Sel-Plex in physiological stability of broiler chickens
Involvement of Sel-Plex in physiological stability and performance of broiler chickens
Published: December 12, 2007
By: FRANK W. EDENS - North Carolina State University (Courtesy of Alltech Inc.)
Incorporation of essential trace elements into the diets of all animals is required for maintenance of health, growth, and myriad biochemicalphysiological functions (Scott et al., 1982). Among these essential trace elements is selenium (Se). Selenium was discovered by Berzelius in Sweden in 1818, but a biological significance of this element was not recognized until it was identified as the toxic agent associated with ‘alkali disease’, now termed selenosis, in the Dakota and Wyoming territories in the United States in 1856 (Franke, 1934).
Unfortunately, the selenosis was due to extremely high concentrations of selenium in soils and plants, and it was considered a dangerous element until 1957. In 1957, Schwarz and Foltz reported that selenium was an essential trace nutrient, and nutritionists then initiated extensive studies to discover the metabolic function of this element and document the consequences of its deficiency in human and animal foodstuffs. Selenium deficiency affects animals and humans and can manifest itself in many diseases and dysfunctions such as liver necrosis, muscular dystrophy, microangiopathy, exudative diathesis, pancreatic fibrosis, poor feathering, retained placenta, mastitis, cystic ovaries, general unthriftiness, Keshan disease, Kashin-Beck disease, cancer, numerous heart diseases, immune deficiencies, reduced fecundity, and many others.
The discovery that glutathione peroxidase contained an integral and stoichiometric quantity of selenium demonstrated a biochemical role for this essential trace element and provided a tool for monitoring its status in all animals (Rotruck et al., 1973). Because of similarities between selenium and sulfur, it has been a long held belief that selenium would follow the sulfur pathways in its metabolism. This concept was strengthened by the discovery that plants and bacteria metabolize selenium to the organic selenomethionine and selenocysteine (Burnell and Whatley, 1977). Selenomethionine is readily utilized as a substrate by enzymes that use methionine, and selenomethionine may be more available than pure methionine (Markham et al., 1980).
However, the multiple roles played by selenium in the maintenance of the homeostatic condition in animals are still being discovered (Arthur and Beckett, 1994). Clinical cases of selenium deficiency are clearly recognized and are easily treated today. However, selenium nutrition in both humans and food production animals appears to be less than optimal in many parts of the world (Rayman, 2000).
THE FUNCTIONALITY OF NATURAL ORGANIC SELENIUM
It has been recognized for more than a quarter century that the selenoamino acids, selenomethionine, selenocysteine and selenocystine, are the primary sources of naturally-occurring selenium in plant-based (Burk, 1976) and meat-based (Levander 1986; Cai et al., 1995) feed ingredients. The selenoamino acids are bound in protein, principally as selenomethionine and selenocysteine and constitute 50 to 80% of the total selenium in plants, grains (Butler and Peterson, 1967) and in Sel-PlexTM, the organic seleniumenriched yeast (Kelly and Power, 1995).
Animals can not synthesize selenomethionine, the primary selenoamino acid, directly from the selenite or selenate forms of inorganic selenium (Cummins and Martin, 1967; Sunde, 1990). However, selenocysteine can be found in the body of animals fed inorganic selenium such as selenite and selenate. The presence of selenocysteine is due to synthesis of glutathione peroxidase and other selenoproteins in which the selenocysteine is incorporated. The selenium in selenocysteine is incorporated cotranslationally using selenide and serine as precursors.
The synthesis of selenocysteine involves a unique process in which selenide is phosphorylated under the influence of selenophosphate synthetase to selenophosphate. The selenophosphate is made available to a unique seryl-tRNASEC that is recognized by selenocysteine synthetase. The selenocysteine synthetase converts seryl-tRNASEC to selenocysteyl-tRNASEC that allows insertion of selenocysteine into a peptide chain. The base triplet UGA that normally functions as a stop codon (Amberg et al., 1996) encodes this process of selenocysteine insertion at its appropriate site in the peptide. The selenocysteine insertion also requires a specific mRNA, an elongation factor, GTP, and the selenocysteine insertion sequence that all interact at the ribosome to read the UGA selenocysteine codon (Low and Berry, 1996). This synthesis and conversion process is energetically expensive.
Selenomethionine is easily converted to selenocysteine via cystothionase (Esaki et al., 1981). Selenomethionine is required by poultry and other animals if there is a conversion of selenomethionine to selenocysteine because they cannot synthesize cysteine de novo (Cummins and Martin, 1967; Esaki et al., 1981). Selenocysteine can be substituted for cysteine in many proteins, but it is not incorporated directly into specific selenoproteins (Sunde, 1990; Daniels, 1996).
In order for selenocysteine to be incorporated into specific selenoproteins, there is a requirement for the selenocysteine-ß-lyase reaction with free selenocysteine to release selenide in the presence of reducing agents (Sunde, 1990; Burk, 1991). Then, selenocysteyl-tRNA[Ser]SEC, which recognizes the specific UGA stop codons in the selenoprotein-mRNA, inserts the new, co-translationally synthesized selenocysteine into the specific selenoprotein (Burk, 1991). Thus, organic selenium must be converted from its original organic to the inorganic form then back to the organic form to fulfill its biological function (Arthur, 1997). This conversion is crucial with regard to synthesis of selenoproteins because it has been reported that 30 to 80% of the selenium in the body may be selenocysteine (Hawks et al., 1985).
Nevertheless, selenomethionine can be a highly available substrate for many proteins and can substitute for methionine in their structure (Daniels, 1996). In cases where supplementation of selenomethionine is at a high level in feeds, it can be demonstrated that 40 to 50% of total body selenium as selenomethionine can be found in muscle (Daniels, 1996). Selenocysteine is the pivotal amino acid in the synthesis of selenium-dependent cytosolic glutathione peroxidase (Rotruck et al., 1973), but only about 30% of the body’s selenium is incorporated into cytosolic glutathione peroxidase. About 70% of total selenium is incorporated into 30 to 100 other selenoproteins in mammals (Burk and Hill, 1993). Therefore, these observations have raised a question about the possibility that selenomethionine, due to its essential nature in the synthesis of selenoproteins and its conversion to selenocysteine, may be the twenty-first amino acid.
THE PROBLEM WITH INORGANIC SELENIUM
Lyons (1996) quoted Don Mahan from Ohio State University and projected that in less than five years, sodium selenite would no longer be supplemented in animal feeds because it is an oxidant. Even though there is an everincreasing database that shows the essential nature of organic selenium in animals, sodium selenite still is used principally as the source of selenium in animal feeds. This seems paradoxical because sodium selenite has a documented pro-oxidant influence (Hafeman et al., 1974; Csallany and Menken, 1986; Spallholz, 1997; Terada et al., 1999). Edens (1996) presented information that illustrated the negative pro-oxidant influence of sodium selenite on breast meat quality.
POSITIVE RESPONSES TO SEL-PLEX BY BROILER CHICKENS
Selenized yeast (Sel-Plex), as a source of selenium in an organic form for broiler production, was recently approved (June 2000) for use in the United States. Selenium is an essential trace element that traditionally has been supplemented in animal feeds in an inorganic form as sodium selenite. Sel- Plex provides a cocktail of selenium compounds (Kelly and Power, 1995), but selenomethionine in the yeast cellular protein component is the primary form of selenium (~50%) in Sel-Plex.
The organic selenium profile in Sel- Plex is similar to the organic selenium profile in plants and grains (Kelly and Power, 1995). The organic selenium in Sel-Plex is readily available and is actively absorbed from the intestine via the Na+-dependent neutral amino acid pathway while selenite is absorbed passively (Schrauzer, 2000).
Additionally, it has been shown that Sel-Plex is superior to sodium selenite in terms of induction of feathering, causing sex-linked slow feathering broiler males and normal feathering females to increase feather development (Edens et al., 2000), in tissue accumulation and retention in broilers (Norheim and Moksnes, 1985), and in reducing drip loss from breast meat (Edens, 1996, Naylor et al., 2000). Not only can Sel-Plex improve broiler meat quality as a consequence of better water holding capacity and less drip loss, but it can also reduce the incidence of pale soft, exudative meat from broilers (Edens, in review; Figure 1).
