Poult Enteritis and Mortality Syndrome: Definition and Nutritional Interventions
Poult enteritis and mortality syndrome (PEMS) was first observed in 1991 and has developed into one of the worst diseases to have afflicted commercial turkeys. Until 1994 PEMS was contained in North Carolina, South Carolina and Georgia, but in 1995, PEMS was confirmed as a disease in turkeys in several other states.
PEMS is still a problem in the Carolinas and Virginia, but in 1997, the incidence decreased significantly with only 15 cases being reported in North Carolina. This disease primarily occurs from mid-May through September, but elevated mortality in off-season flocks appears to be a characteristic of the disease on farms that are at risk of developing PEMS.
Generally, from 1992 through 1996, the first seasonal cases of PEMS in North Carolina have been reported in the 20th week of each year and decline rapidly around the 40th week.
PEMS defined and described
PEMS is an acute transmissible, infectious, enteric disease that affects young turkeys between the ages of seven and 28 days of age (Barnes et al., 1996).
However, older birds, even market-age birds, can show signs that resemble PEMS, and these signs include both diarrhea and stunting. PEMS can be differentiated from early poult mortality, stunting syndrome and avian growth depression.
Poults with a PEMS infection become agitated with high-pitched vocalization and constant movement, they refuse to eat normal amounts of feed and behave as if even feed or water intake irritates their mouth or upper gastrointestinal tract.
Severe diarrhea develops in association with dehydration, weight loss, lymphoid tissue involution (see Table 1 for extensive description of PEMS), nearly 100% morbidity in the flock, and very high rates of mortality. Mortality associated with PEMS is characteristic and develops within four days after birds are exposed to the agent(s) that cause the disease (Figure 1).
Qureshi et al. (1997) have demonstrated a severe immunodysfunction in PEMS-infected poults that is characterized by depressed humoral and cell-mediated responses to various antigenic challenges.
In field cases of PEMS, mortality may range between 25 and 96% demonstrating the severely immunocompromised condition of the poults with the disease. Furthermore, survivors are stunted by 40% or more and never attain historical market weights at the 18 to 20 week market age.
Table 1. Macroscopic and histopathological signs in poults naturally and experimentally infected with PEMS.



+ Low frequency of occurrence; ++, +++ Moderate frequency; ++++ High frequency.
The PEMS problem is complicated by the fact that no etiological agent(s) has been identified (Barnes, 1997; Barnes and Guy, 1995; Barnes et al., 1996; Brown, 1995; Qureshi et al., 1997).
Numerous potential viral agents have been investigated including adenovirus, turkey corona virus (TCV), TCV-like particles, enterovirus, astrovirus, birnavirus (serotype 2), rotavirus (type D), reovirus, bursa epithelial virus, and others.
However, alone, these viruses have not been shown to induce PEMS (Barnes and Guy, 1995; Barnes et al., 1996; Brown, 1995). Nevertheless, Brown (1995) reported that a TCV-like particle and a serotype 2 birnavirus could both reduce growth in 3-week-old poults. Co-challenge with these two agents depressed growth and feed conversion ratio and caused 60% mortality. These observations were similar to the condition found in field cases of PEMS.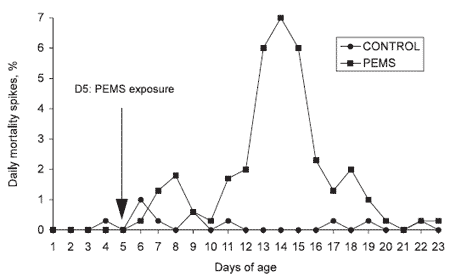
Figure 1. Mortality profile of turkey poults suffering from PEMS.
It also has been observed that cryptosporidiosis and hexamitiasis complicate the PEMS problem on some farms, but neither Cryptosporidium spp., Hexamita, nor any other protozoan infection (such as trichomonads, unidentified flagellates, or Coccidia) alone causes PEMS (Barnes and Guy, 1995; Barnes et al., 1996; Brown, 1995).
On the other hand, Barnes et al. (1996) suggest that after an unidentified virus infects the turkey poults, opportunistic enteric bacteria (Salmonella, E. coli, Clostridium, etc.) further complicate the PEMS condition and cause mortality in the immunocompromised poult.
PEMS: A COMPLEX OF ENTERIC DISEASES?
Over the past seven years, attempts to characterize the PEMS condition have been curtailed since it is not known if PEMS is a single disease or a complex of several diseases. Barnes (1997) has indicated that there are two primary clinical forms of PEMS that are recognized – the milder form called excess mortality (EMT) and the more severe form called spiking mortality of turkey poults (SMT).
When flock mortality equals or exceeds 1% for three or more consecutive days or total mortality exceeds 9% for a three week period, it is believed that the more severe SMT form of PEMS has been evident. When EMT is experienced, mortality does not exceed more than 1% per day for three or more consecutive days and total mortality ranges from 2 to 9% for a three week period.
The PEMS condition is complicated by the fact that TCV is commonly associated with the disease. Nevertheless, Barnes et al. (1997) indicated that there have been field outbreaks of both PEMS-TCV positive and PEMSTCV negative. Furthermore, PEMS has been reproduced experimentally in TCV negative poults. This is especially vexing since TCV appeared to be a leading candidate for the causative etiology for PEMS.
