Performance and Molecular Identification of Bacteria Isolated from The Gut of Broiler Birds After Antibiotic Administration and Enzyme Supplementation
This study evaluated the effect of feed additives (antibiotic or enzyme) on performance and bacteria population in the ileum of one day – old broiler chicks (ANAAC 2000) randomly distributed to three treatments having eight replicates and ten birds per replicate. Completely randomized design was used and experiment lasted for 35 days.
Maize–soybean meal diet without antibiotic administration or enzyme supplementation served as control and was the same diet for all treatments. Antibiotic (Dicoxin plus®) was administered to birds fed diet 2 and enzyme (Roxazyme G 2G®) was supplemented to diet 3.Bacterial specific primers for rRNA gene sequences were used to amplify bacterial genes from samples and sequenced. Bacteria were identified from the gene sequences using Basic Local Alignment Search Tool (BLAST) against the National Centre for Biotechnology Information (NCBI).
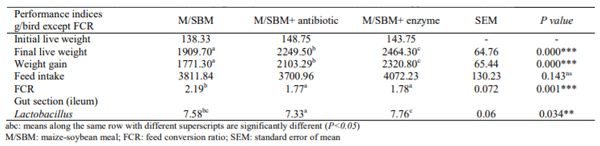
Enzyme supplementation significantly (P<0.01) improved final Live body weight and weight gain compared to control or administration of antibiotic. The FCR was significantly enhanced (P<0.01) by enzyme and antibiotic supplementation. Lactobacillus acidophilus (L. acidophilus), Escherichia coli (E. coli) and Clostridia were identified in digesta sampled. Partial rRNA sequences identical to Clostridia were the lowest (1) in control and enzyme treatment. A value of 4 was recorded in antibiotic treatment. Lactobacillus acidophilus was numerically high in control (8) and enzyme treatment (5) compared to antibiotic administration (1). Partial rRNA sequences identical to Escherichia coli sequences was however high (48) in birds administered antibiotic compared to control (8) and enzyme treatment (9). Results indicated greater improvement in weight gain, FCR and Lactobacillus in broilers fed enzyme supplemented diet. Feed additives may affect the biodiversity of gut bacteria in poultry birds.
Keywords: antibiotics, broilers, enzyme supplementation, molecular identification, performance.
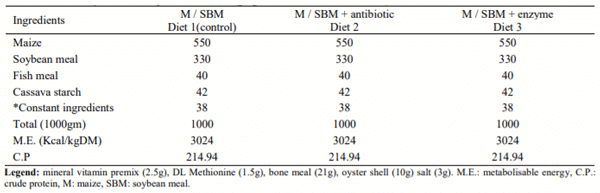

- Endo-1,4-glucanaseactivity: min 8,000 unit per gram (E.C.3.2.1.4.)
- Endo-1,3 (4)-glucanase activity: min 18,000 unit per gram (E.C.3.2.1.6.)
- Endo-1,4-xylanase activity: min 26,000 units per gram (E.C. 3.2.1.8.)

Abule, R. T. S, Obaweki, V. E.,Etebu, E. &Ohimain, E. I. (2014). Effect of Feed addictives on Performance and Lactobacillus Counts in Broilers. 3rd Animal Science Association of Nigeria - Nigerian institute of Animal Science, Joint Annual Meeting (ASAN-NIAS JAM) University of Ibadan, Ibadan – Nigeria. Book of Abstract pg. 81 – 82.
Altschul S. F., Gish W, Miller W, Myers E.W. & Lipman D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215: 403-410. PMID: 2231712[PubMed-indexedfor MEDLINE]
AOAC, (1990). Official methods of analysis, 15th ed. Association of Official Analytical Chemists, Dc.
Apajalathi, J. & Bedford, M.R. (1999). Improved bird performance by feeding its microflora. World Poult. Sci. J. 15:20 – 23.
