Non-phytate phosphorus requirement for male broilers subjected to two calcium supply regimens from 8 to 21 days of age under thermoneutral environment conditions
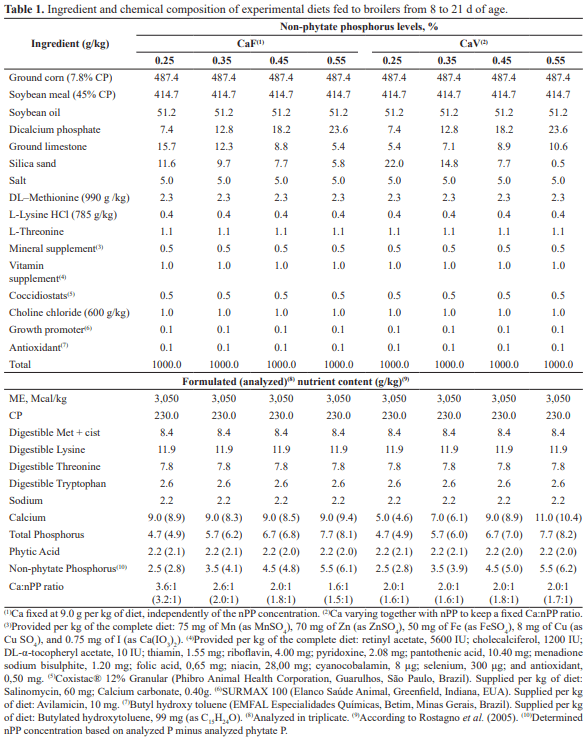
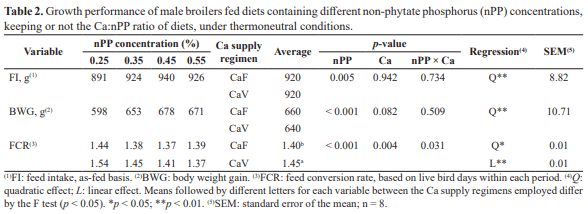
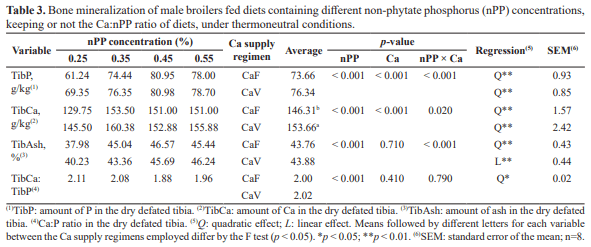
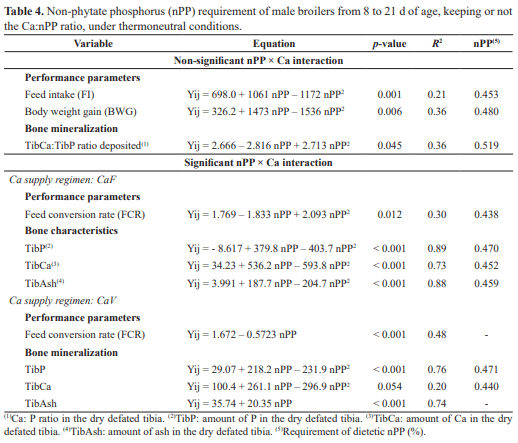
The purpose of this study was to determine the non-phytate phosphorus (nPP) requirement for male broiler subjected to two calcium supply regimens from 8 to 21 d of age, based on performance and bone mineralization. Birds were distributed in a complete randomized 4 × 2 factorial design with four nPP concentrations: 0.25, 0.35, 0.45 and 0.55%, and two Ca supply regimens: Ca fixed at 9.0 g/kg of diet (CaF) or varying together with the nPP concentrations tested keeping a fixed Ca:nPP ratio (CaV), with 8 battery cages/treatment and 9 birds/battery cage. Feed intake (FI), body weight gain (BWG) and Ca:P ratio deposited in the dry defatted tibia (TibCa:TibP) were quadratically increased by the increasing nPP concentrations, regardless of the Ca supply regimen adopted (p<0.01). The nPP levels tested also influenced FCR and TibAsh, which presented a quadratic and linear response when CaF and CaV diets were used, respectively, and TibP and TibCa, which presented a quadratic response with both Ca supply regimen, CaF and CaV diets. Also, CaV diets provided a 2.86 and 5.02% higher FCR (p<0.05) and TibCa (p<0.01), respectively, when compared to CaF diets. Therefore, nPP nutritional requirement for male broilers reared at thermoneutral conditions from 8 to 21 d of age that provided better performance and bone mineralization were, respectively, 0.480 and 0.459% with CaF and 0.550% with CaV diets, indicating that, under thermoneutral conditions, CaV diets negatively affected growth performance of broilers, while positively affecting bone mineralization especially when low nPP levels are applied.