These observations suggest that there may yet be an unidentified virus that is the etiological agent that initiates PEMS. Indeed, researchers at Ohio State University and at Cornell University, working closely with poultry scientists at North Carolina State University, have independently isolated a small virus that is less than 50 nm in diameter that may be the unknown virus that has eluded efforts of many virologists over the past seven years. At this time this virus has not been classified, but preliminary studies indicated that it is pathogenic and can induce a severe diarrhea in young specific-pathogen free turkey poults.
Although research on PEMS has been very slow to yield insight that could lead to a solution to the problem, we have isolated consistently from moribund, PEMS-infected poults two strains of a typical E. coli along with several other E. coli strains.
Although the isolation of these strains was fortuitous, their overwhelming presence in nearly every set of PEMS-infected poults examined by our laboratory from 1995 to 1997 suggested that these E. coli isolates may be very important. Indeed, these E. coli isolates may represent a significant breakthrough in the diagnosis and control of PEMS. Edens et al. (1997a,b,c) believe that atypical E. coli are responsible for the secondary infections and that the E. coli have the potential to be primary pathogens as well.
These bacterial isolates do not appear to be very different from most other E. coli isolates, but several characteristics set them apart from normal enteric bacteria. First, they are highly resistant to most if not all antibiotics commonly used by the turkey industry. However, these isolates were shown to be sensitive to the new fluoroquinalone antibiotics sarafloxacin and enrofloxacin.
The first time that sarafloxacin was used to treat a flock with PEMS, mortality was brought to a complete and abrupt halt, and both of these antibiotics have been used to control newly diagnosed PEMS outbreaks. These observations clearly indicated that there was a significant bacterial component involved in PEMS infections.
However, the E. coli isolates that are commonly found in association with PEMS also develop very rapidly a resistance to the fluoroquinalone antibiotics. In the process of becoming resistant to the fluoroquinalones, the bacteria also change their characteristic biochemical profiles (Edens et al., 1997b).
These E. coli isolates are very aggressive and can penetrate the intestinal epithelium rapidly to become septic within only a few hours after oral administration of as few as 1,000 cells. They are resistant to low levels of formaldehyde often used in disinfection of brooder barns and they are resistant to several other disinfectants commonly used in poultry barns.
PEMS TRANSMISSION
Infection leading to PEMS development is mediated via oral-fecal means and causes frank signs within 24 hours after exposure. Transmission of PEMS agent(s) appears to require physical contact with the poults. Aerosol transmission is difficult, but it is believed that flies and darkling beetles play a major role in the vectoring of the disease agent(s). PEMS is not transmitted vertically through the egg (Edens, 1997), but poults from dams previously exposed to PEMS appear to be slightly less susceptible to the disease.
It was determined early in the emergence of PEMS that once a farm had broken with the disease, it was nearly impossible to prevent its reoccurrence even with extraordinary cleaning and disinfection between flocks. However, we have determined that the E. coli associated with PEMS can be harbored in soil and in well water supplying the brooder barns. Even with rigorous clean-up procedures, it is possible to track the bacteria back into the brooder barns and place new poults at risk for development of a bacterial gastroenteritis.
These observations also explain partially how the disease could be mechanically vectored among widely separated farms. Service personnel and their vehicles may have served as the original vectors. Therefore, very tight biosecurity programs now in place in North Carolina have been successful in reducing the incidence of the disease. Furthermore there has been a concerted effort by several companies to convert farms from multi-age to single-age farms to break the cycle of the disease developing in older birds and transferring to younger birds. Some companies have even gone to off-site cage brooding to prevent development of the disease.
PEMS CAUSES MALABSORPTION ANDWASTING
We have developed an extensive profile of the physiological and biochemical dysfunctions associated with PEMS (Table 2). Based on the data collected from exhaustive studies, it became apparent that PEMS induced a severe malabsorption and metabolic deficiency problem.
During the development of the disease, many poults can be found to have experienced severe wasting of body musculature which in humans could be compared with chronic stress or even diabetes mellitus. Nevertheless, the wasting of the musculature and loss of nearly all of the adipose tissues suggests that even though the poults are eating some feed, nutrient intake is not sufficient to meet body requirements for maintenance and growth. Therefore, it was possible that malabsorption and metabolic dysfunction could be contributing to the disease.
Malabsorption is strongly indicated by pathology associated with the lumenal surface of the intestinal tract. Epithelial cells in the gastrointestinal tract appear to be a target of the undefined etiological agent(s) that cause the disease. Poults with acute gastroenteritis show marked villus atrophy, and often times villi will fuse after an initial contraction. This gives the villus a pleated, rounded appearance.
The enterocytes associated with the jejunum and ileum in PEMS infected poults show significant microscopic and ultrastructural damage. The enterocytes will show characteristic swelling, become pale due to loss of cellular integrity and depletion of cellular components in their structure, and will become distinctly granular due to mitochondrial hypertrophy and disintegration (often times due to invasive E. coli).