The many positive responses to the presence of Sel- Plex in feed has increased interest in the use of Sel-Plex in all sectors of the poultry industry and is driving research in several diverse areas. Therefore, it is important not only to continue the comparison of the influence of selenium sources (Sel-Plex vs. sodium selenite) on production performance of broilers but also to document the influence of organic selenium on physiological and biochemical stability of the broiler chicken.
CURRENT RESEARCH WITH ORGANIC SELENIUM IN BROILERS: PERFORMANCE, YIELD AND PHYSIOLOGICAL RESPONSES
This paper will review findings from research conducted with high meat yielding sex-linked slow feathering male broiler chickens. The objectives of these studies were to 1) determine the influence of selenium source on production performance of broilers, 2) determine the influence of selenium source on cut-up yields from carcasses of broilers, 3) determine the influence of selenium source on drip loss from breast meat of broilers, 4) determine the influence of selenium source on serum thyroid hormone levels in broilers, and 5) evaluate the potential anti-stress properties of organic selenium from Sel-Plex.
METHODS AND MATERIALS
COMPARATIVE EFFECTS OF SEL-PLEX AND SELENITE ON PERFORMANCE, FEATHERING, THYROID HORMONES AND CARCASS COMPOSITION
Several experiments were conducted to evaluate the influence of Sel-Plex on production performance of auto-sexing slow feathering cockerels (Arbor Acres X Arbor Acres). Some of the studies were conducted in conventional broiler production facilities, some were conducted in cages (Havenstein et al., 1998), and some of the studies were conducted in battery units. Typically a minimum of 40 cockerels were placed in either floor pens or in cage pens (0.9 square feet per bird). In battery trials, cockerels were placed at a density of 0.75 square feet per bird until six weeks of age.
The experimental diets consisted of the North Carolina Agricultural Research Service standard starter diet- 3177 kcal/kg ME, 22.5% CP (1-16 days of age), grower diet- 3168 kcal/kg, 19.5% CP (16-35 days of age), and finisher diet- 3160 kcal/kg, 17.5% CP (35-42 days of age). The diets were supplemented with organic selenium yeast (Sel-Plex, Alltech, Inc.) or sodium selenite at 0.2 mg Se/kg of feed. The background levels of selenium were 0.28 mg/kg, 0.28 mg/kg, and 0.24 mg/kg, respectively, in the starter, grower and finisher diets. In the production studies, the starter, grower, and finisher diets were provided to allow per bird consumption of an average of 2 lbs of starter, 5 lbs of grower and 2 lbs of finisher. A typical experiment used a 2 x 2 factorially arranged completely randomized statistical design. Selenium was added to the experimental diets as follows: Diet 1: no supplemental selenium; Diet 2: sodium selenite at 0.2 ppm Se; Diet 3: Sel- Plex at 0.2 ppm Se; Diet 4: sodium selenite (0.1 ppm Se) + Sel-Plex (0.1 ppm Se); feeds were analyzed for levels of selenium at a commercial AAOCcertified laboratory.
In production performance investigations, the following parameters were measured: body weights at 2, 4, and 6 weeks of age, feed conversion ratio (FCR, feed/gain) through six weeks of age, mortality, cut up parts yield as a percentage of carcass weight (40/treatment) and drip loss from breast meat at six weeks of age (20/treatment). Dissected and identified pectoralis major was ice water-chilled for 4 hrs, blotted dry, and a one-inch cube was cut and weighed. The cube was suspended from a stainless steel hook attached to the weight-tared lid of a weight-tared jar. The jar/lid unit was closed, and on a period of 24 hrs, the weight-tared jar was weighed with the lid in place and with the lid removed. By gravimetric analysis, drip loss was determined by weight changes in the lidless, weight-tared jar. Serum thyroid hormones (thyroxin [T4] and triiodothyronine [T3]) were determined with the non-radioactive Morningstar ELISA assay (Naperville, IL).
EFFECT OF SEL-PLEX ON PHYSIOLOGICAL RESPONSES OF BIRDS SUBJECTED TO E. COLI CHALLENGE OR HEAT DISTRESS
Studies were conducted in which Sel-Plex was evaluated without comparison to selenite, but evaluated the influence of supplemented Sel- Plex to a basal corn/soybean-based diets. Newly hatched male broiler chickens (Arbor Acres x Arbor Acres) were obtained from the North Carolina Agricultural Research Service Poultry Field Research Laboratory hatchery and reared in an environmentally-controlled isolation facility. The chicks were placed in heated brooder-grower batteries with raised wire floors. Brooding temperature was 95 °F (35 °C), and this temperature was decreased incrementally (4 °C at a time) to 70 °F (20 °C) by the time the birds were 21 days old. The photoperiod was 23 hrs light and 1 hr darkness.
The experimental diets and background level of selenium in the ingredients was the same as in the previously described trials. Body weights and feed conversions (FCR) were determined at 21 and 42 days of age (42 day data are presented here), and mortality was recorded on a daily basis.
Two different 2 x 2 factorially arranged studies were conducted involving 60 chicks per treatment group in each study. The first involved a day one post-hatch intranasal challenge by an enteropathogenic Escherichia coli, and the second involved an acute heat challenge to chicks that had been raised in a thermoneutral environment.
The enteropathogenic E. coli (serotype O1, EPEC) was grown overnight at 37 °C in trypticase soy broth. The EPEC challenge dose (106 CFU/mL) was administered in a volume of 100 mL into the external nares to day-old chicks. There were four treatment groups: 1) no supplemental selenium, no EPEC (sterile trypticase broth); 2) Sel-Plex, no EPEC (sterile trypticase soy broth); 3) no supplemental selenium + EPEC; 4) Sel-Plex + EPEC. The EPEC challenge was boosted on days 2, 3, and 4 post-hatch by adding to the drinking water 15 mL of the EPEC culture (10 × 106 CFU).
Heat distress treatment groups were as follows: 1) Control (no supplemental selenium, no heat distress) 2) Sel-Plex, no heat distressing 3) Heat distress control (no supplemental selenium + heat distress) and 4) Sel-Plex + heat distress. At 42 days of age, 20 birds per group were subjected to heat distress by a 1hr exposure to an elevated ambient temperature (40 °C) in a temperature controlled chamber. Mortality was monitored, but there were no deaths during the short exposure period.
In each of these two separate studies, 20 birds per treatment group were killed by carbon dioxide asphyxiation. In the EPEC challenge study, a 1 g sample of the liver and two 1 g ileum samples were dissected and placed into ice-cold protein buffer. The tissues were washed three times to rinse away blood and were then homogenized. Supernatants were collected after a 1 hr 100,000 x G centrifugation.
The total protein content was determined for each sample. Total, oxidized and reduced glutathione, and the reduced: oxidized glutathione ratios (R:O) were determined in ileum (EPEC challenge study) and blood (heat distress study) using colorimetric assays modified for a plate reader (OXIS International, Inc., Portland, OR). The heat shock protein 70 (HSP70) determinations were conducted with procedures using an enzyme-linked immunosorbent assay (Anderson et al., 1993). The data were analyzed as a completely randomized design using the general linear models procedure of the Statistical Analysis System (SAS, 1996). When significant (P<0.05), main effect means were separated using least significant difference (LSD; SAS, 1996).
RESULTS AND DISCUSSION
PALE SOFT AND EXUDATIVE BREAST MEAT
Pale, soft, exudative (PSE) meat is a growing problem among high meat yielding animals. It has been studied extensively in swine and in turkeys (Ferket and Foegeding, 1994). Ferket and Foegeding (1994) have demonstrated that a great amount of the PSE problem in turkeys can be controlled by supplementation of high levels of vitamin E in the feed, suggesting that there may be an oxidative problem associated with the post-mortem development of this condition. Generally, PSE develops when there is accelerated post-mortem glycolysis and rapidly decreasing pH in meat that is still warm. These conditions result in meat that is pale with decreased water holding capacity and poor texture (Ferket and Foegeding, 1994).
Data presented in Figure 1 illustrate that Sel-Plex was more efficient than sodium selenite in reducing the frequency of PSE breast meat from chickens reared in spring and summer. The lower level of Sel-Plex (0.1 ppm Se) was equivalent to, or better than, the higher level (0.3 ppm Se) from selenite in reducing the frequency of PSE meat. These data clearly demonstrate that there may be an oxidative stress that contributes to the development of PSE meat.