Apajalati, J., Kettunen, A., & Graham, H. (2004). Characteristics of the gastrointestinal microbial communities with specific reference to the chicken. J. World. Poult. Sci. 60: 223 – 232. http://dx.doi.org/10.1079/WPS200415 Apajalati, J.H.A., Kettunen, A., Bedford, M.R. & Holben, W. (2001). Percent G+C profiling accurately reveals diet related differences in the gastrointestinal microbial community of broiler chickens. Applied Environmental Microbiology, 67:5656 – 5667. http://dx.doi.org/10.1128/AEM.67.12.5656-5667.2001
Bach Knudsen, (1997). Carbohydrate and lignin contents of plant materials used in animal feeding. Animal Feed Science and Technology. 67 (4): 319 – 338.Doi: http://dx.doi.org/10.1016/S0377-8401(97)00009-6
Bao, Y.M., Romero, L.F. & Cowieson, A. J.(2013). Functional patterns of exogenous enzymes in different feed ingredients. Worlds Poult. Sci. J. 69: 759 - 1 http://dx.doi.org/10.1017/S0043933913000792
Barrow, P.A., 1992. Probiotics for chickens, In: R. Fuller (Ed.). Probiotics: The scientific basis, pp255 – 257. (Chapman and Hall, London).
Bedford, M.R. & Cowieson, A. J. (2012).Exogenous enzymes and their effects on intestinal microbiology. Animal Feed Sci. Technol. 173: 76 - 85. http://dx.doi.org/10.1016/j.anifeedsci.2011.12018
Centeno, M.S.J., Ponte, P.I.P., Ribeiro, T., Prates, J.A.M., Ferreira, L.M.A., Soares, M.C., Gilbert, H.J. & Fontes, C.M.G.A. (2006). Galactanases and mannanases improve the nutritive value of maize and soybean meal based diets for broiler chicks. J. Poult. Sci. 43, 344-350. http://doi.org/10.2114/jpsa.43.344 Chesson, A. (2001). Non – starch polysaccharide degrading enzymes in poultry diets: Influence of ingredients on the selection of activities. Worlds. Poult. Sci. J. 57:251 – 263. http://dx.doi.org/10.1079/WPS20010018
Collins, N.E., Moran, E.T. & Stillborn, H.L.(1998). Corn hybrid and bird maturity affect apparent metabolisable energy values. Poult. Sci. Abstract.77:42 – 42.
Courtin C.M., Broekaert W.F., Swennen K, Lescroart O., Onagbesan, O., Buyse, J., Decuypere, E., Van de Wiele, T., Marzorati, M.&Verstraete, W. (2008). Dietary inclusion of wheat bran arabinoxylo oligosaccharides induces beneficial nutritional effects in chickens. Cereal Chemistry, 85: 607 - 613.Doi: http://dx.doi.org/10.1094/CCHEM-85-5-0607
Cowieson, A.J. & Ravindran, V. (2008). Sensitivity of broiler starters to three doses of an enzyme cocktail in a maize-based diet. Br. Poult. Sci. 49: 340 - 346. http://dx.doi.org/10.1080/00071660802126669
Cowieson, A.J. (2005). Factors that affect the nutritional value of maize for broilers. Anim. Feed Sci. Tech. 119:293 305.Doi: http://dx.doi.org/10.1016/j.anifeedsci.2004.12.017
Cowieson, A.J., (2010). Strategic selection of exogenous enzymes for maize/soy-based poultry diets. J. Poult. Sci. 47, 1-7. http://dx.doi.org/10.2141/jpsa.009045 Dibner, J.J. &Buttlin, P. (2002). Use of organic acids as a model to study the impact of gut flora on nutrition and metabolism. J. Appl. Poult. Res. 11: 453 - 1 http://dx.doi.org/10.1093/japr/11.4.453
