nutrition in maintaining quality of poultry products
The role of nutrition in maintaining quality of poultry products
Published: August 1, 2007
By: Steve Leeson - Department of Animal and Poultry Science, University of Guelph, Guelph, Ontario, Canada (Courtesy of Alltech Inc.)
One reason for the success of the poultry meat industry has been the consumer perception of a healthy product. Certainly poultry meat contains less fat than do comparable beef or pork products; and in most situations such fat is predominantly unsaturated.
Eggs, on the other hand, have suffered more negative consumer attitudes related to health, most of which stemmed from the issue of cholesterol metabolism as perpetuated by the medical profession in North America. While issues of the relationship between diet and plasma cholesterol in humans are now far from clear, it is obvious that overwhelming “negative press” takes many years to correct in terms of regaining market share of any food product.
Consequently, today the poultry meat and egg industries are acutely aware of quality and composition of their products as they relate to positioning relative to competing products, and also in relation to consumer attitudes and choice.
Quality, therefore, is emerging as an important issue in the poultry industry, and in a number of situations poultry nutrition can play a significant role. While it is realized that quality encompasses a number of important issues (size, conformation, microbial load, etc.), this review deals essentially in the transport of nutrients from the feed to the bird and how this influences composition relative to current consumer needs and perceptions.
Normal product composition
Both meat and eggs are excellent sources of protein and amino acids and can provide significant portions of the recommended daily intake for consumers. Nutritionally speaking, poultry meat and eggs both provide a rich source of essential amino acids, so just 100 g of poultry meat or one large egg virtually meet the adult’s requirement for amino acids (Table 1).
There seems little potential to further increasing the amino acid profile of poultry products, especially in view of the lack of knowledge shown by most consumers about the need or biological role of amino acids in human nutrition. However, both poultry meat and eggs are deficient in most minerals and vitamins. The fatty acid profile of poultry products tends to reflect that found in the diet, and this situation can be capitalized upon to manipulate fat composition of both eggs and meat.
Table 1. Content of selected nutrients in poultry meat and eggs in relation to human daily requirements.1
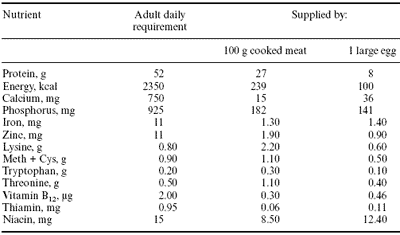
1Leeson, 1993.
PROTEIN AND AMINO ACID COMPOSITION
Virtually all the protein and amino acids in meat birds are found in the muscles, while in eggs about 50% is found in yolk and the remainder in the albumen. As a generalization, the quality and especially the composition of proteins are virtually immune to dietary manipulation.
Feeding diets deficient in amino acids can lead to reduced muscle growth or smaller egg size, yet the amino acid profile of these proteins remains intact. Even within the carcass of the chicken, the different muscles are virtually identical in amino acid composition (Table 2).
The fact that muscles as diverse as the breast and wing are so similar in composition brings into question the industry practice of trying to preferentially influence breast development via manipulation of dietary amino acid profile.
FAT COMPOSITION
Both the quantity and composition (quality) of carcass fat and eggs can be influenced by nutrition. For meat birds there is a direct relationship between diet energy level and fatness and also diet protein and fatness.
Table 2. Amino acid profile of muscles in the broiler chicken (g/100 g protein).1

1Leeson, 1993
Quality parameters today usually dictate as little fat as possible and this can be achieved by feeding high protein and/or low energy (Table 3).
Table 3. Effect of dietary protein or energy on carcass composition of broilers at 49 days.1
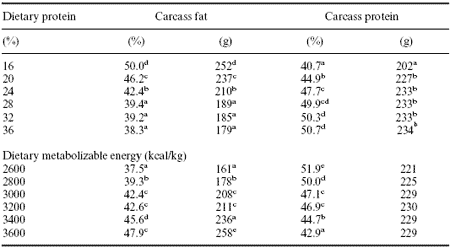
1adapted from Jackson et al. (1982).
a-eP<0.01.
Omega-3 fatty acids
There has been considerable interest in the enrichment of poultry products with omega-3 fatty acids. Increased intake of these nutrients by humans has been shown to offer some potential in reducing the risk of coronary heart disease (Kinsella et al., 1990; Kromhout, 1992).
Other studies reported that consumption of eicosapentaenoic acid (EPA) and decosahexaenoic acid (DHA) reduced serum triglyceride levels and increased platelet n-3 fatty acid content (Sanders and Roshanai, 1983; Holub et al., 1987; Harris, 1989). In addition, DHA is physiologically essential in the membrane phospholipids of the brain and retina for mental performance and visual acuity, respectively (Health and Welfare Canada, 1990).
Fish oils are rich sources of EPA and DHA and poultry meat and eggs can be enriched by simply adding these oils to layer diets. Table 4 shows results of some recent studies where we added menhaden oil to diets of broilers during the finisher period.
Table 4. Effect of dietary fish oils on egg yolk composition (%).1
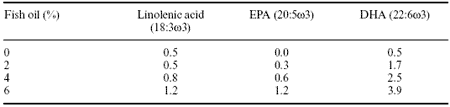
1Gonzalez and Leeson (1998, unpublished data).
A major limitation to this type of enrichment is the fishy ‘taint’ associated with EPA and DHA. In North America 2% fish oil is likely to be the maximum inclusion level for layer diets and even at this level our preliminary results suggest some problems in consumer taste panels.
The inclusion of fish oils containing EPA and DHA in the diets of laying hens also results in the enrichment of eggs with these fatty acids. Diets containing 3% menhaden oil fed to laying hens results in decreased yolk content of arachidonic acid, while EPA,DHAand linolenic acid contents increased. None of the egg production parameters measured was affected by the dietary inclusion of menhaden oil.
Other studies have involved dietary plant feedstuffs rich in n-3 fatty acids that are less likely to impart undesirable organoleptic characteristics in such enriched eggs. Caston and Leeson (1990) fed laying hens diets containing flax seed which is a source of linolenic acid (18:3 n-3) at levels of 0, 10, 20 and 30%. Fatty acid profile of the eggs showed large increases in n-3 and some n-6 fatty acids at all levels of dietary flax seed addition (Table 5).