In both spring and summer, selenite-fed broilers were found to be more susceptible to PSE development than Sel-Plex-fed broilers. The data are somewhat confusing, however, because the higher rate of selenite supplementation reduced the frequency of PSE samples compared to the lower level of selenite supplementation. On the other hand, the lower level of Sel-Plex was as effective as the higher level of selenite in the reduction of PSE. Sel-Plex at the higher level of supplementation (0.3 ppm Se) was very effective in reducing the frequency of PSE-susceptible meat samples.
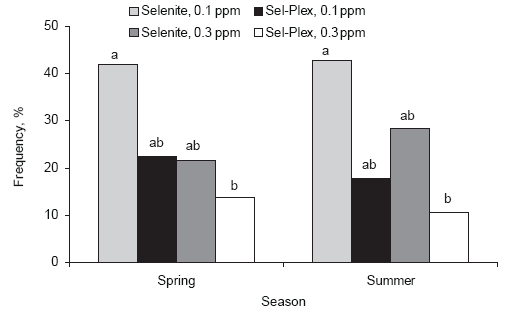
Figure 1.Influence of selenium source and level on distribution frequency of PSE in broiler breast meat. Unlike superscripts indicate significant difference, (P < 0.05).
These observations suggest that superior tissue retention of organic selenium and steady state release of organic selenium for incorporation into the glutathione/glutathione peroxidase antioxidant system was important in the reduction of the PSE condition in broiler chicken meat.
SEL-PLEX INFLUENCE ON BROILER PRODUCTION PERFORMANCE
Background levels of selenium in the unsupplemented basal diet averaged 0.26 ppm during the course of these studies. The background level of selenium represented selenium in the organic form. This background level should have been sufficient to sustain normal broiler chickens based upon the 0.1 ppm Se requirement reported by the National Research Council (1994). It is possible that much of the selenium in the feed could have been unavailable because it was complexed with heavy metals from various sources or that the organic selenium was in a metabolic form that was unavailable to the chicken. Nevertheless, the following data suggest that there may be an additional requirement for selenium by the faster growing, higher yielding broilers that are common today, and that organic selenium may be more useful than inorganic forms of selenium.
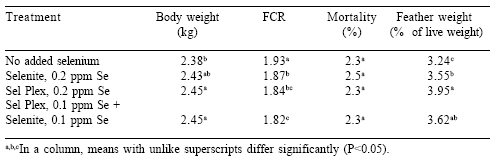
Performance was similar in both experiments and data were pooled. Performance was improved in broilers fed Sel-Plex as compared to birds fed no supplemental selenium or fed sodium selenite (Table 1). Body weight at 42 days was increased in Sel-Plex-fed broilers as compared to those given no supplemental selenium or birds fed sodium selenite. The combination of sodium selenite and Sel-Plex did not improve body weight over the Sel- Plex group. Feed conversions (FCR) were improved by all selenium sources, with the Sel-Plex and Sel-Plex + sodium selenite treatments superior to sodium selenite only. Mortality was not reduced by either selenium source. However, total yield of feathers was increased from those birds given Sel- Plex as compared to all other selenium treatments.
Table 1. Influence of selenium source on body weight, feed conversion, mortality and feather weight.
Body weight improvements were evident in the Sel-Plex-fed chickens as early as 2 weeks of age but were improved significantly at 4 and 6 weeks compared to sodium selenite and no supplemental selenium groups (Figure 2). The data show that the sodium selenite-fed broilers lag behind the Sel- Plex-fed birds until 6 weeks of age, presumably because they were slower to feather during the first 5 weeks of age (Edens et al., 2000). Even though background levels of selenium were sufficient to sustain good performance by the broilers involved in this study, additional selenium appeared to be necessary to optimize growth with birds given Sel-Plex responding to a greater degree. Feed conversions were improved by both selenium sources when compared to FCR for broilers given no supplemental selenium (Figure 3). The data suggest that an additional requirement for selenium may be necessary for the modern broiler chicken, and Sel-Plex appeared to be better than sodium selenite in meeting the additional requirement.
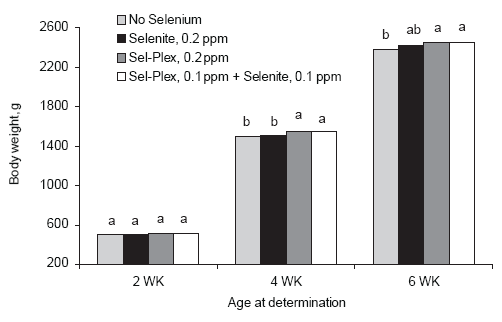
Figure 2.Influence of selenium source on body weight. Unlike letters associated with bars on the histogram indicate a significant difference (P < 0.05).
The data in this investigation have shown that organic selenium provided as Sel-Plex improves the performance of broiler chickens grown in cages. There was improved body weight at 42 days of age in groups of broilers fed Sel-Plex and the combination of Sel-Plex plus selenite as compared to selenite alone or to birds fed a diet with no supplemental selenium. These observations were remarkable considering the fact that there was an average background of 0.26 ppm Se in the basal diet. Given the statement by the National Research Council (1994) that broiler chickens have a requirement of only 0.1 ppm Se, the birds on the basal diet should have performed optimally. A 42 day body weight of 2.38 kg is not a bad body weight, and a feed conversion of 1.93 is not a very bad FCR.
However, provision of selenite improved body weight by an additional 50 g and improved FCR by 6 points, or 3.1% over that for the birds with no supplemental selenium. The feeding of Sel- Plex further improved body weight by 70 g over the broilers with no supplemental selenium and improved FCR by 8 points or 4.7%. These data suggest that the basal diet with 0.26 ppm Se background was not sufficient to optimize growth of the high yielding broilers in this investigation. Additional selenium was needed for optimum growth, and the provision of selenite at 0.2 ppm Se was beneficial, but not as beneficial as Sel-Plex at 0.2 ppm Se.
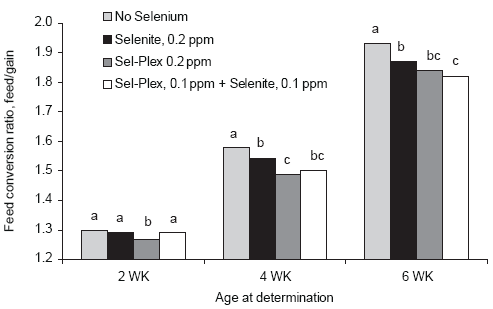
Figure 3.Influence of selenium source on feed conversion. Unlike letters associated with bars on the histogram indicate a significant difference, (P < 0.05).
SEL-PLEX INFLUENCE ON FEATHERING IN AUTO-SEXING SEX-LINKED SLOW FEATHERING BROILER COCKERELS
Improved feather growth can be observed in Sel-Plex-fed auto-sexed sexlinked slow feathering males and normal-feathering females as early as 3 weeks of age as compared to both male and female broilers fed sodium selenite (Figure 4). The feathering process is energetically demanding, but broilers fed Sel-Plex are capable of producing faster growing feathers without loss of performance. In fact, with the earlier feathering, there is a change in the growth pattern and by market age, the Sel-Plex-fed broilers showed significant improvements in performance as compared to sodium selenitefed broilers.
Improved feathering due to Sel-Plex feeding has been reported previously (Edens, 1996; Edens et al., 2000). Downs et al. (1999) reported improved back feathering in Sel-Plex-fed 28 day old broilers. That feathering rate was increased with Sel-Plex without loss of performance is remarkable.
Feather growth is an energetically demanding process that requires a significant allocation of nutrients. These nutrients are drawn from metabolic and dietary pools that could be utilized in other body systems and nutrients that could be stored. Kuenzel and Kuenzel (1977) partitioned layer and broiler-type chicken basal metabolic activity into productive energy usage (protein accretion, feathering, egg production, growth) and maintenance energy (difference between energy intake and energy lost to the environment). Their work indicated that layer stock had higher metabolic demands during the first eight weeks than did broiler stock.