The microvilli and terminal web of the enterocytes become damaged. Tight junctions between the epithelial cells will be violated by invasive bacteria and by eosinophilic granulocytes migrating from the lamina propria through the epithelial cells into the intestinal lumen. Cytoplasmic membranes within the enterocytes will also become damaged, fused, and often times disappear.
Perhaps the most significant lesion is associated with the mitochondria which are edematous and swollen to a size that is approximately three to four times as large as those in the control epithelial cells. These lesions signal a potential problem in absorption of nutrient and transport across the cells.
Table 2. Physiological and biochemical changes associated with PEMS.
Malabsorption is also indicated by microscopic changes in the pancreas.
Early in the infection due to PEMS, pancreatic acinar cells become engorged with zymogen granules. These zymogen granules contain digestive enzymes that are released into the pancreatic ducts for delivery to the duodenum where enzymes catalyze digestion of specific nutrients.
Later, during the course of the PEMS infection, the acinar cells loose their bipolar appearance and take on a distinctly basophilic appearance that signals the development of autolysis of the cell. This change in appearance is accompanied by localized infiltration of lymphocytes and often times eosinophils and heterophils. The pancreas shows signs of atrophy based on significant depressions in actual and relative weight.
The violation of the tight junctions between enterocytes also suggests that a good part of the diarrhea associated with this disease can be attributed to paracellular as well as transcellular exudation of body fluids into the gastrointestinal tract. This loss of body fluid is promoted by degenerative changes in the villi, which show apical necrosis, sloughing of apical enterocytes and exudation of protein-laden fluids into the lumen of the intestine.
In field cases of PEMS, hypoglycemia and hypophosphatemia have been suspected as contributors to the disease. The provision to PEMS-afflicted poults of sugar and phosphate in drinking water showed that the diseased poults were not able to absorb adequate amounts of either glucose or phosphate (Table 3) from the intestinal tract.
Even more vital, it appeared that PEMS-infected poults had an impaired capability for utilization of glucose from the circulatory system. This assessment was based upon the inability of PEMSinfected poults to synthesize hepatic glycogen (Table 3) or to down regulate hepatic glucose-6-phosphatase (Table 3), an enzyme that is pivotal in the metabolism of glucose for energy and for glycogen synthesis. These observations confirmed both a severe malabsorption condition and a serious metabolic dysfunction, as well, in PEMS-afflicted poults.
Table 3. Influence of drinking water sugar and phosphate supplementation on selected chemical measurements in poults suffering from PEMS.
a,b,c In a row, means with unlike superscripts differ significantly (P£ 0.05).
Inspection of the data presented in Table 3 suggested that part of the metabolic dysfunction caused by PEMS infection was associated with an apparent inability of the poult to absorb and utilize glucose. This conclusion was based on the observation (1) of continued hypoglycemia after water supplementation of sugar, and (2) that neither hepatic glucose-6-phosphatase decreased nor hepatic glycogen increased in poults that survived PEMS and had water-supplemented sugar available for immediate utilization.
Based upon other data (Table 2), it was concluded that a great deal of the problem associated with an apparent metabolic dysfunction was the inability of the liver to utilize blood glucose, and this was attributed to greatly decreased levels of many metabolically active hormones.
Nutritional interventions for PEMS
After analyzing the profiles for mortality, feed and water consumption, blood and hepatic chemical data, and macroscopic and microscopic lesions associated with PEMS, it became very apparent that there had to be research efforts to determine if there were nutritional interventions that could be implemented to ameliorate the devastating effects of PEMS.
Taking into account the highly significant involvement of pathogenic E. coli in PEMS, we first decided to attempt to control its role by possible adsorption with Bio-Mos (mannanoligosaccharide derived from the cell wall of yeast; Alltech, Inc.), which has been demonstrated to be highly effective in the control of intestinal Enterobacteriaceae including certain E. coli and Salmonella.
Preliminary studies had demonstrated that certain organic acids had some positive effects in ameliorating the effects of PEMS. Therefore, we incorporated into our experimental design the use of Mold-Zap (Alltech, Inc.), which contains buffered propionic acid as one of its active components.
Additionally, our data indicated that there was a metabolic problem in which the poults did not appear to be capable of utilizing blood glucose in energy metabolism (Edens et al., 1997c). It has been demonstrated that chromium (Cr) deficiency caused dysfunctional carbohydrate and protein metabolism, reduced insulin sensitivity in peripheral tissues, impaired growth rate, and Cr deficient diets produced animals that were very susceptible to many different types of stressors (Anderson, 1994; Doisy, 1978; Lindeman, 1996; Mowat, 1994; Mowat et al., 1995; Pagan et al., 1995).
Therefore, in a second experiment, we incorporated BioChrome (chromium yeast, Alltech Inc.) into the diet of control and PEMS-exposed turkey poults.
BIO-MOS AND MOLD-ZAP INFLUENCE ON PEMS
Methods
In these studies, Bio-Mos was incorporated at 0.91 kg/ton, and Mold-Zap was incorporated into the diet at 3.64 kg/ton feed. The diets were provided to the turkey poults from the day of hatching and placement into heated brooding batteries. In each of two trials, five replicate pens with 15 poults per pen in each of eight treatments (Control, Bio-Mos, Mold-Zap, Bio-Mos + Mold- Zap, PEMS, PEMS + Bio-Mos, PEMS + Mold-Zap, and PEMS + Bio-Mos + Mold-Zap) were used.