Table 5. Effect of feeding flax on egg composition.1
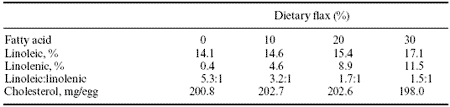
1Adapted from Caston and Leeson, 1990.
Eggs from hens fed a 30% flax seed diet had a linoleic acid:linolenic acid ratio of 1.5:1 versus 35.3:1 for the unsupplemented diet. In a similar study Cherian and Sim (1991) investigated the effects of both dietary flax seed and canola seed on yolk fatty acid composition. Sixteen percent dietary canola meal resulted in 2.4% linolenic acid in the yolk compared to 8.8% in the flax seed treatment.
However, both canola meal and flax seed increased EPA and DHA in the egg yolk to about the same level. Consistent with the results of earlier reports by Caston and Leeson (1990), these findings illustrate the possibility for enrichment of eggs with n-3 fatty acids by the use of normal dietary components. These changes in egg fatty acid profile provide an alternative or additional source of linolenic acid to the human diet.
Consumption of n-3 enriched eggs has produced changes in serum and platelet lipid composition of human subjects similar to those produced by the use of other dietary sources of n-3 fatty acids. Ferrier et al. (1992) studied the changes in serum lipids and platelet fatty acid composition in human subjects following consumption of linolenic acid enriched eggs.
Consumption of linolenic acid enriched eggs, but not regular eggs, for one or two weeks resulted in a marked decline (35%) in serum triglyceride levels with no change in total or high density lipoprotein (HDL) cholesterol.
There was an even more dramatic change in platelet phospholipid n-3 fatty acid level, where DHA content increased by some 60% in subjects consuming n-3 enriched eggs.
There are differences in the composition of n-3 fatty acids obtained as a result of using fish versus vegetable feedstuffs in the hen’s diet. Low levels of fish oil (3.0%) resulted in higher increases in the levels of DHA (3.5%) and EPA (0.52%) compared to relatively lower increases brought about by the use of much higher levels of seed meals (Hargis and Van Elswyk, 1991). The differences in the egg contents of the 20-carbon family of n-3 fatty acids is an indication of poor efficiency of conversion of linolenic acid to DHA and EPA by poultry.
Cholesterol
Cholesterol is mainly an issue with eggs, although it is interesting that consumers have little knowledge or concern about consumption of other cholesterol-rich foods such as shellfish. Eggs have been forever linked with the cholesterol issue, and in reality the only positive marketing approach is related to lowered cholesterol in any product. Unfortunately, the egg is very resistant to manipulation of its cholesterol content via layer nutrition or any other means.
Eggs are high in cholesterol because of its role in sustaining the developing embryo. Cholesterol has many and varied uses in the developing embryo including its role as a structural component of cell membranes, and as a precursor for sex and adrenal hormones, vitamin D, and the bile acids (Griffin, 1992).
Young chicks do not have the enzymes necessary for cholesterol synthesis, which emphasizes the importance of cholesterol being deposited in the egg. An egg contains about 200-250 mg cholesterol, and most of this is transferred from the yolk to the embryo during the final week of incubation (Griffin, 1992). Cholesterol content of the egg can be influenced to some extent by intakes of energy and fat (Leeson, 1993).
Diet fat per se does not seem to be a factor, although in most instances high-fat diets imply that high energy diets are used. Sutton et al. (1984) reported that restricting the energy intake of laying hens to 75% of ad libitum intake resulted in decreased total amount of cholesterol excreted via the egg, although this was associated with reduction in egg production.
The influence of dietary energy and body weight of the hen on egg cholesterol is mediated through their effects on egg size.
Reducing energy intake in order to achieve a measurable reduction in egg cholesterol concentration has the disadvantage of adversely affecting egg production.
One percent cholesterol added to the diet of laying hens increased egg cholesterol by about 25% compared to an unsupplemented group that had no change in plasma cholesterol concentration (Sutton et al., 1984).
Dietary fibre influences cholesterol metabolism by a possible combination of different processes. These include lowered cholesterol absorption, binding with bile salts in the intestinal tract, shortening the intestinal transit time, and increasing fecal sterol excretion (Hargis, 1988).
Turk and Barnett (1972) tested the effects of different fibre sources added to corn/soya diets fed laying hens on egg yolk cholesterol level. Alfalfa was the most effective of all the fibre sources tested that concomitantly had the least harmful effects on egg size, egg production and feed efficiency. In a similar study comparing the effects of different fibres, Story and Kritchevsky (1976) found that cellulose bound about 1.4% of all the bile acids tested, whereas alfalfa bound up to 15.9%. Nakaue et al. (1980) reported that the superior cholesterol lowering effect of alfalfa is not related to its saponin content.
Reduction in egg cholesterol achieved by such dietary manipulations are, however, only marginal, with little evidence to suggest a commercially significant reduction.
The reasons behind the lack of success in attempts to lower egg cholesterol content have been described by Griffin (1992) in a recent review article. It was proposed that egg cholesterol is determined by the cholesterol content of individual yolk lipoprotein moieties, rather than by the bird’s plasma cholesterol concentration. Given that most cholesterol in lipoproteins is associated with the surface layers, reduction in egg cholesterol content can therefore occur only when the lipoprotein particle size is increased. Such a scenario will reduce the contribution of surface cholesterol molecules relative to total fat.
Unfortunately, an increase in lipoprotein particle size will tend to reduce the efficiency of the critical transport of bigger sized “molecules” through the follicle wall. It seems, therefore, that meaningful reduction in egg cholesterol content will likely accrue only through the genetic manipulation of processes involved in lipoprotein synthesis and transport to the developing follicle.
Fat stability
With a trend toward replacement of saturated with unsaturated fats in poultry products there is increased concern over long term stability of both meat and eggs. In poultry nutrition this situation is accentuated due to the trend for increasing fat usage in various diets together with the fact that most fat sources today are tending towards products with high unsaturated fatty acid content (eg., poultry grease vs tallows).