However, when the data were manipulated on the basis of metabolic body size, the differences were apparent only through the first three to four weeks. This is important because feather cover is almost completed by three weeks of age in the layer type but not until nearly six weeks of age in the broiler type. After feather cover is established in layer and broiler type chickens, basal metabolic activity declines in both types. Using the data generated by Kuenzel and Kuenzel (1977), the cost of feather production (kJ/g) would be roughly 167 kJ (29% of daily energy intake) for the layer type and 251 kJ (38% of daily energy intake) for the broiler type. These values compare with 104 kJ/g dry feathers for the molting kestrel (Falco tinnunculus), which had a resting basal metabolic rate (BMR) of 108kJ/g (Dietz et al., 1992); 505 kJ/g (20 to 40% of BMR) in white-crowned sparrows (King, 1981); 588 kJ/g (23% of BMR) in Chaffinches (Gavrilov, 1974).
However, during periods of feather growth, the elevated productive energy is not completely allocated to feather growth, but is also used to maintain body temperature. To accomplish this physiologically important function, thyroid hormone levels increase, and the bird increases heat production. As feather cover increases, the BMR falls and heat production is diminished because oxidative metabolism decreases.
Involvement of thyroid hormones in feathering has long been recognized (Radi and Warren, 1938; Parker, 1943; Boone et al., 1950). Feeding thyroprotein increases feathering rate in slow feathering birds. In thyroprotein there is a predominance of thyroxine (T4), but in poultry, triiodothyronine (T3) is the most active thyroid hormone, and it is known that T3 is intimately involved in feather development. A small quantity of T3 is produced in the thyroid, but the greatest quantity is converted from T4 to T3 in the liver by the type I 5'-deiodinase enzyme that predominates in the chicken (May, 1989). The type I 5'-deiodinase enzyme derives its biochemical properties from selenocysteine (Berry et al., 1991).
The use of Sel-Plex during the first few weeks of life in the broiler apparently improves feathering, but more importantly, the provision of organic selenium may also play a significant role in antioxidant functions in the growing broiler. Roch et al. (2000) have demonstrated that adding Sel-Plex to broiler diets caused a significant increase in glutathione peroxidase (GSHPx) activity in comparison to selenite feeding. If high yielding broiler chickens are susceptible to the damaging effects of free radical production that increases during the non-feathered condition frequently associated with autosexing-slow feathering male broiler chickens, then the provision of Sel- Plex to induce elevated GSH-Px activity should prevent much of the cellular damage caused by those free radicals. The data from this study strongly suggest that Sel-Plex must be inducing biochemical and physiological reactions that improve metabolism, possibly due to less free radical damage.
This would explain improved feathering without loss of performance as indicated by improved body weights and feed conversion ratios.
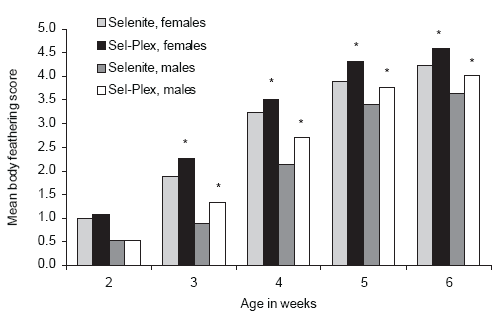
Figure 4.Effect of dietary selenium on whole body feathering of broiler chickens. (* P< 0.05)
SEL-PLEX INFLUENCE ON CUT-UP YIELDS OF HIGH-YIELDING BROILER COCKERELS
On a percentage of carcass weight, yields of viscera, feet, and neck were higher in birds given Sel-Plex (Table 2). Yields of leg and pectoralis major, as percentages of carcass weight, were increased and decreased slightly, respectively, in birds given Sel-Plex (Table 3). In addition, there was a trend toward higher yield of thigh meat from the group given Sel-Plex.
Table 2. Influence of selenium source on percentage yield of parts based on carcass weight.
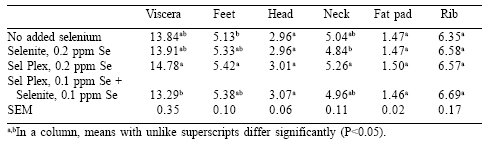
Table 3. Influence of selenium source on percentage yield of parts based on carcass weight.
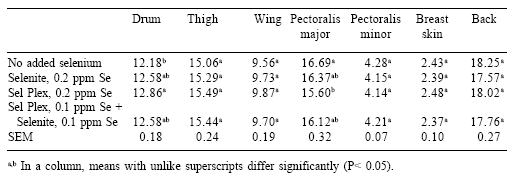
Cut-up yields of carcasses were influenced by selenium source in this study. A significant increase in the yields of viscera, feet and neck as a percentage of carcass and live weight in Sel-Plex-fed animals was somewhat surprising. A larger viscera weight might indicate that feed passage could be somewhat slower and feed retention in the gut could be longer.
The result of this condition possibly could be more efficient utilization of feed. Such was the case in this study and in others (Naylor et al., 2000; Roch et al., 2000) where improved FCR was reported. Increased yields of feet and neck may also reflect improved growth in the Sel-Plex-fed broilers. Increased yields of feet has an added benefit in parts of Asia where the feet are sold at a premium. Increased neck yields also can be viewed as a resource for novel/ niche food or for deboned flesh for value-added products. Leg yields were increased with the feeding of Sel-Plex confirming earlier observations (Edens, 1996; Naylor et al., 2000). Additionally in this study, thigh yields tended toward increased yields with Sel-Plex, and this was consistent with earlier observations (Edens, 1996; Naylor et al., 2000).
However, one of the more perplexing and negative aspects of Sel-Plex feeding is the reduced yields of pectoralis major muscle in the breast of male broilers. This negative response was first reported by Edens (1996), but this loss of pectoralis major muscle was small and may not be of major consequence. It may be possible to improve the yield of the breast muscle by making minor changes in the starter diet by adjusting arginine:lysine ratios or by being cognizant that the source of methionine may also play a role in the subsequent muscle yield of the breast. On the other hand, Naylor et al. (2000) found an increase in the yield of total breast from broilers fed Sel-Plex.
SEL-PLEX INFLUENCE ON BREAST MEAT DRIP LOSS
Selenite feeding was associated with increased breast meat drip loss as compared to both the Sel-Plex and no supplemental selenium treatments (Figure 5). The data suggest that selenite selenium may be associated with an oxidative process that promotes postmortem-development of compromised cell membranes and facilitates increased moisture loss from processed breast meat. Mahan (1999) reported a similar observation in swine.
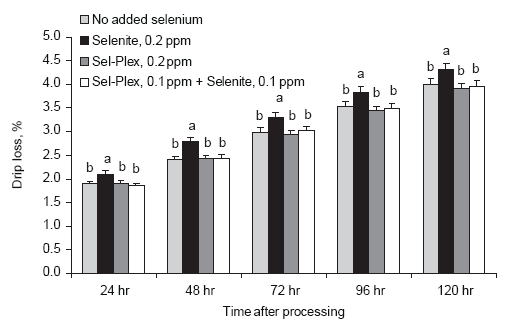
Figure 5. Effect of selenium source on drip loss from male broiler breast meat. Unlike lower case letters associated with bars on the histogram indicate a significant difference (P < 0.05).
BROILER RED BLOOD CELL MEMBRANE INTEGRITY MAY BE AN INDICATOR OF POTENTIAL DRIP LOSS
A recent observation (Edens, 2001, unpublished) has shown that red blood cell membrane resistance to osmotic stress was improved in broilers fed Sel-Plex (Figure 6). This observation is important because it illustrates the potential for decreasing drip loss in processed poultry meat. During processing, whole carcasses are often chilled in an ice bath. The ice bath is hypotonic to the intracellular compartment of the muscles and other cell types. During this chilling process, large quantities of fluid cross the cell membrane and enter the cell by simple osmotic movement. Muscle cells are no different from other somatic cells, such as blood cells. Eventually, the osmotic stress induced by the hypotonic ice water can cause muscle cells to swell and rupture. When this occurs, intracellular fluids leak out and appear as drip loss in the packages of processed meat. Therefore, improved cellular integrity owing to improved cellular selenium status could be reflected as decreased drip loss from processed meat.