At five days after hatching, the poults in the PEMSexposed groups were given a 0.1 ml oral gavage of a 10% suspension of fecal material derived from PEMS-infected poults. This procedure has been documented to induce in experimental conditions a disease state that is equal in every detail to field cases of PEMS (Edens et al., 1997a,b,c).
A daily determination of mortality due to PEMS was made, and body weights and certain organ weights (thymus and bursa of Fabricius) were determined at specific intervals through 21 days of age when the experiments were terminated. Cecal contents were sampled periodically, and a determination of bacterial loads was made for birds in the eight different treatments.
Results
Control poults given the Bio-Mos + Mold-Zap combination showed an improvement in body weight in comparison to all other treatment groups (Table 4).
Additionally, mortality in this non-infected group was also the lowest among all other treatment groups. When other parameters (relative weights of thymus and bursa of Fabricius and cecal bacterial load) were considered, it was evident that the combination of Bio-Mos and Mold-Zap had a significant effect on the poults.
The improvement in body weight may be attributed partially to the significant reduction in cecal E. coli loads, and this may be reflected in the largest relative weight of the thymus found among all treatment groups (Table 4). However, when Mold-Zap without Bio-Mos was provided in the feed, there was a significant reduction in three-week body weight in comparison to the non-infected Control group. This observation may be due to a significant reduction in total bacterial count in the cecum and to a five log decrease in E. coli counts (Table 4).
In the PEMS-infected groups, equivocal responses to Bio-Mos, Mold-Zap and their combination were found (Table 4). There was a strong tendency for Bio-Mos and Mold-Zap alone to reduce PEMS-associated mortality (Table 4).
In fact, mortality was delayed in its appearance in these treatment groups. While the combination of Bio-Mos and Mold-Zap did not reduce PEMS-associated mortality (Table 4), the body weights of the poults in this PEMS-infected group were significantly larger than weights of the unsupplemented PEMS-infected group and the PEMS/Mold-Zap group.
Body weights of the PEMS/Bio-Mos treatment group did not differ from the combination PEMS/Bio-Mos and Mold- Zap group. When the relative weights of the thymus of poults infected with PEMS were compared, poults given Mold-Zap alone had the lowest thymus relative weight followed by the PEMS-alone groups.
The thymus weights for the infected poults given Bio-Mos and those given the combination Bio-Mos and Mold-Zap were the largest among all of the PEMS-infected groups. This coincidentally corresponded to the largest body weights among the four infected treatment groups. Among the PEMS-infected treatment groups, there was no apparent relationship between cecal bacterial and cecal E. coli counts and performance characteristics of the poults in these groups (Table 4).
Bio-Mos has been shown to improve significantly the body weight, livability, condemnation rate, and feed conversion of growing turkeys (Olsen, 1995). Savage and Zakrzewska (1996) provided evidence of Bio-Mosimproved performance in brooding turkey poults when they demonstrated improved weight gain, livability and feed conversion in the poults through four and eight weeks of age.
Bradley et al. (1994) fed turkey poults a diet that contained Saccharomyces cerevisiae and found a morphological change in the villi of the ileum in the intestinal tract. Bradley et al. (1994) found reduced numbers of goblet cells per villus and decreased depth of the crypts of Lieberkuhn, which are indicative of reduced bacterial load, in the gut of Bio-Mos fed poults.
Spring (1996, 1997) indicated that mannanoligosaccharides in broilers increased intestinal villus length allowing development of a greater absorptive surface area. However, Spring (1996,1997) also noted increased crypt depth in the Bio-Mos fed broilers, indicating a greater energy expenditure for production of the greater absorptive surface area. Edens and Doerfler (1997b) reported a significant increase in the number of goblet cells and shortened villi in PEMS-infected poults.
Table 4. Influence of Bio-Mos and Mold-Zap on the performance of PEMS-infected turkey poults.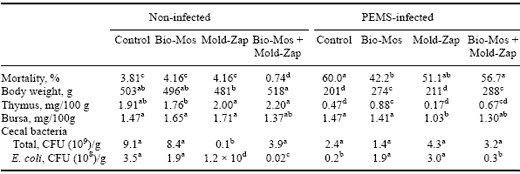
In a row, means with unlike superscripts differ significantly (P£ 0.05).
Savage and Zakrzewska (1996) showed that Bio-Mos fed turkey poults had higher secretory IgA levels in bile and significantly elevated IgG levels in blood, indicating an enhanced immune response that would have been associated with the gut mucosa and humoral immunity as well.
In the gut, an enhanced immune response would be associated with better exclusion of potentially pathogenic microorganisms and heightened levels of immunoglobulins in the blood would be associated with enhanced ability to recognize and combat systemic infections.
Cotter (1997) explored the influence of feeding Bio- Mos on the development of the cell-mediated immune response in female chickens. Using the most basic of tests for modulation of cellular immunity, the phytohemagglutinin (PHA) wattle response, he documented a modulatory role of Bio-Mos in primary, secondary and tertiary kinetic reactions when comparison of the experimental results were made with control responses.
