improvement in macrophage function of weaned calves, supplementation of cow herds with Sel-PlexTM pre-weaning may lead to reductions in morbidity and mortality of weaned calves compared to calves weaned from Se-deficient dams.
Figure 2. Effect of selenium level and source on calf skin-swelling response to intradermal phytohemagglutinin injection. The effect of selenium supplementation (P=0.12), time after injection (P < 0.01), and time by selenium supplement interaction (P=0.97).
Acknowledgements
This project was conducted with funding from the Arkansas Agric. Exp. Sta., Hatch Project No. AR001735, and gifts from Alltech Inc. (Nicholasville, KY), The Wax Seed Co. (Amory, MS), and Fort Dodge Animal Health (Overland Park, KS). We express our appreciation to P. Capps, B. Kirkpatrick, J. Loe, B. Stewart, J. Weyers, and S. Krumpelman for help in completing these projects.
References
AOAC. 1990. Official Methods of Analysis. 15th ed. Association of Official Analytical Chemists, Arlington, VA.
Olsen, O. E., I.S. Palmer and E.E. Clary. 1975. Modification of the official fluorometric method of selenium in plants. J. Assoc. Off. Anal. Chem. 58:117-121.
Aréchiga, C.F., O. Ortíz and P.J. Hansen. 1994. Effect of prepartum injection of vitamin E and selenium on postpartum reproductive function of dairy cattle. Theriogenology 41:1251-1258.
Arthur, J.R. 1981. Myopathy in selenium deficient cattle. In: Tracer Element Metabolism in Man and Animals. Vol. IV (J.M. Howell, J.M. Grawthorna and C.L. White, ed). Australian Acad. Sci., Canberra, pp. 356-359.
Blecha, F., D.S. Pollmann and D.A. Nichols. 1983. Weaning pigs at an early age decreases cellular immunity. J. Anim. Sci. 56:396-400.
Boyne, R. and J.R. Arthur. 1979. Alterations of neutrophil function in selenium-deficient cattle. J. Comp. Path. 89:151-158.
Bruce, L.B. 1997. Effects of selenium on cold adapted beef cattle. Asian-Aust. J. Anim. Sci. 11:265-567.
Burk, R.F. 1983. Biological activity of selenium. In: Annual Review of Nutrition. Vol. II (W.J. Darby, H.P. Broquist and R.E. Olson, ed). Annual Reviews Inc., Palo Alto, CA, pp. 53-70.
Buskirk, D.D., J. VanLente, T.M. Harrigan, A.J. Zanella, D.R. Hawkins, M. Kaercher and J. Cowley. 2000. In: Evaluation of Hay Loss from Large Round Bale Feeders. Michigan Agric. Exp. Sta. Res. Rep. No. 569. Michigan State Univ., East Lansing, pp. 2-7.
Cao, Y.Z., J.F. Maddox, A.M. Mastro, R.W. Scholz, G. Hildenbrandt and C.C. Reddy. 1992. Selenium deficiency alters the lipoxygenase pathway and mitogenic response in bovine lymphocytes. J. Nutr. 122:2121-2127.
Carter, D.L., M.J. Brown and W.H. Allaways. 1968. Selenium content of forage and hay crops in the Pacific Northwest. Agron. J. 60:532-534.
Chapman, S.L. 1998. Soil Test Recommendations Guide. Univ. Arkansas Coop. Ext. Ser. AGR-9. Little Rock.
Clarkson, M.J., W.B. Faull and J.B. Kerry. 1985. Vaccination of cows with clostridial antigens and passive transfer of clostridial antibodies from bovine colostrums to lambs. Vet. Rec. 116:467- 469.
Dietert, R.R., G.F. Combs, H.K. Lin and J.V. Puzzi. 1990. Impact of combined vitamin E and selenium deficiency on chicken macrophage function. Ann. N.Y. Acad. Sci. 587:281-282.
Erksine, R.J. 1993. Nutrition and mastitis. The Veterinary Clinics of North America. Food Anim. Practice 9:551-561.
Gould, D.H., M.M. McAllister, J.C. Savage and D.W. Hamar. 1991. High sulfide concentration in rumen fluid associated with nutritionally induced polioencephalomalacia in calves. Am. J. Vet. Res. 52:1164-1169.
Gunter, S.A., K.M. Cassida, P.A. Beck and J.M. Phillips. 2002. Winter-annual grasses as a supplement for beef cows. J. Anim. Sci. 80:1157- 1165.
Hidiroglou, M. and K.J. Jenkins. 1975. Effect of selenium and vitamin E, and copper administrations on weight gains of beef cattle raised in a selenium-deficient area. Can. J. Anim. Sci. 55:307-313.
Hoekstra, W.G. 1974. Biochemical role of selenium. In: Trace Element Metabolism in Animals. Vol. II (W.G. Hoekstra, J.W. Suttie, H.E. Ganther and W.Mertz, ed). Univ. Park Press, Baltimore, MD, pp. 61-77.
Koller, L.D., G.A. Whitbeck and P. J. South. 1984. Transplacental transfer and colostral concentrations of selenium in beef cattle. Am. J. Vet. Res. 45:2507-2510.
Kubota, J., W.H. Allaways and D.L. Carter. 1967. Selenium in crops in the United States in relation to selenium-responsive diseases of animals. J. Agric. Food Chem. 15:448-453.
Lawrence, R.A., R.A. Sunde, G.L. Schwartz and W.G. Hoekstra. 1974. Glutathione peroxidase activity in rat lens and other tissues in relation to dietary selenium intake. Exp. Eye Res. 18:563- 569.