Furthermore, selenite selenium has been shown to be an oxidant in certain conditions (Terada et al., 1999). Selenite selenium in the presence of cysteine in total parenteral nutrition caused endothelial damage resulting from generation of reactive oxygen metabolites while selenomethionine and selenate selenium did not cause such damage. Selenite selenium reacted with the cysteine sulfhydryl in the GHS molecule producing reactive oxygen metabolites (Terada et al., 1999).
Therefore, it is possible that free selenite selenium in muscle tissues could also react with thiol groups on other chemical species to generate free radicals that would damage cell membranes leading to increased drip loss from processed poultry and other meat sources. This hypothesis would appear to be supported by the work of Roch et al. (2000) who found increased antioxidant activity due to elevated activities of GSH-Px in Sel- Plex fed broilers.
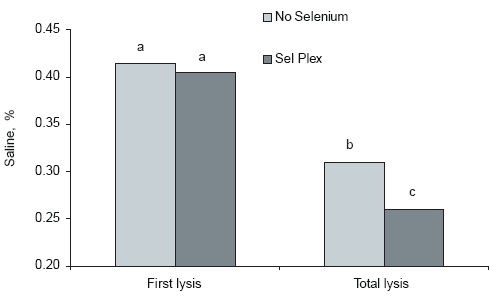
Figure 6.Sel-Plex influence on red blood cell osmotic fragility. Background selenium in the basal feed was 0.28 ppm, and Sel-Plex was added at 0.20 ppm to the basal diet. Unlike letters associated with bars on the histogram indicate significant difference, (P< 0.05).
SERUM THYROID HORMONES REFLECT SELENIUM STATUS IN BROILERS
Serum thyroxin (T4) levels were higher in birds given no supplemental selenium compared to those given selenite or Sel-Plex (Figure 7). There was no difference in serum T4 concentrations from 2 to 6 weeks of age due to due to selenium source.
Serum triiodothyronine (T3) levels were lower in broilers given no supplemental selenium compared to broilers given either selenite or Sel- Plex (Figure 8). Serum T3 in broilers given selenite was lower, but not significantly, than the serum T3 in broilers fed Sel-Plex. Nevertheless, the data do indicate that supplemental selenium was necessary for increased conversion of serum T4 to T3. This was another indication that the background level of 0.26 ppm Se in the basal diet was not sufficient to effectively meet the requirements of the high yielding broiler line used in this study.
The ratios between serum T4 and T3 are shown in Figure 9. Even at two weeks of age, the ratio between these two thyroid hormones was elevated significantly in those broilers given diets without supplemental selenium as compared to those broilers given selenite or Sel-Plex. However, the ratios between these two thyroid hormones suggest strongly that organic selenium supplementation facilitated the conversion of T4 to T3. This observation suggests that there is a good probability that the extra-thyroidal conversion of T4 to T3 was mediated by the hepatic selenium dependent type I, 5’- iodothyronine deiodinase enzyme. Higher tissue levels of selenium appear to be correlated with greater activity of this enzyme.
Values for serum T4 and T3 generated in this study were lower than expected. However, McNabb (2000) reported that serum T4 concentrations in birds can range between 5 and 15 ng/mL and T3 concentrations can range between 0.5 and 4 ng/mL. Although the concentrations for T4 to T3 reported here are low, they are within normal ranges. The thyroid hormones in chickens have been reported to be equipotent as metabolic stimulators, and there are reports that T3 is the more potent of the two hormones (McNabb, 2000). The thyroid hormone data in this study, while not conclusive, do suggest that conversion of T4 to T3 was more efficient when Sel-Plex was the selenium source. Since thyroid hormones are involved in feathering, the slightly higher levels of T3 in Sel-Plex-fed birds may be related to the improved feathering rate seen in birds given Sel-Plex (Edens, 1996; Edens et al., 2000).
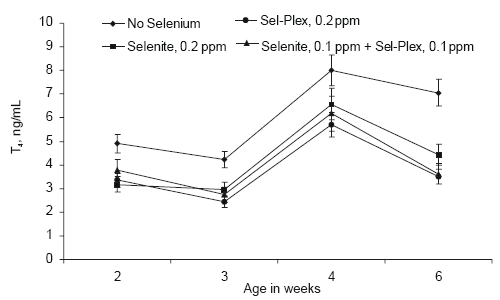
Figure 7.Effect of selenium source on serum thyroxine (T4) in broiler chickens.
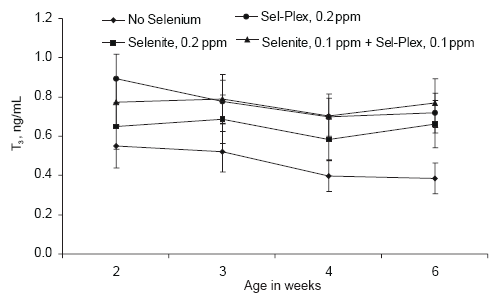
Figure 8.Effect of selenium source on serum triiodothyronine (T3) in broiler chickens.
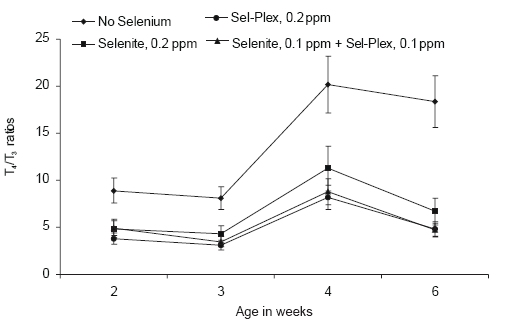
Figure 9.Effect of selenium source on serum T4/T3 ratios in broiler chickens.
SEL-PLEX FUNCTIONS AS AN ANTI-STRESS AGENT IN BROILER CHICKENS
Optimum growth of broilers in a field environment is seldom achieved because many distressful events can be experienced during a brooding and growing period. Such factors include enteric bacterial infections, dietary lipid peroxidation and formation of free radicals, thermal stressors that facilitate oxidative metabolism and subsequent production of reactive oxygen metabolites (free radicals; ROM). These stressors, along with many other factors that induce ROM formation, can have a negative impact on performance, induce immunological reactions that stimulate macrophage activity with subsequent production of hydrogen peroxide and ROM.
Furthermore, interruptions in feed and water supply can also cause stressful reactions and promote oxidative metabolism of body energy stores and subsequent oxidative damage to organs and cells. Many of these problems might be alleviated by the use of certain feed grade antioxidants or vitamin E in the diets of the broilers, but these remedies do not prevent or correct all of the oxidative damage. Thus, it is important that additional feed ingredients such as selenium, which are involved in natural protection of the body against oxidative damage, be provided. Selenium functions in a pivotal role in the production of GSH-Px, a potent cytosolic antioxidant and free radical scavenger in all vertebrate animals.
When animals are exposed to high ambient temperatures or other environmental stressors, they universally alter cellular protein synthesis profiles by elevating several classes of proteins called heat shock proteins (HSP; Ashburner and Bonner, 1979; Craig, 1985; Lindquist, 1986). Induction of HSP results in a depression in the synthesis of normal cellular proteins (Kelly and Schlesinger, 1978; Ashburner and Bonner, 1979; Atkinson, 1981), and this ultimately explains the reason for poor performance in animals grown under the influence of high ambient temperature or other stressors that affect body functions.
Heat shock protein 70 (HSP70) plays a major role in cellular adaptive responses to a number of stressors (Edens et al., 1992; Wang and Edens, 1994, 1998). Exposure of poultry species to mild stressors over a period of time enhances HSP70 expression, but eventually the birds become acclimated and no further increase in cellular HSP70 can be demonstrated (Wang and Edens, 1998). However, one of the negative aspects of induction of HSP70 in response to a stressor is the fact that HSPs are synthesized preferentially to other cellular proteins (Edens et al., 1992; Wang and Edens, 1994, 1998).
It is an accepted fact that selenium is an essential trace element involved in the regulation and control of the body’s antioxidant glutathione and GSHPx system. This system plays a major role in the control of ROM (Palmer and Paulson, 1997). It has been reported that selenium supplementation of chickens improved resistance to the stress caused by E. coli infection and by exposure to cold (Larsen et al., 1997). Thermal extremes and bacterial infection are stressors that increase metabolic activity with a resultant increase in ROM production (Palmer and Paulson, 1997). HSP70 is also involved in mechanisms protecting the body from the deleterious effects of ROM (Favatier et al., 1997; Polla, 1998). Furthermore, there are data that suggest a strong relationship between thiol oxidation and increased HSP70 synthesis in stressed cells (Sierra-Rivera et al., 1994).