Bio-Mos fed birds did not permit unrestricted swelling in secondary and tertiary, but in fact, Bio-Mos fed birds had reduced swelling of the wattle. This reduced PHAwattle response indicated that Bio-Mos fed birds had developed an immunological memory that enhanced the removal of the antigen more quickly in the sensitized bird thereby accelerating the return of tissue homeostasis.
Ostensibly, these improvements in poultry performance were due, in part, to the non-specific immunoenhancing properties of Bio-Mos (Newman, 1994, 1995). Bio-Mos is capable of stimulating phagocytic and macrophage activity thereby increasing immune surveillance of the first-line immunological defense mechanism in the rat and in mitogen-stimulated monocytes (Newman, 1994, 1995). Newman (1994) provided evidence of the pathogen-binding action of Bio-Mos showing that the mannanoligosaccharide in Bio-Mos competed for binding sites on the intestinal epithelial cells preventing certain bacterial species from binding and causing pathology.
Broiler chickens, market turkeys, pigs, and calves all showed improved performance when Bio-Mos was provided in their feed (Newman, 1994). In broiler chickens, Bio-Mos improved livability and feed conversion in broilers grown in houses with a history of poor performance (Macdonald, 1995).
Stewart (1995) also demonstrated a significant effect of Bio-Mos in broiler chickens when he reported significantly improved resistance of broiler chickens to Salmonella challenge and to aflatoxin in the feed. These pathogen-binding and detoxification properties of Bio-Mos were attributed to the mannanoligosaccharides associated with the strain of Saccharomyces cerevisiae used to make the product. Killeen and Rosell (1996) reviewed the scientific literature dealing with mechanism of action of mannanoligosaccharides to bind bacteria in several animal species and concluded that the use of mannans in combination with pathogen-associated antigens could induce protective cellular immune responses.
Doerfler et al. (1997) reported that the use of organic acids in diets of PEMSexposed turkey poults resulted in improved livability. However, continuous use of an organic acid-based (propionic acid) product at high levels caused 21-day body weight of female poults to be reduced but not the weights of male poults. Feed conversions were improved by low level feeding of the organic acids.
However, body weight reduction due to PEMS infection was not alleviated by the organic acid product similar to the response seen in this study with Mold-Zap. However, both Mold-Zap in this study, and the propionic acid product in the previous study clearly decreased cecal bacterial loads in control poults; and in the previous study bacterial loads were reduced significantly in PEMS-infected poults as well.
In the previous study, reduced body weight in the organic acid-fed birds was attributed to decreased intestinal microflora populations similar to this study with Mold-Zap. In both studies, there was reduced mortality when the organic acids were incorporated into the diet of PEMS-infected and in non-infected control poults.
Summary: Bio-Mos and Mold-Zap
These data show that the use of Bio-Mos and Mold-Zap in the feed of newly hatched non-infected turkey poults has some significant benefits. In the appropriate level and combination, there are pathogen-reducing effects that result in improved growth, livability and thymus status that may be indicative of improved immunological status. This also extended to the poults that were PEMS-infected.
The use of Bio-Mos and Mold-Zap alone and in combination in the PEMS-infected poults also resulted in larger poults and improved livability. The improved body weights noted with the combination of Bio-Mos with Mold-Zap probably suggest an improved absorption of nutrients in the PEMS-infected and in control poults.
Edens and Doerfler (1997a) and Edens et al. (1997a) have shown that PEMSinfected poults have shortened intestinal villi and reduced functional microvilli on enterocytes. Improvement in either area (villus height or in enterocyte functional microvilli numbers) would improve nutrient absorption. Additionally, the known immunodysfunction associated with PEMS infection may have been modulated by both Bio-Mos and Mold-Zap.
Certainly cecal bacterial load was affected by Mold-Zap and to a limited extent by Bio-Mos as well. These reductions in bacterial load may indicate an improved gut-associated immune status in the PEMS-infected and control poults alike, but further study is required in this area. Although these products were not the panacea that turkey producers are seeking, they do have an apparent value as interventions against PEMS development and in promotion of better performance in non-infected poults.
BIOCHROME (CHROMIUM YEAST) INFLUENCE ON PEMS
Methods
In these studies, BioChrome, an organic Cr produced by yeast, was incorporated into the diet at the level of 400 ppb Cr (364 mg/ton) of feed. The diet was provided from the day of hatching and placement into heated brooding batteries.
In each of two trials, eight replicate pens with 15 poults per pen in each of four treatments (Control, BioChrome, PEMS, and PEMS + BioChrome) were used. At five days after hatching, the poults in the PEMS-exposed groups were given a 0.1 ml oral gavage of a 10% suspension of fecal material derived from PEMS-infected poults. A daily determination of mortality due to PEMS was made, and body weights and blood samples were collected at specified intervals through 21 days of age when the experiments were terminated. Blood samples were analyzed for a series of metabolically active hormones and other blood chemistries.