Dibner, J.J. & Richard, J.D.(2004). Digestive system: Challenges and opportunities. Journ. Of Applied Poult. Res. 13:86 – 93. http://dx.doi.org/10.1093/japr/13.1.86
Dibner, J.J. & Richards, J. D. (2005): Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci 84: 634 - 643. http://dx.doi.org/10.1093/ps/84.4.634
Droleskey, R.E., Oyofo, B.A., Hargis, B.M., Corrier, D.E. & Deloach, J.R.(1994). Effect of mannose on Salmonella typhimurium – mediated loss of mucosal epithelial integrity in cultured chick intestinal segments. Avian Diseases. 38: 275 – 281. http://dx.doi.org/10.2307/1591949
Ezenwa, V., Gerardo, N., Inuoye, D., Medina, M. & Xavier, J. (2012). Animal behavior and the microbiome. Science.338:198 – 199. http://dx.doi.org/10.1126/science.1227412
Falk, P.G., Hooper, L.V., Midtvedt, T. & Gordon, J.I. (1998). Creating and maintaining the gastrointestinal ecosystem: what we need to know from gnotobiology. Microbial Molecular Biology Rev. 62:1157 – 1170. http://dx.doi.org/1092-2172/98/$04.00+0
Gianneanas, I. (2008). How to use plant extracts and phytogenics in animal diets In: The future of Animal production. Binder, E.M., Schatzmayr, G. (editor). Nottingham University Press, Nottingham: 111 - 129.
He, J., Yin, J., Wang, L., Yu, B. & Chen, D. (2010).Functional characterization of a recombinant xylanase from Pichia pastoris and effect of the enzyme on nutrient digestibility in weaned swine. Br. J. Nutr. 103: 1507 - 1513. http://dx.doi.org/10.1017/S0007114509993333
Hetland, H., Choct, M. & Shivus, B. (2004). Role of insoluble non – starch polysaccharides in poultry nutrition. Worlds Poult.Sci.J60:415 – 422. http://dx.doi.org/10.1079/wps200325
Hillman, K, Spencer, R.J., Murdoch, T.A.& Stewart, C.S. (1995).The effect of mixtures of Lactobacillus spp. on the survival of entero toxigenic Escherichia coli in in -vitro continuous culture of porcine intestinal bacteria. Lett. Appl. Microbiol. 20: 130 - 133. http://dx.doi.org/10.1111/j.1471-765x.1995.tb01303.x Hogberg, A. & Lindberg, J.E. (2004). Influence of cereal non - starch polysaccharides and enzyme supplementation on digestion site and gut environment in weaned piglets. Anim. Feed Sci. Technol. 116: 113 - 128. http://dx.doi.org/10.1016/j.anifeedsci.2004.03.010
Hubener, K., Vahjen, W. & Simon, O. (2002). Bacterial responses to different dietary cereal types and xylanase supplementation in the intestine of broiler chickens. Arch. Anim.Nutr. 56: 167 - 187.Doi: PMID:12391903[PubMed-indexed for MEDLINE]
Ikoro, S.G.(2010). Effect of enzyme supplemented maize – soybean meal based diet on gut microflora, pH and performance of broilers. B.Sc. project, department of Animal Science, Niger Delta University.