Of even greater concern is the use of so-called restaurant greases, where fatty acid profile and stability are more questionable (see next section on trans fatty acids). Unstabilized fat will eventually oxidize, where the characteristic rancid taste and smell produced are due to accumulation of products such as aldehydes, ketones and hydroxy acids. Such oxidation is inevitable but can be managed by judicious use of antioxidants and cold-temperature storage.
As previously indicated, the fat composition of both eggs and meat is influenced by diet fat profile. Therefore feeding unsaturated oils (e.g., soybean oil) is likely to cause more problems than feeding palm or coconut oils. Oxidative degeneration of fats and oils begins at the CH bond next to the unsaturated C=C bond. Problems occur when so-called free radicals are generated, which have an unpaired electron resulting in instability and reactivity.
The process therefore becomes autocatalytic and once started the process of ‘rancidity’ occurs quickly and is virtually irreversible. This oxidation can occur in the feed, in the chicken and/or in the egg or meat in storage. The role of antioxidants is to become oxidized themselves by reacting with the free electron. However, unlike other fatty acids, the ‘reacted’ antioxidant is stable and so the chain reaction stops. It is obvious therefore that antioxidantsmust be added to fats as soon after manufacture as possible.
Vitamin E has long been known for its antioxidant properties. Vitamin E scavenges peroxyl free radicals that lead to further chain reactions and there is a correlation between vitamin E content of membranes and their susceptibility to oxidation. Vitamin C also plays some role as an antioxidant and complements vitamin E in being most active in the aqueous phase.
In studies with broiler chickens, Bartov and Bornstein (1978) showed that vitamin E was effective in stabilizing abdominal fat, while stability of thigh meat was only protected by synthetic antioxidants. While vitamin E still plays an important role as an antioxidant, use of new types of processed fats and oils often dictates the need to select more effective synthetic antioxidants.
Our recent studies outline the problem encountered when using oxidized fat in diets for young birds (Table 6). Young broilers digested the oxidized fat very poorly and consequently there was reduced diet apparent metabolizable energy (corrected for nitrogen) (AMEn) and slower early growth. There was also a significant increase in liver size, suggestive of abnormal metabolism.
Of more direct interest in terms of meat quality is the effect of rancid fat on the accumulation of peroxide type products in meat or eggs. Lin et al (1989) show significant accumulation of oxidation products in thigh meat of broilers even during storage at –20wC (Table 7). Oxidized oil obviously caused a substantial rise in thigh meat thiobarbituric acid (TBA) indicative of oxidative rancidity. The levels achieved after two months storage at –20wC were of sufficient magnitude to cause potential organoleptic problems.
As previously discussed, a more recent concern with fat quality is the trend to incorporation of longer chain fatty acids (w-3, w-6) etc. where rate and susceptibility to rancidity may be greater. To some extent this situation can be counteracted by judicious use of antioxidants. Table 8 shows oxidative rancidity of high w-3 eggs stored at 4 or 25wC for up to five weeks.
Table 6. Effect of fat oxidation on early broiler growth and development.

1Namkung and Leeson (unpublished data).
Table 7. Effect of feeding regular or oxidized fat with or without antioxidants on thigh meat TBA values.1
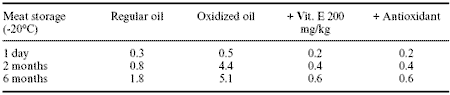
1Adapted from Lin et al. (1989).
Table 8. Effect of storage time and temperature on malondialdehyde accumulation in regular or high omega-3 eggs (μg/g egg yolk).1
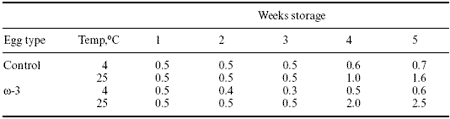
1Leeson and Squires (unpublished data).
At 4wC, which is the recommended temperature for egg storage, there is minimal rancidity of the w-3 eggs, and accumulation of malondialdehyde is comparable to that occurring in conventional eggs. At 25wC, which is obviously an unusual storage condition, there is greater rancidity of the w-3 eggs, but not until after three weeks storage.
Trans fatty acids
There is current concern about the trans fatty acid content of poultry products, which develop as a result of feeding hydrogenated restaurant fats and oils. Deep-fat frying now represents a major use of edible oils in North America; and the trend in recent years has been for these restaurants to replace tallows and lards with vegetable oils. This change has been undertaken on the basis of consumer concerns about saturated fats.
However, vegetable oils are notoriously unstable, and so they are often hydrogenated as a means of ensuring greater stability for repeated or continuous cooking procedures. During hydrogenation the level of polyunsaturates is reduced, but concomitantly there is increased formation of so called trans fatty acids. Trans fatty acids are isomers of the naturally occurring cis fatty acids. Trans fatty acids have been associated with impaired lipid metabolism in general and especially in an animal’s ability to synthesize essential fatty acids and prostaglandins (Kinsella et al., 1980).
In assaying a number of commercial grade feed fats, Al-Athari and Watkins (1988a) found trans acids of 18:1 to range from 0-11.7% while levels of trans 18:2 weremuch lower and rarely exceeded 1%. Emken (1984) suggests that hydrogenated soybean oil will usually contain around 12% trans 18:1 and about 9% other trans fatty acids. Al-Athari andWatkins (1988b) showed that chicks fed trans fatty acids in the form of hydrogenated soybean oil had poorer performance and transfatty acids accumulated in the abdominal fat and liver. These workers concluded that trans fatty acids impaired essential fatty acid metabolism in the chick. There seems little doubt that dietary trans fatty acids are deposited in the tissue of most animals (Bonaga et al., 1980; Elson et al., 1981) and so this becomes a quality issue for the poultry industry.
The uptake of trans fatty acids by humans eating poultry meat or eggs, even from birds fed quite high levels of trans fatty acids, will be quite small. In relation to consumption of margarines, etc., the contribution of poultry products is minimal. Even cows’ milk naturally contains 5-10 times the levels of trans fatty acids (due to hydrogenation action of rumen microbes) as found in poultry products. Our main concern should perhaps be bird health and performance, but with today’s impressionable consumer, even trace amounts of potentially hazardous nutrients can become a marketing nightmare.