Results
BioChrome added to diets of non-infected Control and PEMS-infected poults did not affect mortality rates. As expected, PEMS mortality rates were elevated significantly (Table 5). In non-infected Control poults, BioChrome did not alter body weight, but in PEMS-infected poults, BioChrome caused a small but significant increase in body weight (Table 5).
In previous studies, PEMS infection was associated with significantly decreased serum levels of insulin, thyroxine, triiodothyronine, and an elevation in glucagon (Edens and Doerfler, 1997a; Doerfler et al., 1998). In this study PEMS significantly decreased insulin, thyroxine, and triiodothyronine but elevated glucagon (Table 5). BioChrome added to diets of PEMS-infected poults significantly elevated serum glucagon, thyroxine, and triiodothyronine (Table 5).
Table 5. Influence of BioChrome on the performance of PEMS-infected turkey poults.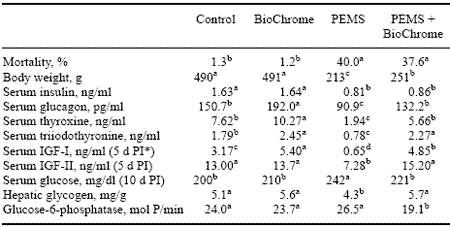
a,b,c,d In a row, means with unlike superscripts differ significantly (P£ 0.05).
*PI = post-infection.
Serum insulin was unchanged in both Control and PEMS-infected poults (Table 5). In this study, we also measured serum IGF-1 and IGF-2. Generally, BioChrome added to Control poult diets resulted in a significant elevation in serum IGF-1 but not IGF-2 (Table 5). In the PEMS-infected poults (Table 5), BioChrome caused both serum IGF-1 and IGF-2 to be elevated significantly to a level similar to non-infected Controls. PEMS infection without supplemental BioChrome caused both IGF-1 and IGF-2 to be depressed significantly (Table 5).
Serum glucose was elevated in PEMS-exposed poults that had survived the elevated mortality period of the disease (Table 5). Generally, serum glucose was unaffected by BioChrome (Table 5). However, an interesting BioChrome effect was found in the hepatic glycogen response to PEMS.
In earlier studies we reported a significant depression in hepatic glycogen in PEMS-infected poults (Doerfler et al., 1998), and in this study a similar response was seen.
However, in the BioChrome-supplemented PEMS-infected poults, hepatic glycogen levels were comparable to levels found in non-infected Controls. Hepatic glucose-6- phosphatase activity in PEMS survivor poults was near the levels found in noninfected Control poults (Table 5); but in PEMS-infected poults, BioChrome significantly decreased glucose-6-phosphatase activity. The decrease in hepatic glucose-6-phosphatase activity would be expected if glucose uptake was induced by elevated blood glucose or by stimulation of insulin receptors that should facilitate glucose transport into the liver.
Summary: Biochrome and PEMS
Although BioChrome did not reduce mortality associated with PEMS, there was a small but significant increase in body weight in the infected poults given the Cr supplement. This increase in body weight appears to be related to altered levels of metabolically active serum hormones. The hormones of primary interest in this study were the thyroid hormones and insulin.
In a previous study (Doerfler et al., 1998), we documented severe hypothermia, hypothyrosis and hypoglycemia in PEMS-infected poults. Additionally, Edens and Doerfler (1997a) and Doerfler et al. (1998) reported that there appeared to be an inability in PEMS-infected poults to absorb glucose from the intestine and to utilize blood glucose at the level of the liver.
These problems were attributed to the decreased level of metabolically active hormones in the circulatory system. In this scenario, insulin plays a major role. Insulin is required for entry of blood glucose into the liver and for stimulation of the biochemical pathways of glycolysis in the TCA cycle leading to energy metabolism. In the liver, under the influence of insulin, glucose is converted to lipids in the form of fatty acids, but in PEMS this does not appear to occur normally.
Insulin inhibits proteolysis that is evident in the PEMS condition; and it enhances protein and amino acid sequestration in all target tissues. In this mode, insulin is a potent anabolic hormone. Insulin also contributes to body growth by stimulating the synthesis of macromolecules in such tissues as cartilage and bone, further contributing to the growth of animals.
The reduced level of serum insulin noted in this study in PEMS-infected poults can explain the wasting and reduced growth of infected poults. However, BioChrome supplementation of infected poults did not elevate serum insulin, but body weight did show a small but significant increase. In addition, hepatic glycogen was elevated and hepatic glucose-6-phosphatase activity decreased in the PEMS-infected poults given BioChrome. This indicated that they had likely responded to BioChrome by becoming more sensitive to the lower levels of available serum insulin.
Insulin also regulates the transcription of related gene growth factors such as insulin-like growth factors 1 and 2. These factors are also called somatomedins. Fasting, protein deprivation, and insulin deficiency all lead to diminished hepatic synthesis and secretion of the somatomedins.
In PEMS infected poults, significant depressions in serum levels of both IGF-1 and IGF-2 were found, but BioChrome supplementation increased levels to those similar to non-infected poults to non-infected levels. These observations also support the idea that BioChrome in the PEMS-infected poults had sensitized their insulin receptors permitting the infected poults to develop a metabolic condition that could better sustain extreme stress.