Kiarie, E., Nyachoti, C.M., Slominski, B.A.& Blank, G. (2007). Growth performance, gastrointestinal microbial activity, and nutrient digestibility in early - weaned swine fed diets containing flaxseed and carbohydrase enzyme. J. Anim. Sci. 85: 2982 - 2993.Doi: PMID:17686904[PubMed-indexed for MEDLINE] Klasing, K.C., Laurin, D. E., Peng, R.K. & Fry, M.(1987). Immunology mediated growth depression in chicks: Influence of feed intake, corticosterone and interleukin – 1. Journal of Nutrition 117:1629 – 1637. http://dx.doi.org/0022-3166/87$3.00
Lan, Y., Verstegen, M.W., Tammings, S. & Williams, B.A., (2005). The role of the commercial gut microbial community in broiler chickens. Worlds Poult. Sci. J. 61: 95 – 104.Doi: http://dx.doi.org/10.1079/wps200445
Leeson, S., Yersin, A. & Volker, L., (1993). Nutritive value of corn crop. J. Appld. Poult. Res. 2: 208 – 213. http://dx.doi.org/10.1093/japr/2.3.208
Martin F.P.& Collen M.C. (1998).Bias in template-to-product ratios in multi template PCR. Appl and Environ. Microbiol. 64(10): 3724 —3730. PMCID:OMC10653
Meng, X. &Slominski, B.A.(2005). Nutritive value of corn, soybean meal, canola meal and peas for broiler chickens as affected by a multi carbohydrase preparation of cell wall degrading enzymes. Poult. Sci. 84:1242 – 1251. http://dx.doi.org/10.1093/ps/84.8.1242
Neish, A.S.(2002).The gut microflora and intestinal epithelial cells. Microbes Infect, 4: 309 – 317. http://dx.doi.org/10.1016/S1286-457(02)01543-5
Noy, Y. & Sklan, D. (1995). Digestion and absorption in the young chick. Poult. Sci. 74:366 – 373. http://dx.doi.org/10.1093/ps/74.366
Ohimain, E.I. & Ofongo, R.T.S. (2013). Effect of enzyme supplemented diet on gut microflora, digesta ph and performance of broiler chickens. Journal of microbiology biotechnology and food sciences3 (2)127-131. http://www.jmbfs.org/wp-content/uploads/2013/09/m...
Olukosi, O.A., Cowieson, A.J. & Adeola, O. (2007). Age-related influence of a cocktail of xylanase, amylase and protease or phytase individually or in combination in broilers. Poult. Sci. 86:77 – 86. http://dx.doi.org/10.1093/ps/86.1.77
Partanen, K.H. & Mroz, Z. (1999). Organic acids for performance enhancement in pig diets. Nurt. Res. Rev. 12: 117 – 145. http://dx.doi.org/10.1079/095442299108728884
Patterson, J.A.& Burkholder, K.M. (2003).Application of prebiotics and probiotics in poultry production. Poult. Sci. 82: 627 - 631. http://dx.doi.org/10.1093/ps/82.4.627
Pirgozliev, V. & Ross, S.P. (1999). Net energy systems for poultry feeds: A quantitative review. Worlds Poult. Sci. J. 55: 23 - 36.Doi: http://dx.doi.org/10.1079/wps19990003
Pluske, J.R., Kim, J.C., McDonald, D.E., Pethick, D.W.& Hampson, D.J.(2001). Non - starch polysaccharides in the diets of young weaned piglets. Pages 81 - 112 in: The Weaner Pig: Nutrition and Management. M. A. Varley and J. Wiseman, ed. CABI Publishing, Wallingford, U.K.
Ravindran, V.(2012). Advances and future directions in poultry nutrition: An overview. Korean J. Poult. Sci,39: (1), 53 - 62. http://dx.doi.org/10.5536/KJPS2012.39.1.053
Ravindran, V.(2006). Broiler nutrition in New Zealand – Challenges and strategies. Accessed 2013 from www.feedinfo.com
Ricke, S.C. (2003). Perspectives on the use of organic acids and short-chain fatty acids as antimicrobials. Poult. Sci. 82: 632 - 639. http://dx.doi.org/10.1093/ps/82.4.632
Savage, D. C. (1977). Microbial ecology of the gastrointestinal tract. Animal review of Microbiology 31: 107 – 133. http://dx.doi.org/10.1146/annurev,mi.31.100177.000543