Although there are only likely to be relatively low amounts of trans fatty acids accumulated in broilers or turkeys fed hydrogenated restaurant greases, there is long-termconcern about accumulation caused by continual recycling of poultry fats within integrated operations. There is no work currently available on the effect of continually (eg., 4-6 cycles) recycling poultry fat that initially contains moderate levels of trans fatty acids. In this same context there is concern about accumulation of non-volatile breakdown products from cooking fats that are known to be toxic and will be present to some degree in restaurant greases. Unfortunately, use of restaurant greases per se is difficult to rationalize in attempting to optimize the quality of poultry products.
Eggs, on the other hand, have suffered more negative consumer attitudes related to health, most of which stemmed from the issue of cholesterol metabolism as perpetuated by the medical profession in North America. While issues of the relationship between diet and plasma cholesterol in humans are now far from clear, it is obvious that overwhelming “negative press” takes many years to correct in terms of regaining market share of any food product.
Consequently, today the poultry meat and egg industries are acutely aware of quality and composition of their products as they relate to positioning relative to competing products, and also in relation to consumer attitudes and choice.
Quality, therefore, is emerging as an important issue in the poultry industry, and in a number of situations poultry nutrition can play a significant role. While it is realized that quality encompasses a number of important issues (size, conformation, microbial load, etc.), this review deals essentially in the transport of nutrients from the feed to the bird and how this influences composition relative to current consumer needs and perceptions.
Normal product composition
Both meat and eggs are excellent sources of protein and amino acids and can provide significant portions of the recommended daily intake for consumers. Nutritionally speaking, poultry meat and eggs both provide a rich source of essential amino acids, so just 100 g of poultry meat or one large egg virtually meet the adult’s requirement for amino acids (Table 1).
There seems little potential to further increasing the amino acid profile of poultry products, especially in view of the lack of knowledge shown by most consumers about the need or biological role of amino acids in human nutrition. However, both poultry meat and eggs are deficient in most minerals and vitamins. The fatty acid profile of poultry products tends to reflect that found in the diet, and this situation can be capitalized upon to manipulate fat composition of both eggs and meat.
Table 1. Content of selected nutrients in poultry meat and eggs in relation to human daily requirements.1
1Leeson, 1993.
PROTEIN AND AMINO ACID COMPOSITION
Virtually all the protein and amino acids in meat birds are found in the muscles, while in eggs about 50% is found in yolk and the remainder in the albumen. As a generalization, the quality and especially the composition of proteins are virtually immune to dietary manipulation.
Feeding diets deficient in amino acids can lead to reduced muscle growth or smaller egg size, yet the amino acid profile of these proteins remains intact. Even within the carcass of the chicken, the different muscles are virtually identical in amino acid composition (Table 2).
The fact that muscles as diverse as the breast and wing are so similar in composition brings into question the industry practice of trying to preferentially influence breast development via manipulation of dietary amino acid profile.
FAT COMPOSITION
Both the quantity and composition (quality) of carcass fat and eggs can be influenced by nutrition. For meat birds there is a direct relationship between diet energy level and fatness and also diet protein and fatness.
Table 2. Amino acid profile of muscles in the broiler chicken (g/100 g protein).1
1Leeson, 1993
Quality parameters today usually dictate as little fat as possible and this can be achieved by feeding high protein and/or low energy (Table 3).
Table 3. Effect of dietary protein or energy on carcass composition of broilers at 49 days.1
1adapted from Jackson et al. (1982).
a-eP<0.01.
Omega-3 fatty acids
There has been considerable interest in the enrichment of poultry products with omega-3 fatty acids. Increased intake of these nutrients by humans has been shown to offer some potential in reducing the risk of coronary heart disease (Kinsella et al., 1990; Kromhout, 1992).
Other studies reported that consumption of eicosapentaenoic acid (EPA) and decosahexaenoic acid (DHA) reduced serum triglyceride levels and increased platelet n-3 fatty acid content (Sanders and Roshanai, 1983; Holub et al., 1987; Harris, 1989). In addition, DHA is physiologically essential in the membrane phospholipids of the brain and retina for mental performance and visual acuity, respectively (Health and Welfare Canada, 1990).
Fish oils are rich sources of EPA and DHA and poultry meat and eggs can be enriched by simply adding these oils to layer diets. Table 4 shows results of some recent studies where we added menhaden oil to diets of broilers during the finisher period.
Table 4. Effect of dietary fish oils on egg yolk composition (%).1
1Gonzalez and Leeson (1998, unpublished data).
A major limitation to this type of enrichment is the fishy ‘taint’ associated with EPA and DHA. In North America 2% fish oil is likely to be the maximum inclusion level for layer diets and even at this level our preliminary results suggest some problems in consumer taste panels.
The inclusion of fish oils containing EPA and DHA in the diets of laying hens also results in the enrichment of eggs with these fatty acids. Diets containing 3% menhaden oil fed to laying hens results in decreased yolk content of arachidonic acid, while EPA,DHAand linolenic acid contents increased. None of the egg production parameters measured was affected by the dietary inclusion of menhaden oil.
Other studies have involved dietary plant feedstuffs rich in n-3 fatty acids that are less likely to impart undesirable organoleptic characteristics in such enriched eggs. Caston and Leeson (1990) fed laying hens diets containing flax seed which is a source of linolenic acid (18:3 n-3) at levels of 0, 10, 20 and 30%. Fatty acid profile of the eggs showed large increases in n-3 and some n-6 fatty acids at all levels of dietary flax seed addition (Table 5).
Table 5. Effect of feeding flax on egg composition.1
1Adapted from Caston and Leeson, 1990.
Eggs from hens fed a 30% flax seed diet had a linoleic acid:linolenic acid ratio of 1.5:1 versus 35.3:1 for the unsupplemented diet. In a similar study Cherian and Sim (1991) investigated the effects of both dietary flax seed and canola seed on yolk fatty acid composition. Sixteen percent dietary canola meal resulted in 2.4% linolenic acid in the yolk compared to 8.8% in the flax seed treatment.
However, both canola meal and flax seed increased EPA and DHA in the egg yolk to about the same level. Consistent with the results of earlier reports by Caston and Leeson (1990), these findings illustrate the possibility for enrichment of eggs with n-3 fatty acids by the use of normal dietary components. These changes in egg fatty acid profile provide an alternative or additional source of linolenic acid to the human diet.