Thyroid hormones have been reported to decrease significantly in PEMSinfected poults (Edens and Doerfler, 1997a,b; Doerfler et al., 1998), and in this study both triiodothyronine and thyroxine were decreased significantly by the infection. Nevertheless, in PEMS-infected poults given BioChrome, both of these serum hormones were elevated to normal, non-infected levels.
The most obvious effects of thyroid hormones are to increase the rate of oxygen utilization and basal heat production. The decrease in body temperature in PEMS, therefore, can be related directly to the hypothyrosis that normally occurs with this disease.
In this study, the increase in serum thyroid hormones in response to BioChrome supplementation apparently resulted in a metabolic energy demand. Even though the PEMS-infected poults responded positively to BioChrome supplementation, body weights were not corrected in the time frame of the study. Additionally, feed consumption during the peak of the PEMS infection was reduced to approximately 25% of the Control feed intake, and this would further reduce the ameliorating effects of BioChrome.
The effects of BioChrome in poultry species are not well documented, but it appears that poultry respond in a manner similar to mammals. Specifically, BioChrome appears to exert its effects through sensitization of insulin receptors and through its influence on glucose metabolism. This mediation of glucose metabolism in PEMS-infected poults appears to be through stimulation of the thyroid hormones. Furthermore, Cr in mammals is recognized as an anti-stress agent (Mowat, 1994; Pagan et al., 1995; Mallard and Borgs, 1997), and it is evident that PEMS infection in turkeys is extremely stressful.
| Conclusions Nutritional interventions into the PEMS problem appear to have a potential that is even greater than the use of antibiotics. This is especially important because many of the bacterial pathogens isolated from PEMS-infected poults are antibiotic resistant or develop resistance very rapidly. It has been our position that the pathogens that cause PEMS induce pathology via disruption of the normal biochemical and physiological mechanisms that establish the individual bird’s homeostatic condition. Disruption of any element in the normal bird will contribute to its eventual demise. Since the lack of assimilation of nutrients and ability to utilize the nutrients once absorbed from the intestinal tract are primary contributors to the mortality and lack of growth in PEMS afflicted poults, it is reasonable to think that early nutritional prophylaxis would be beneficial to the birds. In fact, the data collected in these initial studies strongly support the use of dietary supplements that will control intestinal pathogens, modulate the immune system and provide a stimulus to the metabolic state of PEMS-afflicted poults. |
References
Anderson, R.A. 1994. Stress effects on chromium nutrition of humans and farm animals. In: Biotechnology in the Feed Industry. Proc. Alltech’s 10th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 267–274.
Barnes, H.J. 1997. Prevention, control, and treatment of poult enteritismortality syndrome (spiking mortality of turkeys). In: Proc. Pfizer Animal Health Conference, National Turkey Federation Annual Convention, San Francisco, CA, January 13, 1997. Pfizer Animal Health Group, Pfizer, Inc., New York, NY 10017–5755. pp. 9–13.
Barnes, H.J. and J.S. Guy. 1995. Spiking mortality of turkeys (SMT) and related disorders: an update. Proc. 19th Annual North Carolina Turkey Industry Days Conference. NCSU, Raleigh, NC., pp. 16-21.
Barnes, H.J., J.S. Guy, T.P. Brow and F.W. Edens. 1996. Poult enteritis and mortality syndrome (“spiking mortality in turkeys”) and related disorders: An update. College of Veterinary Medicine, North Carolina State University, Raleigh, NC 27695. October 6, 1996. pp. 1–11.
Barnes, H.J., J.S. Guy, J.T. Weaver and S.R. Jennings. 1997. Turkey flocks with high spiking mortality that are negative for turkey coronavirus. Proc. Amer. Vet. Med. Assoc. Conv. (Reno, NV) 134: 168.
Bradley, G.L., T.F. Savage and K.I. Timm. 1994. The effect of supplementing diets with Saccharomyces cerevisiae var. boulardii on male poult performance and ileal morphology. Poultry Sci. 73:1766–1770.
Brown, T.P. 1995. Spiking mortality: pathology, performance, and prevention. Proc. 6th Annual Eli Lilly Technical Seminar. Indianapolis, IN. 6:34-44.
Cotter, P.F. 1997. Modulation of the immune response: Current perceptions and future prospects with an example from poultry. In: Biotechnology in the Feed Industry. Proc. Alltech’s 13th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 195–203.
Doerfler, R.E., F.W. Edens, M.A. Qureshi, C.R. Parkhurst and G.B. Havenstein. 1997. Influence of Mycocurb on performance of turkey poults with experimentally-induced poult enteritis and mortality syndrome. Poultry Sci. 76(Suppl. 1):124 (Abstr).
Doerfler, R.E., F.W. Edens, C.R. Parkhurst, G.B. Havenstein and M.A. Qureshi. 1998. Hypothermia, hypoglycemia, and hypothyrosis associated with poult enteritis and mortality syndrome. Poultry Sci. 77(in press).
Doisy, R.J. 1978. Effect of nutrient deficiencies in animals: Chromium. In: CRC Handbook Series in Nutrition and Food. Section E: Nutritional Disorders. Vol. II Effect of Nutrient Deficiencies in Animals. (M. Rechcigl, Jr ed.) CRC Press, Inc. West Palm Beach, FL. pp. 341–342.