Shibuya, N. & Iwasaki, T.(1985).Structural features of rice bran hemicellulose. Phytochemistry24: 285-289. http://dx.doi.org/10.1016/5003-9422(00)83538-4 Singh, A., Masey O’Neill, H.V., Ghosh, T.K., Bedford, M.R. & Haldar, S., (2012). Effects of xylanase supplementation of performance, total volatile fatty acids and selected bacterial population in caeca, metabolic indices and peptide YY concentrations in serum of broiler chickens fed energy restricted maize-soybean based diets. Anim. Feed Sci. Technol. 177, 194-203. http://dx.doi.org/10.1016/j.anifeedsci.2012.08.005
Steel, R. G. D. & Torrie, J.H.(1995). Principles and procedures of statistics, a biometric Approach (2nd Edition). Mc Graw Hill publishers, New York p.633 Tahir, M., Saleh, F., Amjed, M., Ohtsuka, A. & Hayashi, K. (2005). Synergistic effect of cellulase and hemicellulase on nutrient utilization and performance in broilers fed corn - soybean meal diet. Anim. Sci. J. 76: 559 - 565. http://dx.doi.org/10.1111/j.1740-0929.2005.00304.x
Tahir, M., Saleh, F., Ohtsuka, A. & Hayashi, K.(2008). An effective combination of carbohydrase s that enables reduction of dietary protein in broilers: Importance of hemicellulase. Poult. Sci. 87: 713 - 718. http://dx.doi.org/10.3383/ps.2007 - 00340
Tellez, G., Higgins, S., Donoghue, A. &Hargin, B. (2006).Digestive physiology and the role of microorganisms. J. App. Poult. Res. 15:136 – 144. http://dx.doi.org/10.1093/japr/151.136
Thrope,J. & Beal, J.D. (2001).Vegetable protein meals and the effects of enzymes. Pp 125 – 143 in: Enzymes in Farm Animal Nutrition. M.R. Bedford. Tierlynck, E., Bjerrum, L., Eeckhaut, V., Huygebaert, G., Pasmans, F., Haesebrouck, F., Dewulf, J., Ducatelle, R. & Van Immerseel, F.(2009). The cereal type in feed influences gut wall morphology and intestinal immune cell infiltration in broiler chickens. Br. J. Nutr. 102: 1453 - 1461.Doi: http://hdl.handle.net/1854/LU-898321
Torok, V.A., Orphei-Keller, K., Loo, M. & Hughes, R. (2008). Application of methods for identifying broiler chicken gut bacteria species linked with increased energy metabolism. Applied and Environmental Microbiol, 74: 783 - 791. http://dx.doi.org/10.1128/AEM.01384-07
Vahjen, W., Glaser, K., Schafer, K. & Simon, O.(1998). Influence of xylanase-supplemented feed on the development of selected bacterial groups in the intestinal tract of broiler chicks. J. Agric. Sci. 130: 489 - 500.Doi: http://dx.doi.org
Van Immerse, F., De Buck, J., Pasmans, F., Huyghebaert, G., Haesebrouck, F. &Ducatelle, R., (2004). Clostridium perfringens in poultry: an emerging threat for animal and public health. Avian pathology, 33:537 – 549.Doi: http://dx.doi.org/10.1080/03079450400013162
Weisburg W.G., Barns SM, Pelletier DA & Lane DJ (1991).16S Ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173: 697-703. http://dx.doi.org/0021-9193/91/020697-07$02.0010
Williams, R.B., (2005). Inter – current coccidiosis and necrotic enteritis of chicken: Rational, integrated disease management by maintenance of gut integrity. Avian Pathology 34:159 – 180.Doi: www.tandfonline.com/doi/pdf/10.1080/03079450500112195
Yang, Y., Iji, P.A. & Choct, M.(2009). Dietary modulation of gut microflora in broiler chickens: A review of the role of six kinds of alternatives to in-feed antibiotics. Worlds Poult. Sci. J. 65: 97 - 114.Doi: http://dx.doi.org/10.1017/S0043933909000087
Zanella, I., Sakomura, N.K., Silversides, F.G., Fiqueirdo, A. & Pack, M., (1999). Effect of enzyme supplementation of broiler diets based on maize and soybeans. Poult. Sci. 78, 561-568. http://dx.doi.org/10.1093/ps/78.4.56








.jpg&w=3840&q=75)