Consumption of n-3 enriched eggs has produced changes in serum and platelet lipid composition of human subjects similar to those produced by the use of other dietary sources of n-3 fatty acids. Ferrier et al. (1992) studied the changes in serum lipids and platelet fatty acid composition in human subjects following consumption of linolenic acid enriched eggs.
Consumption of linolenic acid enriched eggs, but not regular eggs, for one or two weeks resulted in a marked decline (35%) in serum triglyceride levels with no change in total or high density lipoprotein (HDL) cholesterol.
There was an even more dramatic change in platelet phospholipid n-3 fatty acid level, where DHA content increased by some 60% in subjects consuming n-3 enriched eggs.
There are differences in the composition of n-3 fatty acids obtained as a result of using fish versus vegetable feedstuffs in the hen’s diet. Low levels of fish oil (3.0%) resulted in higher increases in the levels of DHA (3.5%) and EPA (0.52%) compared to relatively lower increases brought about by the use of much higher levels of seed meals (Hargis and Van Elswyk, 1991). The differences in the egg contents of the 20-carbon family of n-3 fatty acids is an indication of poor efficiency of conversion of linolenic acid to DHA and EPA by poultry.
Cholesterol
Cholesterol is mainly an issue with eggs, although it is interesting that consumers have little knowledge or concern about consumption of other cholesterol-rich foods such as shellfish. Eggs have been forever linked with the cholesterol issue, and in reality the only positive marketing approach is related to lowered cholesterol in any product. Unfortunately, the egg is very resistant to manipulation of its cholesterol content via layer nutrition or any other means.
Eggs are high in cholesterol because of its role in sustaining the developing embryo. Cholesterol has many and varied uses in the developing embryo including its role as a structural component of cell membranes, and as a precursor for sex and adrenal hormones, vitamin D, and the bile acids (Griffin, 1992).
Young chicks do not have the enzymes necessary for cholesterol synthesis, which emphasizes the importance of cholesterol being deposited in the egg. An egg contains about 200-250 mg cholesterol, and most of this is transferred from the yolk to the embryo during the final week of incubation (Griffin, 1992). Cholesterol content of the egg can be influenced to some extent by intakes of energy and fat (Leeson, 1993).
Diet fat per se does not seem to be a factor, although in most instances high-fat diets imply that high energy diets are used. Sutton et al. (1984) reported that restricting the energy intake of laying hens to 75% of ad libitum intake resulted in decreased total amount of cholesterol excreted via the egg, although this was associated with reduction in egg production.
The influence of dietary energy and body weight of the hen on egg cholesterol is mediated through their effects on egg size.
Reducing energy intake in order to achieve a measurable reduction in egg cholesterol concentration has the disadvantage of adversely affecting egg production.
One percent cholesterol added to the diet of laying hens increased egg cholesterol by about 25% compared to an unsupplemented group that had no change in plasma cholesterol concentration (Sutton et al., 1984).
Dietary fibre influences cholesterol metabolism by a possible combination of different processes. These include lowered cholesterol absorption, binding with bile salts in the intestinal tract, shortening the intestinal transit time, and increasing fecal sterol excretion (Hargis, 1988).
Turk and Barnett (1972) tested the effects of different fibre sources added to corn/soya diets fed laying hens on egg yolk cholesterol level. Alfalfa was the most effective of all the fibre sources tested that concomitantly had the least harmful effects on egg size, egg production and feed efficiency. In a similar study comparing the effects of different fibres, Story and Kritchevsky (1976) found that cellulose bound about 1.4% of all the bile acids tested, whereas alfalfa bound up to 15.9%. Nakaue et al. (1980) reported that the superior cholesterol lowering effect of alfalfa is not related to its saponin content.
Reduction in egg cholesterol achieved by such dietary manipulations are, however, only marginal, with little evidence to suggest a commercially significant reduction.
The reasons behind the lack of success in attempts to lower egg cholesterol content have been described by Griffin (1992) in a recent review article. It was proposed that egg cholesterol is determined by the cholesterol content of individual yolk lipoprotein moieties, rather than by the bird’s plasma cholesterol concentration. Given that most cholesterol in lipoproteins is associated with the surface layers, reduction in egg cholesterol content can therefore occur only when the lipoprotein particle size is increased. Such a scenario will reduce the contribution of surface cholesterol molecules relative to total fat.
Unfortunately, an increase in lipoprotein particle size will tend to reduce the efficiency of the critical transport of bigger sized “molecules” through the follicle wall. It seems, therefore, that meaningful reduction in egg cholesterol content will likely accrue only through the genetic manipulation of processes involved in lipoprotein synthesis and transport to the developing follicle.
Fat stability
With a trend toward replacement of saturated with unsaturated fats in poultry products there is increased concern over long term stability of both meat and eggs. In poultry nutrition this situation is accentuated due to the trend for increasing fat usage in various diets together with the fact that most fat sources today are tending towards products with high unsaturated fatty acid content (eg., poultry grease vs tallows).
Of even greater concern is the use of so-called restaurant greases, where fatty acid profile and stability are more questionable (see next section on trans fatty acids). Unstabilized fat will eventually oxidize, where the characteristic rancid taste and smell produced are due to accumulation of products such as aldehydes, ketones and hydroxy acids. Such oxidation is inevitable but can be managed by judicious use of antioxidants and cold-temperature storage.
As previously indicated, the fat composition of both eggs and meat is influenced by diet fat profile. Therefore feeding unsaturated oils (e.g., soybean oil) is likely to cause more problems than feeding palm or coconut oils. Oxidative degeneration of fats and oils begins at the CH bond next to the unsaturated C=C bond. Problems occur when so-called free radicals are generated, which have an unpaired electron resulting in instability and reactivity.
The process therefore becomes autocatalytic and once started the process of ‘rancidity’ occurs quickly and is virtually irreversible. This oxidation can occur in the feed, in the chicken and/or in the egg or meat in storage. The role of antioxidants is to become oxidized themselves by reacting with the free electron. However, unlike other fatty acids, the ‘reacted’ antioxidant is stable and so the chain reaction stops. It is obvious therefore that antioxidantsmust be added to fats as soon after manufacture as possible.