Therefore, it was important to explore the possibility that selenium might be playing a role in the induction of heat shock proteins as well as playing a role in maintenance of the glutathione system. Studies were conducted to determine the influence of Sel-Plex supplementation to broiler chickens on the induction of HSP70 and glutathione response either to a bacterial challenge or to an acute heat distress.
There were significant effects due to Sel-Plex and EPEC challenge on performance of broiler chickens (Table 4). Broilers given Sel-Plex were significantly heavier, had improved FCR and lower mortality rates than birds not given a selenium supplement. Additionally, there was a Sel-Plex x E. coli interaction in which Sel-Plex-supplemented broilers with EPEC challenge were significantly heavier, had improved FCR, and lower mortality than EPEC-challenged broilers without the selenium. In the heat distress study, Sel-Plex-supplemented broilers had greater body weight (2291g vs. 2004g), improved FCR, (1.84 vs. 1.98) and reduced mortality (5.7% vs. 18.3%) compared to those broilers with no selenium supplementation. There were no mortalities in broilers subjected to acute heat distress.
Table 4. Performance at 42 days of age of male broiler chickens given a dietary supplement of organic selenium and challenged with an enteropathogenic E. coli.
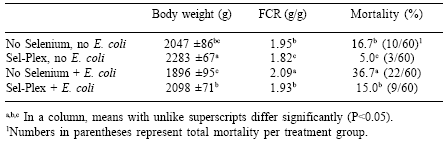
The HSP70 concentrations in the ileum from EPEC-challenged broilers were elevated significantly in both Sel-Plex (Sel-Plex, no E. coli, 0.305c; Sel-Plex + E. coli, 0.806b OD/ng total protein) and no selenium (no selenium, no E. coli, 0.277c; no selenium + E. coli, 1.229a OD/ng total protein) groups. Sel-Plex supplementation allowed a smaller intestinal HSP70 elevation when the broilers were challenged. Hepatic HSP70 was unaffected either by EPEC challenge or by selenium. Compared with unsupplemented birds, Sel-Plex was associated with significantly lower constitutive (0.90c vs 1.04b OD/ng total protein) and inducible (0.93c vs. 1.12a OD/ng total protein) hepatic HSP70 concentrations.
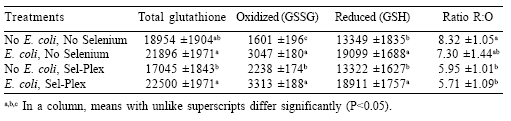
EPEC challenge increased ileum total glutathione content, but selenium did not influence that response (Table 5). Oxidized (GSSG) and reduced (GSH) glutathione in the ileum were elevated significantly by EPEC challenge, and Sel-Plex treatment increased ileum GSSG in both challenged and nonchallenged birds. The concentration of GSH was increased significantly by EPEC challenge but was not affected by selenium. The ratios between GSH and GSSG (R:O) were elevated significantly in those birds given the diets without supplemental selenium in comparison to the ratios for birds in the Sel-Plex groups.
Table 5. Influence of Sel-Plex on intestinal total glutathione (μM/g), oxidized glutathione (μM/g), reduced glutathione (μM/g), and the reduced:oxidized glutathione ratios in broiler chickens given a dietary supplement of organic selenium and challenged with an enteropathogenic E. coli.
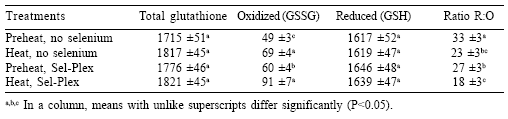
Neither heat distress nor Sel-Plex had any influence on total blood glutathione or GSH concentrations (Table 6). However, blood GSSG was elevated by both heat distress and Sel-Plex supplementation. Both Sel-Plex and heat distress were associated with lower blood R:O ratios.
Table 6. Influence of Sel-Plex on blood total glutathione (μM/L), oxidized glutathione (μM/ L), reduced glutathione(μM/L), and the reduced:oxidized glutathione ratios in broiler chickens given a dietary supplement of organic selenium and subjected to acute heat distress.
The National Research Council established 0.1 ppm Se as the minimum necessary amount to sustain growth and performance in broiler chickens (NRC, 1994); and therefore the background selenium levels in the feeds used in these studies (0.24 to 0.28 ppm) ostensibly were adequate. However, those background levels were not sufficient to allow broilers to attain the same body weight performance as those given Sel-Plex. The FCR and mortality rates also were elevated significantly when the birds were given diets with less than adequate selenium; and those birds ultimately suffered from the development of a stress reaction characterized by elevated tissue concentrations of HSP70. Based upon the HSP70 response, one could postulate that consumption of inadequate selenium conferred the ability to resist some stressors, but that may be an inappropriate conclusion.
The expression of HSP70 is a classical sign of stress in animals because it is the physical manifestation of specific genes that are induced to combat stressors (Edens et al., 1992; Wang and Edens, 1994; 1998; Polla, 1998). However, there is a cost for this response; and it is paid in reduced growth due to inhibition of synthesis of structural proteins in chronically stressed animals (Kelly and Schlesinger, 1978; Ashburner and Bonner, 1979; Atkinson, 1981).
In the studies reported herein, broilers given diets without supplemental selenium had higher constitutive levels of HSP70 than those given organic selenium as Sel-Plex. Additionally, when those animals were subjected either to an EPEC challenge or to an acute heat distress, the inducible HSP70 concentrations were significantly greater in those birds given diets without supplemental selenium. The first impression that could be drawn from these observations might be that selenium inhibited development of the HSP70 response, making birds more susceptible to the deleterious effects of stressors, but the HSP70 and body weight data suggest the opposite. One could also conclude that inadequate dietary selenium caused a stressful condition in those birds. This conclusion is supported by the decreased body weight, elevated FCR and elevated levels of HSP70 noted.
However, the glutathione redox cycle must be considered, because HSP70 expression is responsive to increased thiol oxidation (GSSG production) and loss of cellular GSH as indicated by a lower R:O ratio (Sierra-Rivera et al., 1994). Normally, oxidative stress will induce an increase in blood and tissue concentrations of GSH (Cantor et al., 1982) consistent with the observations reported here. The increased concentrations of GSH can then react with more ROM leading to the formation of GSSG.
This was also found in the studies with EPEC and heat distress and would be consistent with the presence of increased ROM. The increase in GSSG would then induce an elevation in HSP70 that was apparent in those groups of animals given diets with inadequate selenium. However, with Sel-Plex, the ileum HSP70 concentration in EPEC-challenged birds and hepatic HSP70 concentrations in heat-distressed birds were decreased significantly. This was contrary to the expected response, but points to a mechanism whereby lower concentrations of HSP70 indicated improved tolerance to stressors.
Although not measured in these studies, GSH-Px activity was elevated significantly with the addition of organic selenium to poultry diets (Cantor et al., 1982). Reduction of organic peroxides by glutathione is catalyzed by GSH-Px and leads to increased oxidation of GSH to GSSG (Burk et al., 1978). Higher GSH-Px activity in Sel-Plex fed broilers will catalyze a more rapid oxidation of GSH to GSSG, consistent with observations in these studies. Normally, GSSG is reduced back to GSH by GSH reductase. Reduction of GSSG back to GSH can be limited by the GSH reductase activity if there is a rapid build-up of GSSG and marginal levels of riboflavin in diets.
In fact, there was an accumulation of GSSG in the Sel-Plex-fed broilers indicating a more efficient clearing of ROM in those birds. If it is true that accumulation of GSSG increases HSP70 expression (Sierra-Rivera et al., 1994), then tissues from stressed chickens should have shown increased HSP70 concentrations, but that was not the condition in Sel-Plex-fed broilers. It is speculated that early in the Sel-Plex feeding regimen, HSP70 induction may have occurred. However, induction of HSP70 is an acute phase response that cannot be tolerated for long periods of time without resulting in the slowing of growth and performance in animals.