Additional keywords: mineralization; nutritional requirement; performance; thermal comfort; tibia.
Angel R, 2011. Calcium and phosphorus requirements in broilers. III Int Symp on Nutritional Requirements of Poultry and Swine, Viçosa (Brazil), March 29-31. pp: 77-96.
Ankra-Badu GA, Aggrey SE, Pesti GM, Bakalli RI, Edwards HM, 2004. Modeling of parameters affecting phytate phosphorus bioavailability in growing birds. Poult Sci 83: 1083-1088. https://doi.org/10.1093/ps/83.7.1083
AOAC, 2000. Official methods of analysis, 17th edn. AOAC Int, MD, USA.
Boling SD, Douglas MW, Johnson ML, Wang X, Parsons CM, Koelkebeck KW, Zimmerman RA, 2000. The effects of dietary available phosphorus levels and phytase on performance of young and older laying hens. Poul Sci 79: 224-230. https://doi.org/10.1093/ps/79.2.224
Brugalli I, Silva DJ, Albino LFT, Gomes PC, Rostagno HS, Silva MA, 1999. Available phosphorus requirement and effect of particle size on phosphorus bioavailability from meat and bone meal for broiler chicks. Rev Bras Zootec 28: 1288- 1296. https://doi.org/10.1590/S1516-35981999000600016
Buffington DE, Collazo-Arocho A, Canton GH, Pitt D, 1981. Black globe-humidity index (BGHI) as comfort equation for dairy cows. Am Soc Agric Eng 24: 711-714.
Bünzen S, Rostagno HS, Borsatto CG, Pessõa GBS, Carvalho TA, Messias RKG, 2008. Exigência de fósforo disponível para frangos de corte de 22 a 35 dias de idade, mantendo-se a relação Ca:Pdisp em 2:1. Conferência Apinco de Ciência e Tecnologia Avícola, Campinas (Brazil), May 27-29. pp: 45.
Cardoso A Jr, Rodrigues PB, Bertechini AG, Freitas RTF, Lima RR, Lima GFR, 2010. Levels of available phosphorus and calcium for broilers from 8 to 35 days of age fed rations containing phytase. Rev Bras Zootec 39: 1237-1245. https:// doi.org/10.1590/S1516-35982010000600011
Cobb Vantress, 2012. Cobb Broiler Management Guide. CobbVantress Inc, Siloam Springs, AR, USA. Crenshaw TD, Rortvedt LA, Hassen Z, 2011. Triennial Growth Symp: A novel pathway for vitamin D-mediated phosphate homeostasis: Implications for skeleton growth and mineralization. J Anim Sci 89: 1957-1964. https://doi. org/10.2527/jas.2010-3411
Cromwell GL, Stahly TS, Monegue HJ, 1989. Effect of source and level of copper on performance and liver copper, stores in weanling pigs. J Anim Sci 67: 2996-3002. https://doi. org/10.2527/jas1989.67112996x
Detmann E, Souza MA, Valadares Filho SC, Queiroz AC, Berchielli TT, Saliba EOS, Cabral LS, Pina DS, Ladeira MM, Azevedo JAG, 2012. Métodos para análise de alimentos, 1st ed. Suprema, Visconde do Rio Branco, Brazil. 214 pp
Ellis R, Morris ER, Philpot C, 1977. Quantitative determination of phytate in the presence of high inorganic phosphate. Anal Biochem 77: 536-539. https://doi.org/10.1016/0003- 2697(77)90269-X
Gomes PC, Gomes MFM, Lima GJMM, Bellaver C, 1993. Exigência de fósforo e sua disponibilidade nos fosfatos monoamônio e monocálcico para frangos de corte até 21 dias de idade. Rev Bras Zootec 22: 755-763.
Kill JL, Donzele JL, Cardoso EF, Haddade I, Rossoni MC, Haese D, Pires AF, Lima AL, 2008. Exigência de fósforo para frangos de corte de 7 a 28 dias de idade. Congresso Brasileiro de Zootecnia, João Pessoa (Brazil), May 26-30.