Vitamin E has long been known for its antioxidant properties. Vitamin E scavenges peroxyl free radicals that lead to further chain reactions and there is a correlation between vitamin E content of membranes and their susceptibility to oxidation. Vitamin C also plays some role as an antioxidant and complements vitamin E in being most active in the aqueous phase.
In studies with broiler chickens, Bartov and Bornstein (1978) showed that vitamin E was effective in stabilizing abdominal fat, while stability of thigh meat was only protected by synthetic antioxidants. While vitamin E still plays an important role as an antioxidant, use of new types of processed fats and oils often dictates the need to select more effective synthetic antioxidants.
Our recent studies outline the problem encountered when using oxidized fat in diets for young birds (Table 6). Young broilers digested the oxidized fat very poorly and consequently there was reduced diet apparent metabolizable energy (corrected for nitrogen) (AMEn) and slower early growth. There was also a significant increase in liver size, suggestive of abnormal metabolism.
Of more direct interest in terms of meat quality is the effect of rancid fat on the accumulation of peroxide type products in meat or eggs. Lin et al (1989) show significant accumulation of oxidation products in thigh meat of broilers even during storage at –20wC (Table 7). Oxidized oil obviously caused a substantial rise in thigh meat thiobarbituric acid (TBA) indicative of oxidative rancidity. The levels achieved after two months storage at –20wC were of sufficient magnitude to cause potential organoleptic problems.
As previously discussed, a more recent concern with fat quality is the trend to incorporation of longer chain fatty acids (w-3, w-6) etc. where rate and susceptibility to rancidity may be greater. To some extent this situation can be counteracted by judicious use of antioxidants. Table 8 shows oxidative rancidity of high w-3 eggs stored at 4 or 25wC for up to five weeks.
Table 6. Effect of fat oxidation on early broiler growth and development.
1Namkung and Leeson (unpublished data).
Table 7. Effect of feeding regular or oxidized fat with or without antioxidants on thigh meat TBA values.1
1Adapted from Lin et al. (1989).
Table 8. Effect of storage time and temperature on malondialdehyde accumulation in regular or high omega-3 eggs (μg/g egg yolk).1
1Leeson and Squires (unpublished data).
At 4wC, which is the recommended temperature for egg storage, there is minimal rancidity of the w-3 eggs, and accumulation of malondialdehyde is comparable to that occurring in conventional eggs. At 25wC, which is obviously an unusual storage condition, there is greater rancidity of the w-3 eggs, but not until after three weeks storage.
Trans fatty acids
There is current concern about the trans fatty acid content of poultry products, which develop as a result of feeding hydrogenated restaurant fats and oils. Deep-fat frying now represents a major use of edible oils in North America; and the trend in recent years has been for these restaurants to replace tallows and lards with vegetable oils. This change has been undertaken on the basis of consumer concerns about saturated fats.
However, vegetable oils are notoriously unstable, and so they are often hydrogenated as a means of ensuring greater stability for repeated or continuous cooking procedures. During hydrogenation the level of polyunsaturates is reduced, but concomitantly there is increased formation of so called trans fatty acids. Trans fatty acids are isomers of the naturally occurring cis fatty acids. Trans fatty acids have been associated with impaired lipid metabolism in general and especially in an animal’s ability to synthesize essential fatty acids and prostaglandins (Kinsella et al., 1980).
In assaying a number of commercial grade feed fats, Al-Athari and Watkins (1988a) found trans acids of 18:1 to range from 0-11.7% while levels of trans 18:2 weremuch lower and rarely exceeded 1%. Emken (1984) suggests that hydrogenated soybean oil will usually contain around 12% trans 18:1 and about 9% other trans fatty acids. Al-Athari andWatkins (1988b) showed that chicks fed trans fatty acids in the form of hydrogenated soybean oil had poorer performance and transfatty acids accumulated in the abdominal fat and liver. These workers concluded that trans fatty acids impaired essential fatty acid metabolism in the chick. There seems little doubt that dietary trans fatty acids are deposited in the tissue of most animals (Bonaga et al., 1980; Elson et al., 1981) and so this becomes a quality issue for the poultry industry.
The uptake of trans fatty acids by humans eating poultry meat or eggs, even from birds fed quite high levels of trans fatty acids, will be quite small. In relation to consumption of margarines, etc., the contribution of poultry products is minimal. Even cows’ milk naturally contains 5-10 times the levels of trans fatty acids (due to hydrogenation action of rumen microbes) as found in poultry products. Our main concern should perhaps be bird health and performance, but with today’s impressionable consumer, even trace amounts of potentially hazardous nutrients can become a marketing nightmare.
Although there are only likely to be relatively low amounts of trans fatty acids accumulated in broilers or turkeys fed hydrogenated restaurant greases, there is long-termconcern about accumulation caused by continual recycling of poultry fats within integrated operations. There is no work currently available on the effect of continually (eg., 4-6 cycles) recycling poultry fat that initially contains moderate levels of trans fatty acids. In this same context there is concern about accumulation of non-volatile breakdown products from cooking fats that are known to be toxic and will be present to some degree in restaurant greases. Unfortunately, use of restaurant greases per se is difficult to rationalize in attempting to optimize the quality of poultry products.
Vitamin composition
Consumers are aware of vitamin content of foods as a quality issue. The cereal industry has labeled their products for vitamins for many years, even though their contribution of certain vitamins is quite low. Consumer surveys have shown that vitamin enriched cereals are selected over non-enriched products when price is comparable. Recent analysis of eggs from different sources shows high variability in their vitamin contents (Naber and Squires, 1993). The results show a greater range in egg content of vitamins A and B12 but a lower range for riboflavin.
This variability in egg vitamin content is of concern when consistency of food composition is desired, and particularly when one considers the nutrient labeling of foods (Naber, 1993). There is potential to increase the vitamin content of both meat and eggs although to date most attention has centred on eggs. Layer nutrition is a major factor influencing egg vitamin content.
However, the influence of dietary vitamin intake on vitamin enrichment of eggs is very variable among different vitamins. Squires and Naber (1993a) reported that egg riboflavin level in yolk and albumen responded rapidly to the dietary level of this vitamin. Similarly, the egg content of vitamin B12 was almost exactly proportional to diet content over one to four times the NRC (1984) requirement for the hen (Squires and Naber, 1992). There did not seem to be a ceiling on vitamin B12 transfer to the eggs even when up to four times the NRC requirement were given to the hens (Naber, 1993).