Edens, F.W. 1997. Researchers battle disease that affects young poults. Cuddy Farms – The Feather File, Autumn 1997. pp. 1–2.
Edens, F.W. and R.E. Doerfler. 1997a. Glucose in metabolism in poult enteritis and mortality syndrome. In: Proc. 20th Technical Turkey Conference, April 16–17, 1997. Pott Shrigley, Near Macclesfield, Cheshire, England. pp. 106–119.
Edens, F.W. and R.E. Doerfler. 1997b. Cellular and biochemical lesions associated with poult enteritis and mortality syndrome. Proc. Amer. Vet. Med. Assoc. Conv. (Reno, NV) 134:169.
Edens, F.W., C.R. Parkhurst, M.A. Qureshi, I.A. Casas and G.B. Havenstein. 1997a. Atypical Escherichia coli strains and their association with poult enteritis and mortality syndrome. Poultry Sci. 76: 952–960.
Edens, F.W., R.A. Qureshi, C.R. Parkhurst, M.A. Qureshi, G.B. Havenstein and I.A. Casas. 1997b. Characterization of two Escherichia coli isolates associated with poult enteritis and mortality syndrome. Poultry Sci. 76:1665–1673.
Edens, F.W., M.A. Qureshi, S.E. Mann, C.R. Parkhurst and G.B. Havenstein. 1997c. The involvement of eosinophils in the pathogenesis of poult enteritis and mortality syndrome. Poultry Sci. 76(Suppl. 1):111.
Edens, F.W., K.A. Joyce, C.R. Parkhurst, G.B. Havenstein and M.A. Qureshi. 1998. Effect of litter moisture and brooding temperature on growth and livability of turkey poults experiencing poult enteritis and mortality syndrome. Poultry Sci. 77: (in press).
Killeen, G. and V. Rosell. 1996. The potential of polysaccharide supplements in diets for livestock and pets. In: Biotechnology in the Feed Industry. Proc. Alltech’s 12th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 149–158.
Lindeman, M.D., 1996. Organic chromium – The missing link in farm animal nutrition. In: Biotechnology in the Feed Industry. Proc. Alltech’s 12th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 299–314.
Macdonald, F. 1995. Use of immunostimulants in agricultural applications. In: Biotechnology in the Feed Industry. Proc. Alltech’s 11th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 97–103.
Mallard, B.A. and P. Borgs. 1997. Effects of supplemental trivalent chromium on hormone and immune responses of cattle. In: Biotechnology in the Feed Industry. Proc. Alltech’s 13th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 241–250.
Mowat, D.N. 1994. Organic chromium:Anew nutrient for stressed animals. In: Biotechnology in the Feed Industry. Proc. Alltech’s 10th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp 275–282.
Mowat, D.N., A. Subiyatno and W.Z. Wang. 1995. Chromium deficiency in first parity cows. In: Biotechnology in the Feed Industry. Proc. Alltech’s 11th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 309–314.
Newman, K.E. 1994. Mannan-oligosaccharides: natural polymers with significant impact on the gastrointestinal microflora and the immune system. In: Biotechnology in the Feed Industry. Proc. Alltech’s 10th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 167–174.
Newman, K.E. 1995. The immune system: Nature’s defense mechanism – Manipulating it through nutrition. In: Biotechnology in the Feed Industry. Proc. Alltech’s 11th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 77–86.
Olsen, R. 1995. Mannanoligosaccharides: Experience in commercial turkey production. In: Biotechnology in the Feed Industry. Proc. Alltech’s 11th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 389–392.
Pagan, J.D., S.G. Jackson and S.E. Duren. 1995. The effect of chromium supplementation on metabolic response to exercise in throughbred horses. In: Biotechnology in the Feed Industry. Proc. Alltech’s 11th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 249–256.
Qureshi, M.A., F.W. Edens and G.B. Havenstein. 1997. Immune system dysfunction during exposure to poult enteritis and mortality syndrome. Poultry Sci. 76: 564–569.
Savage, T.F. and E.I. Zakrzewska. 1996. The performance of male turkeys fed a starter diet containing a mannan-oligosaccharide (Bio-Mos) from day old to eight weeks of age. In: Biotechnology in the Feed Industry. Proc. Alltech’s 12th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 47–54.
Spring, P. 1996. Effects of mannanoligosaccharide on different cecal parameters and on cecal concentrations of enteric pathogens in poultry. Diss. ETH no. 11897. ETH Zurich, Switzerland.
Spring, P. 1997. Understanding the development of the avian gastrointestinal microflora:An essential key for developing competitive exclusion products. In: Biotechnology in the Feed Industry. Proc. Alltech’s 13th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 313–324.
Stewart, G.G. 1995. Non-traditional uses of yeast and its products: the past fifteen years. In: Biotechnology in the Feed Industry. Proc. Alltech’s 11th Ann. Symp. (T.P. Lyons and K.A. Jacques, eds.) Nottingham University Press, Loughborough, Leics, UK. pp. 105–115.







.jpg&w=3840&q=75)