McDowell, L.R. 1997. Selenium. In: Minerals in Animal Nutrition. Acad. Press, San Diego, CA.
Monteleone, G., T. Parrello, I. Monteleone, S. Tamarro, F. Luzza and F. Pallone. 1999. Interferon-gamma (INF-γ) and prostaglandin E2 (PGE2) regulate differently IL-12 production in human intestinal laminal propria mononuclear cells (LPMC). Clin. Exp. Immunol. 177:469-475.
Mulhern, S.A., G.L. Taylor, L.E. Magruder and A.R. Vessey. 1985. Deficient levels of dietary selenium suppress the antibody response in first and second generation mice. Nutr. Res. 5:201-210.
Nibbering, P.H., P.C.J. Leijh and R. Van Furth. 1987. Quantitative immunocytochemical characterization of mononuclear phagocytosis. II. Monocytes and tissue macrophages. Immunology 60:171-176.
Nicholson, J.W.G., R.E. McQueen and R.S. Bush. 1999. Response of growing cattle to supplementation with organically bound or inorganic sources of selenium or yeast cultures. Can. J. Anim. Sci. 71:803-811.
NRC. 1996. Nutrient Requirements of Beef Cattle. 7th ed. Natl. Acad. Press, Washington, DC.
Ortman, K. and B. Pehrson. 1999. Effect of selenate as a feed supplement to dairy cows in comparison to selenite and selenium yeast. J. Anim. Sci. 77:3365-3370.
Paglia, D.E. and W.N. Valentine. 1967. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 70:158-169.
Pehrson, B., M. Knutsson and M. Gyllensward. 1989. Glutathione peroxidase activity in heifers fed diets supplemented with organic and inorganic selenium compounds. Swed. J. Agric. Res. 19:53- 56.
Pehrson, B., K. Ortman, N. Madjid and U. Trafikowska. 1999. The influence of dietary selenium as selenium yeast or sodium selenite on the concentration of selenium in the milk of suckler cows and on the selenium status of their calves. J. Anim. Sci. 77:3371-3376.
Phillips, J.M., A.H. Brown, Jr. and R.W. Parham. 1989. Growth of the beef calf with selenium supplementation. Nutr. Rep. Int. 39:537-545.
Pollock, J.M., J. McNair, S. Kennedy, D.G. Kennedy, D.M. Walsh, E.A. Goodall, D.P. Mackie and A.D. Crockard. 1994. Effects of dietary vitamin E and selenium on in vitro cellular immune responses in cattle. Res. Vet. Sci. 56:100-107.
Puls, R. 1989. Mineral levels in animal health: diagnostic data. Sherpa Int., Clearbrook, British Columbia, Canada.
Reamer, D.C. and C.A. Veillon. 1983. Elimination of perchloric acid in digestion of biological fluids for fluorometric determination of selenium. Anal. Chem. 55:1605-1606.
Siddons, R.C. and C.F. Mills. 1981. Glutathione peroxidase activity and erythrocyte stability in calves differing in selenium and vitamin E status. Br. J. Nutr. 46:345-355.
Sirois, P.K., M.J. Reuter, C.M. Laughlin and P.J. Lockwood. 1991. A method for determining macro and micro elements in forages and feeds by inductively coupled plasma atomic emission spectrometry. The Spectroscopist 3:6-9.
Spears, J.W., R.W. Harvey and E.C. Segerson. 1986. Effects of marginal deficiency and winter protein supplementation on growth, reproduction, and selenium status of beef cattle. J. Anim. Sci. 63:586-594.
Stabel, J.R., B.J. Nonnecke and T.A. Reinhardt. 1990. Effect of in vitro selenium repletion on bovine lymphocyte proliferation. Nutr. Res. 10:1053-1059.
Steel, R.G.D. and J.H. Torrie. 1980. Principles and Procedures of Statistics: A Biometrical Approach. McGraw-Hill Book Co., New York.
Turner, R.J., L.E. Wheatley and N.F.G. Beck. 1985. Stimulatory effects of selenium on mitogen responses in lambs. Vet. Immunol. Immunopathol. 8:119-124.
Vandervelde, K., R. Wallace and W. Singleton. 1990. Reproduction. In: BIF Guidelines for Uniform Beef Improvement Programs (F.H. Baker ed)a. 6th ed. Beef Improv. Fed. Oklahoma State Univ., Stillwater.
van Heugten, E.J., J.W. Spears and M.T. Coffey. 1994. The effect of dietary protein on performance and immune response in weanling pigs subjected to an inflammatory challenge. J. Anim. Sci. 72:2661-2669.
van Vleet, J.F. 1980. Current knowledge of selenium-vitamin E deficiency in domestic animals. J. Am. Vet. Med. Assoc. 176:321-326.
Wagner, J.J., K.S. Lusby, J.W. Oltjen, J. Rakestraw, R.P. Wettermann and L.E. Walters. 1988. Carcass composition in mature Hereford cows: estimation and effect on daily metabolizable energy requirements. J. Anim. Sci. 66:603-612.
Weiss, W.P., H.R. Conrad and N.R. St.-Pierre. 1992. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed Sci. Technol. 39:95-110.
Wright, P.L. and M.C. Bell. 1966. Comparative metabolism of selenium and tellurium in sheep and swine. Am. J. Physiol. 211:6-10.
Authors: STACEY A. GUNTER1, PAUL A. BECK1, TROY J. WISTUBA2, MARI E. DAVIS2 and J. M. PHILLIPS1
1 Southwest Research and Extension Center, University of Arkansas, Arkansas, US
2 Department of Animal Science, University of Arkansas, Fayetteville, Arkansas, US