There is, however, a ceiling on the amount of riboflavin that the laying hens can transfer to the egg when fed diets containing between two and four times their requirement as established by NRC (1984). On the other hand, when laying hens were fed vitaminAat four times their requirement, the egg content of vitamin A responded only partially (Squires and Naber, 1993b). This was attributed to the modulating effect and storage capacity of vitamin A in the liver.
Naber (1993), in a review of factors influencing egg vitamin content, concluded that feed vitamin content has the greatest and most widespread influence on egg vitamin content. Using data from studies that reported diet vitamin level and feed intake on the one hand and egg output, i.e., egg weight and production on the other, Naber (1993) calculated the efficiency of vitamin transfer into eggs as a function of intake (Table 9).
Table 9. Classification of vitamins by relative transfer efficiency from diet to egg.1
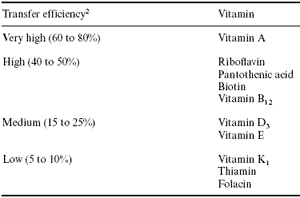
1Adapted from Naber, 1993.
2When the vitamin is fed at one or two times the NRC dietary requirement.
The transfer efficiency of vitamin A at dietary levels equivalent to one or two times the NRC requirement was very high (up to 80%), but this dropped markedly when the dietary level was raised to four times requirement (Squires and Naber, 1993b). The increase in egg yolk vitamin A content of some 67% in response to dietary increase from one to two times the requirement is consistent with results of similar studies reported earlier by Reddy et al. (1978). This is an indication of the possibility of egg enrichment with vitamin A, even though this trend declines at high levels of diet vitamin enrichment.
The transfer of dietary vitamin B12 into eggs was as efficient as for riboflavin, pantothenic acid and biotin, i.e., about 50% with dietary levels at one to two times requirement. Unlike riboflavin, however, this level of transfer efficiency continued in the case of vitamin B12 even at very high dietary levels of up to 40 times requirement. Clearly, extensive enrichment of eggs with vitamin B12 is possible (Naber, 1993).
Consumers are aware of vitamin content of foods as a quality issue. The cereal industry has labeled their products for vitamins for many years, even though their contribution of certain vitamins is quite low. Consumer surveys have shown that vitamin enriched cereals are selected over non-enriched products when price is comparable. Recent analysis of eggs from different sources shows high variability in their vitamin contents (Naber and Squires, 1993). The results show a greater range in egg content of vitamins A and B12 but a lower range for riboflavin.
This variability in egg vitamin content is of concern when consistency of food composition is desired, and particularly when one considers the nutrient labeling of foods (Naber, 1993). There is potential to increase the vitamin content of both meat and eggs although to date most attention has centred on eggs. Layer nutrition is a major factor influencing egg vitamin content.
However, the influence of dietary vitamin intake on vitamin enrichment of eggs is very variable among different vitamins. Squires and Naber (1993a) reported that egg riboflavin level in yolk and albumen responded rapidly to the dietary level of this vitamin. Similarly, the egg content of vitamin B12 was almost exactly proportional to diet content over one to four times the NRC (1984) requirement for the hen (Squires and Naber, 1992). There did not seem to be a ceiling on vitamin B12 transfer to the eggs even when up to four times the NRC requirement were given to the hens (Naber, 1993).
There is, however, a ceiling on the amount of riboflavin that the laying hens can transfer to the egg when fed diets containing between two and four times their requirement as established by NRC (1984). On the other hand, when laying hens were fed vitaminAat four times their requirement, the egg content of vitamin A responded only partially (Squires and Naber, 1993b). This was attributed to the modulating effect and storage capacity of vitamin A in the liver.
Naber (1993), in a review of factors influencing egg vitamin content, concluded that feed vitamin content has the greatest and most widespread influence on egg vitamin content. Using data from studies that reported diet vitamin level and feed intake on the one hand and egg output, i.e., egg weight and production on the other, Naber (1993) calculated the efficiency of vitamin transfer into eggs as a function of intake (Table 9).
Table 9. Classification of vitamins by relative transfer efficiency from diet to egg.1
1Adapted from Naber, 1993.
2When the vitamin is fed at one or two times the NRC dietary requirement.
The transfer efficiency of vitamin A at dietary levels equivalent to one or two times the NRC requirement was very high (up to 80%), but this dropped markedly when the dietary level was raised to four times requirement (Squires and Naber, 1993b). The increase in egg yolk vitamin A content of some 67% in response to dietary increase from one to two times the requirement is consistent with results of similar studies reported earlier by Reddy et al. (1978). This is an indication of the possibility of egg enrichment with vitamin A, even though this trend declines at high levels of diet vitamin enrichment.
The transfer of dietary vitamin B12 into eggs was as efficient as for riboflavin, pantothenic acid and biotin, i.e., about 50% with dietary levels at one to two times requirement. Unlike riboflavin, however, this level of transfer efficiency continued in the case of vitamin B12 even at very high dietary levels of up to 40 times requirement. Clearly, extensive enrichment of eggs with vitamin B12 is possible (Naber, 1993).
| Conclusions While egg and poultry meat products are well positioned relative to other animal proteins in terms of “marketable” nutrient profile, there is potential for improvement in perceived quality. Such quality issues are enrichment of fatty acids now deemed useful in human nutrition together with limitation of undesirable products such as lipid peroxides and trans fatty acids. Once thought of as only valid for the insignificant ‘health-store’ market, designer poultry products are now entering mainstream marketing. |
References
Al-Athari, A.K. and B.A.Watkins. 1988a. Chromatographic analysis of 18:1 isomers in blended feed-grade fats used in poultry diets. Poultry Sci. 67: 307-312.
Al-Athari, A.K. and B.A. Watkins. 1988b. Distribution of trans and cis 18:1 fatty acid isomers in chicks fed different fats. Poultry Sci. 67: 778-786.
Bartov, I. and S. Bornstein. 1978. Stability of abdominal fat and meat of broilers: Effect of duration of feeding antioxidants. Br. Poultry Sci. 19: 129-135.