Previous work from this laboratory (Wang and Edens, 1998) has shown that with chronic exposure to heat distress, HSP70 expression decreased as the birds became acclimated. It is hypothesized that the feeding of Sel-Plex has conferred stress resistance by causing an early HSP70 response that later developed into an acclimation condition. In the long term of the six week growing period for Sel-Plex-fed broilers, resistance to stressors was improved as indicated by superior performance traits and a lower HSP70 production.
SUMMARY AND CONCLUSIONS
Selenium is unique among trace minerals because it is the only one with a special mRNA (Allan et al., 1999) and a special tRNA (Commans and Böck, 1999) that are required for its incorporation into an amino acid (selenocysteine). Cysteine and selenocysteine cannot be synthesized de novo in chickens, but these are conversion products from methionine and selenomethionine metabolism. Edens et al. (2000) have shown that provision of selenomethionine in selenized yeast facilitates faster feather development in auto-sexing sex-linked slow feathering males and their normal feathering female siblings.
Studies with selenite and organic selenium in broilers have shown that selenium incorporation into feathers is increased significantly when organic selenium is fed (Edens, 2001, unpublished) suggesting that selenomethionine and selenocysteine may be required for optimized feather growth.
In this study and an earlier investigation (Edens, 1996), less drip loss was observed in breast meat from broilers fed organic selenium. In the processing arena, the carcass is chilled in a hypotonic ice-water bath. The hypotonic water from the ice bath can be absorbed by the flesh of the carcass because the intracellular tonicity of the muscle cell is hypertonic to the ice-water bath, and the cell will swell and in many instances rupture if the amount of absorbed water exceeds the water holding capacity of the cell similar to the rupture of red blood cells in a hypotonic medium. Therefore, increased drip loss can be measured. In cells that may contain large numbers of ROM, cellular membrane integrity can be compromised. This is a high probability in meat from animals fed selenite selenium because selenite selenium is indeed responsible for ROM production. Additionally, a slightly smaller yield of pectoralis major breast muscle and larger yields of leg and thigh have been observed when organic selenium has been provided to broiler chickens.
These observations suggest that cysteine/selenocysteine for feathering may have been diverted from the breast muscle. Furthermore, the natural form of selenium in plants and grains is selenomethionine, and its conversion to selenocysteine or selenocystine for incorporation into seleniumdependent proteins such as glutathione peroxidase and hepatic deiodinase is more efficient.
Finally, the supplementation of organic selenium as Sel- Plex was shown to impart an anti-stress condition in broilers subjected either to EPEC challenge or to acute heat distress. The lower concentration of HSP70 that was generated during these challenges indicated that the broilers were more capable of resisting the rigors of those stressors. Ostensibly, the improved stress resistance in the Sel-Plex-fed animals, during either acute heat stress or as the result of EPEC infection and oxidative damage, was due to more efficient scavenging of ROM than in birds that were not given the Sel-Plex supplement.
Therefore, even with minimal data at this time, one might conclude that selenomethionine may be an essential form of methionine for vertebrate animals such as poultry. The beneficial effects of Sel-Plex on production performance and physiological responses (Table 7) of broiler chickens appear to support this conclusion.
Table 7. Summarized results and conclusions from the four Sel-Plex studies.

REFERENCES
Allan, C.B., G.M. Lacourciere and T.C. Stadtman. 1999. Responsiveness of selenoproteins to dietary selenium. Ann. Rev. Nutri. 19:1-16.
Amberg, R., T. Mizutani, X.Q. Wu and H.J. Gross. 1996. Selenocysteine synthesis in mammalia: An identity switch from tRNA(Ser) to tRNA(SEC). J. Mol. Biol. 263:8-19.
Anderson, R.L., C.T. Wang, I. Van Kersen, K.J. Lee, W.J. Welch, P. Lavagnini and G.M. Hahn. 1993. An Immunoassay for heat shock protein 73/72: Use of the assay to correlate HSP77/72 levels in mammalian cells with heat response. Int. J. Hyperthermia 9:539-552.
Arthur, J.R. 1997. Non-glutathione peroxidase functions of selenium. In: Biotechnology in the Feed Industry. Proc. of the 13th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, Nottingham, UK, pp. 143-154.
Arthur, J.R. and G.J. Beckett. 1994. New metabolic roles for selenium. Proc. Nutri. Soc. 53:615-624.
Ashburner, M. and J.J. Bonner. 1979. The induction of gene activity in Drosophila by heat shock. Cell 17:241-254.
Atkinson, B.G. 1981. Synthesis of heat-shock proteins by cells undergoing myogenesis. J. Cell Biol. 89:666-673.
Berry, M.J., J.D. Kieffer, J.W. Harney and P.R. Larsen. 1991. Selenocysteine confers the biochemical properties characteristic of the type I iodothyronine deiodinase. J. Biol. Chem. 266:14155-14158.
Boone, M.A., J.A. Davidson and E.P. Reineke. 1950. Thyroid studies in fast- and slow-feathering Rhode Island Red chicks. Poultry Sci. 29:195- 200.
Burk, R.F. 1976. Selenium in man. In: Trace Elements in Human Health and Disease (A.S. Prasad and D. Oberleas, eds.). Academic Press, New York, pp. 105-133.
Burk, R.F. 1991. Molecular biology of selenium with implications for its metabolism. FASEB J. 5:2274-2279.
Burk, R.F. and K.E. Hill. 1993. Regulation of selenoproteins. Ann. Rev. Nutri. 13:65-81.
Burk, R.F., K. Nishiki, R.A. Lawrence and B. Chance. 1978. Peroxide removal by selenium-dependent and selenium-independent peroxidases in hemoglobin-free perfused rat liver. J. Biol. Chem. 253:43-56.
Burnell, J.N. and F.R. Whatley. 1977. Sulfur metabolism in Paracoccus denitrificans: Purification, properties, and regulation of cysteinyl- and methionyl-tRNA synthetase. Biochem. Biophys. Acta 481:266-278.
Butler, G.W. and P.J. Peterson. 1967. Uptake and metabolism of inorganic forms of selenium-75 by Spiroldela oligorrhiza. Aust. J. Biol. Sci. 20:77.
Cai, X., E. Block, P.C. Uden, X. Zhang, B.D. Quimby and J.J. Sullivan. 1995. Allium Chemistry: Identification of selenoamino acids in ordinary and selenium-enriched garlic, onion and broccoli using gas chromatography with atomic emission detection. J. Agric. Food Chem. 43:1754-1757.
Cantor, A.H., P.D. Moorhead and M.S. Musser. 1982. Comparative effects of sodium selenite and selenomethionine upon nutritional muscular dystrophy, selenium-dependent glutathione peroxidase and tissue selenium concentrations of turkey poults. Poultry Sci. 61:478-484.
Commans, S. and A. Böck. 1999. Selenocysteine inserting tRNAs: An overview. FEMS Microbiol. Rev. 23:335-351.
Craig, E.A. 1985. The heat shock response. CRC Crit. Rev. Biochem. 18:239-280.
Csallany, A.S. and B.Z. Menken. 1986. Effect of dietary selenite on hepatic organic solvent-soluble lipofuscin pigments. J. Am. Coll. Toxicol. 5:79-85.
Cummins, L.M. and J.L. Martin. 1967. Are selenocystine and selenomethionine synthesized in vivo from sodium selenite in mammals? Biochem. 6:3162-3168.
Daniels, L.A. 1996. Selenium metabolism and bioavailability. Biol. Trace Elem. Res. 54:185-199.
Dietz, M.W., S. Daan and D. Masman. 1992. Energy requirements for molt in the kestel, Falco tinnunculus. Physiol. Zool. 65:1217-1235.
Downs, K.M., J.B. Hess and S.F. Bilgili. 1999. Influence of selenium source on broiler carcass and fillet weep loss. Poultry Sci. 78(Suppl. 1):23.
Edens, F.W. 1996. Organic selenium: From feathers to muscle integrity to drip loss. Five years onward: No more selenite! In: Biotechnology in the Feed Industry. Proc. 12th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, Nottingham, UK, pp. 165- 185.
Edens, F.W. 2001. Influence of selenium source on broiler carcass yield and breast meat drip loss. J. Appl. Poultry Res. (In review).