Bonaga, C., M.G. Trizzino, M.A. Pasquariello and P.L. Biagi. 1980. Nutritional aspects of trans fatty acids. Note I. Their accumulation in tissue lipids of rats with normolipidic diets containing margarine. Biochem. Exp. Biol. 16: 51-4.
Caston, L. and S. Leeson. 1990. Dietary flax and egg composition. Poultry Sci. 69: 1617-20.
Cherian, G. and J.S. Sim. 1991. Effect of feeding full fat flax and canola seeds to laying hens on the fatty acid composition of eggs, embryos and newly hatched chicks. Poultry Sci. 70: 917-22.
Elson, C.E., N.J. Benevenga, D.J. Canty, R.H. Grummer, J.J. Lalich, J.W. Porter and A.E. Johnson. 1981. The influence of dietary unsaturated cis and trans and saturated fatty acids on tissue lipids of swine. Atherosclerosis 40: 115-37.
Emken, E.A., 1984. Nutrition and biochemistry of trans and positional fatty acid isomers in hydrogenated oils. Annu. Rev. Nutr. 4:339-376.
Ferrier, L.K., L. Caston, S. Leeson, E.J. Squires, B. Celi, L. Thomas and B.J. Holub. 1992. Changes in serum lipids and platelet fatty acid composition following consumption of eggs enriched in alpha-linolenic acid (LnA). Food Res. Intern. 25: 263.
Griffin, H.D. 1992. Manipulation of egg yolk cholesterol: A physiologist’s view. World’s Poultry Sci. J. 48: 101-112.
Hargis, P.S., 1988. Modifying egg yolk cholesterol in the domestic fowl - a review. World’s Poultry Sci. 44:17-29.
Hargis, P.S. and M.E. Van Elswyk, 1991. Modifying yolk fatty acid composition to improve health quality of eggs. Pages 249-260 in C. Haberston and C.E. Morris, eds. Fat and cholesterol reduced foods: technologies and strategies. Gulf Publishing Co., Houston, U.S.A.
Harris,W.S., 1989. Fish oils and plasma lipid and lipoprotein metabolism in humans: a critical review. J. Lipid Res. 30:785-807.
Health and Welfare Canada, 1990. Nutrition recommendations. Minister of Supply and Services, Ottawa, ON.
Holub, B.J., D.J. Bakker and C.M. Skeaff, 1987. Alterations in molecular species of cholesterol esters formed via plasma lecithin-cholesterol acyltransferase in human subjects consuming fish oil. Atherosci. 66:11-18.
Jackson, S., J.D. Summers and S. Leeson. 1982. Effect of dietary protein and energy on broiler carcass composition and efficiency of nutrient utilization. Poultry Sci. 61: 2224-2231.
Kinsella, J.E., G. Bruckner, J. Mai and J. Shimp. 1980. Metabolism of trans fatty acids with emphasis on the effects of trans, transoctadecadienoate on lipid composition, essential fatty acid, and prostaglandins: an overview. Am. J. Clin. Nutr. 34: 2307-18.
Kinsella, J.E., B. Lokesh and R.A. Stone. 1990. Dietary n-3 polyunsaturated fatty acids and amelioration of cardiovascular disease: possible mechanism. Am. J. Clin. Nutr. 52:1-28.
Kromhout,D., 1992. Dietary fats: Long-termimplications for health. Nutr. Rev. 50:49-53.
Leeson, S. 1993. Potential of modifying poultry products. J. Appl. Poultry Res. 2: 380-384.
Lin, C.F., A. Ashgar, J.I. Gray, D.J. Buckley, A.M. Booren, R.L. Crackel and C.J. Flegal. 1989. Effects of oxidized dietary oil and antioxidant supplementation on broiler growth and meat stability. Br. Poultry Sci. 30: 855-864.
Naber, E.C. 1993. Modifying vitamin composition of eggs: A review. J. Appl. Poultry Res. 2: 385-393.
Naber, E.C. and M.W. Squires. 1993. Vitamin profiles as indicators of nutritional status in the laying hen: Diet to egg transfer and commercial flock survey. Poultry Sci. 72: 1046-1053.
Nakaue, N.S., R.R. Lowry, P.R. Sheeke, andG.H. Arscott, 1980. The effect of dietary alfalfa of varying saponin cotent on yolk cholesterol level and layer performance. Poutlry Sci. 59:2744-2748.
National Research Council, 1984. Nutrient requirements of Poultry. 8th rev. ed. National Academy Press, Washington, DC.
Reddy, V.R., B. Panda and V.R. Sadagopan. 1978. Relation of different levels of dietary vitamin A of laying hens to vitamin A content of egg yolk and newly hatched chicks. Ind. Vet. J. 55: 554-558.
Sanders, T.A. and F. Roshanai. 1983. The influence of different types of omega-3 polyunsaturated fatty acids on blood lipids and platelet function in healthy volunteers. Clin. Sci. 64: 91-99.
Squires, M.W. and E.C. Naber. 1992. Vitamin profiles of eggs as indicators of nutritional status in the laying hen: Vitamin B12 study. Poultry Sci. 71: 2075-2082.
Squires, M.W. and E.C. Naber. 1993a. Vitamin profiles of eggs as indicators of nutritional status in the laying hen: Riboflavin study. Poultry Sci. 72: 483-484.
Squires, M.W. and E.C. Naber. 1993b. Vitamin profiles of eggs as indicators of nutritional status in the laying hen: Vitamin A study. Poultry Sci. 72: 154-164.
Story, J.A. andD. Kritchevsky. 1976. Comparison of the binding of various bile acids and bile salts in vitro by several types of fibre. J. Nutr. 106: 1292-1294.
Sutton, C.C.,W.M. Muir andG.E. Mitchell. 1984. Cholesterol metabolism in the laying as influenced by cholesterol, caloric intake and genotype. Poultry Sci. 63: 972-980.
Turk, D.E. and B.D. Bernett. 1972. Diet and egg cholesterol content. Poultry Sci. 51(Suppl. 1): 1881 (Abstr.)
Related topics
Authors:

Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post






You may be interested in
_%20Why%20are%20egg%20prices%20on%20the%20rise(10min).jpg&w=3840&q=75)