Edens, F.W., C.H. Hill and S. Wang. 1992. Heat shock protein response in phosphorus-deficient heat-stressed broiler chickens. Comp. Biochem. Physiol. 103B:827-831.
Edens, F.W., T.A. Carter, C.R. Parkhurst and A.E. Sefton. 2000. Effect of selenium source and litter type on broiler feathering. J. Appl. Poultry Res. 9:407-413.
Esaki, N., T. Nakamura, H. Tanaka, T. Suzuki, Y. Morino and K. Soda. 1981. Enzymatic synthesis of selenocysteine in rat liver. Biochem. 20:4492- 4496.
Favatier, F., L. Bornman, L.E. Hightower, E. Günther and B.S. Polla. 1997. Variation in Hsp gene expression and Hsp polymorphism: Do they contribute to differential disease susceptibility and stress tolerance? Cell Stress & Chaperones 2:141-155.
Ferket, P.R. and E.A. Foegeding. 1994. How nutrition and management influence PSE in poultry meat In: Proceedings from BASF Technical Symposium, Multi-State Poultry Feeding and Nutrition Conference, Indianapolis, IN, pp. 64-78.
Franke, K.W. 1934. A new toxicant occurring naturally in certain samples of plant foodstuffs. I. Results obtained in preliminary feeding trials. J. Nutri. 8:597.
Gavrilov, V.M. 1974. Metabolism of molting birds. Zool. Zh. 53:1363-1375.
Hafeman, D.G., R.A. Sunde and W.G. Hoekstra. 1974. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutri. 104:580-587.
Havenstein, G.B., J.L. Grimes, P.R. Ferket, C.R. Parkhurst, F.W. Edens, J. Brake and J.H. van Middelkoop. 1998. Recent experiences with reduced or non-litter systems for growing broilers and turkeys In: Proceedings: 1998 National Poultry Waste Management Symposium. Springdale, AR. October 19-21, pp. 225-240.
Hawkes, W.C., E.C. Wilhelmsen and A.L. Tappel. 1985. Abundance and tissue distribution of selenocysteine-containing proteins in the rat. J. Inorg. Biochem. 23:77-92.
Kelly, M.P. and R.F. Power. 1995. Fractionation and identification of the major selenium compounds in selenized yeast. J. Dairy Sci. 78(Suppl. 1):237.
Kelly, P.M. and M.J. Schlesinger. 1978. The effect of amino acid analogues and heat shock gene expression in chicken embryo fibroblasts. Cell 15:1277- 1286.
King, J.R. 1981. Energetics of avian molt. Proc. Internat. Ornitholog. Cong. 17:312-317.
Kuenzel, W.J. and N.T. Kuenzel. 1977. Basal metabolic rate in growing chicks Gallus domesticus. Poultry Sci. 56:619-627.
Larsen, C.T., F.W. Pierson and W.B. Gross. 1997. Effect of dietary selenium on the response of stressed and unstressed chickens to Escherichia coli challenge and antigen. Biol. Trace Elem. Res. 58:169-176.
Levander, O. 1986. Selenium. In: Trace Elements in Human and Animal Nutrition (W. Mertz, ed.). Academic Press, Orlando, FL, pp 209-280.
Lindquist, S. 1986. The heat-shock responses. Ann. Rev. Biochem. 55:1151- 1191.
Low, S.C. and M.J. Berry. 1996. Knowing when not to stop: Selenocysteine incorporation into eukaryotes. TIBS 21:203-208.
Lyons, T.P. 1996. Goal 2000: A truly global science-based company that responds rapidly to emerging issues. In: Biotechnology in the Feed Industry. Proc. 12th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, Nottingham, UK, pp. 1-20.
Mahan, D.C. 1999. Organic selenium: using nature’s model to redefine selenium supplementation for animals. In: Biotechnology in the Feed Industry. Proc. 15th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, Nottingham, UK, pp. 523-535.
Markham, G.D., E.W. Hafner, C.W. Tabor and H. Tabor. 1980. Sadenosylmethionine synthetase from Escherichia coli. J. Biol. Chem 255:9082-9092.
May, J.D. 1989. The role of the thyroid in avian species. Crit. Rev., Poultry Biol. 2:171-189.
McNabb, F.M.A. 2000. Thyroids. In: Sturkie’s Avian Physiology, 5th edition (G.C. Whittow, ed.) Academic press, New York, pp. 461-471.
National Research Council. 1994. Nutrient Requirements of Poultry, 9th edition. National Academy Press, Washington, DC.
Naylor, A.J., M. Choct and K.A. Jacques. 2000. Effects of selenium source and level on performance and meat quality in male broilers. Poultry Sci. 79(Suppl. 1):117.
Norheim, G. and K. Moksnes. 1985. Distribution and elimination of selenium and glutathione peroxidase (GSH-Px) in chickens after supplementation with sodium selenite or selenomethionine. In: Proceedings: Fifth International Symposium on Trace Elements in Man and Animals (C.F. Mills, I. Bremner and J.K. Chesters, eds.). CAB, Farnham Royal, Slough, UK, pp. 493-495.
Palmer, H.J. and K.E. Paulson. 1997. Reactive oxygen species and antioxidants in signal transduction and gene expression. Nutri. Rev. 55:353- 361.
Parker, J.E. 1943. Influence of thyroactive iodocasein on growth of chicks. Proc. Soc. Exp. Biol. Med. 52:234-236.
Polla, B.S. 1998. A role for heat shock protein in inflammation. Immunology Today 9:134-137.
Radi, M.H. and D.C. Warren. 1938. Studies on the physiology and inheritance of feathering in growing chicks. J. Agri. Res. 56:679-705.
Rayman, M. 2000. The importance of selenium to human health. The Lancet 356:233-241.
Roch, G., M. Boulianne and L. De Roth. 2000. Effect of dietary antioxidants on the incidence of pulmonary hypertension syndrome in broilers. In: Biotechnology in the Feed Industry. Proc. Alltech’s 16th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, Nottingham, UK, pp. 261-276.
Rotruck, J.T., A.L. Pope, H.E. Ganther, A.B. Swanson, D.G. Hafeman and W.G. Hoekstra. 1973. Selenium: Biochemical role as a component of glutathione peroxidase. Science 179:588-590.
SAS Institute. 1996. SAS® User’s Guide: Statistics. SAS Institute, Cary, NC, USA.
Schrauzer, G.N. 2000. Selenomethionine: A review of its nutritional significance, metabolism, and toxicity. J. Nutri. 130:1653-1656.
Schwarz, K. and C.M. Foltz. 1957. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 79:3292- 3293.
Scott, M.L., M.C. Nesheim and R.J. Young. 1982. Nutrition of the Chicken. 3rd Edition. M. L. Scott & Associates, Ithaca, NY.
Sierra-Rivera, E., M.J. Meredith, G.J. Voorhees, L.W. Oberley, D.R. Eisert and M.L. Freeman. 1994. Synthesis of heat shock proteins following oxidative challenge: role of glutathione. Int. J. Hyperthermia 10:573-586.
Spallholz, J.E. 1997. Free radical generation by selenium compounds and their pro-oxidant toxicity. Biomed. Environ. Sci. 10:260-270.
Sunde, R.A. 1990. Molecular biology of selenoproteins. Ann. Rev. Nutri. 10:451-474.
Terada, A., M. Yoshida, Y. Seko, T. Kobayashi, K. Yoshida, M. Nakada, K. Nakada, H. Echizen, H. Ogata and T. Rikihisa. 1999. Active oxygen species generation and cellular damage by additives of parenteral preparations: Selenium and sulfhydryl compounds. Nutrition 15:651-655.
Wang, S. and F.W. Edens. 1994. Hsp70 mRNA expression in heat stressed chickens. Com. Biochem. Physiol. 107B:33-37.
Wang, S. and F.W. Edens. 1998. Heat conditioning induces heat shock proteins in broiler chickens and turkey poults. Poultry Sci. 77:1636-1645.
Author: FRANK W. EDENS
Department of Poultry Science, North Carolina State University, Raleigh, North Carolina, USA
Related topics
Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post
18 de diciembre de 2007
It is good article about performance of broiler chickens







You may be interested in
.jpg&w=3840&q=75)