Lana SRV, Oliveira RFM, Donzele JL, Gomes PC, Vaz RGMV, Rezende WO, 2005. Requirements of dietary digestible lysine for broilers from 22 to 42 days old on thermoneutral environment. Rev Bras Zootec 34: 1614-1623. https://doi. org/10.1590/S1516-35982005000500023
Lima IL, 1995. Disponibilidade de fósforo e de flúor de alguns alimentos e exigência nutricional de fósforo para frangos de corte. Doctoral thesis. Univ. Federal de Viçosa, Viçosa, Brazil. 121 pp.
Maia APA, Antunes MVL, Campos PHRF, Souza MF, Balbino EM, Oliveira RFM, Donzele JL, 2009. Available phosphorus levels in diets for male broilers from 8 to 21 days of age kept in high temperature and thermoneutral environment. Congr Int de Zootecnia, Águas de Lindoia (Brazil), May 18-22.
Mello HHC, Gomes PC, Rostagno HS, Albino LFT, Rocha TC, Almeida RL, Calderano AA, 2012. Dietary requirements of available phosphorus in growing broiler chickens at a constant calcium:available phosphorus ratio. Rev Bras Zootec 41 (11): 2323-2328. https://doi.org/10.1590/S1516- 35982012001100004
Persia ME, Saylor WW, 2006. Effects of broilers strain, dietary nonphytate phosphorus, and phytase supplementation on chick performance and tibia ash. J Appl Poult Res 15: 72-81. https://doi.org/10.1093/japr/15.1.72
Puppo D, Cardoso EF, Kill JL, Haese D, Rossoni MC, Haddade I, 2008. Exigência de fósforo para frangos de corte dos 8 aos 21 dias de idade. V Congr Nordestino de Produção Animal, Aracaju (Brazil), Nov 24-27.
Qian H, Kornegay ET, Denbow DM, 1997. Utilization of phytate phosphorusand calcium as influenced by microbial phytase, cholecalciferol, and the calcium: total phosphorus ratio in broilers diets. Poult Sci 76: 37-46. https://doi.org/10.1093/ ps/76.1.37
Rama Rao SV, Ramasubba Reddy V, 2003. Relative bioavailability and utilization of phosphatic fertilizers as sources of phosphorus in broilers and layers. Br Poult Sci 44 (1): 96- 103. https://doi.org/10.1080/0007166031000085274
Rama Rao SV, Raju MVLN, Reddy MR, Pavani P, 2006. Interaction between dietary calcium and non-phytate phosphorus levels on growth, bone mineralization and mineral excretion in commercial broilers. Anim Feed Sci Technol 131: 133-148. https://doi.org/10.1016/j.anifeedsci.2006.02.011
Ronchi C, 2004. Principais práticas de manejo para aves recémnascidas. Revista AveWorld 6: 26-30.
Rostagno HS, Albino LFT, Donzele JL, Gomes PC, Oliveira RFM, Lopes DC, Soares AF, Barreto SLT, 2005. Brazilian tables for poultry and swine: composition of feedstuffs and nutritional requirements, 2nd ed. UFV, Viçosa, Brazil. 186 pp.
Rostagno HS, Albino LFT, Donzele JL, Gomes PC, Oliveira RFM, Lopes DC, Soares AF, Barreto SLT, Euclides RF, 2011. Brazilian tables for poultry and swine: composition of feedstuffs and nutritional requirements, 3rd ed. UFV, Viçosa, Brazil. 252 pp.
Runho RC, Gomes PC, Rostagno HS, Albino LFT, Lopes PS, Pozza PC, 2001. Available phosphorus requirement of male and female broilers from 1 to 21 days of age. Rev Bras Zootec 30: 187-196. https://doi.org/10.1590/S1516- 35982001000100027
Saraiva A, Donzele JL, Oliveira RF, Abreu ML, Silva FC, Guimarães SE, Kim SW, 2012. Phosphorus requirements for 60- to 100-kg pigs selected for high lean deposition under different thermal environments. J Anim Sci 90: 1499-1505. https://doi.org/10.2527/jas.2010-3623
SAS Inst, 2012. JMP Pro 10 Basic Analysis and Graphing. SAS Institute Inc., Cary, NC, USA.
Sethi PK, McMurtry JP, Pesti GM, Edwards HM Jr, Aggrey SE, 2008. Physiological responses to divergent selection for phytate phosphorus bioavailability in a randombred chicken population. Poult Sci 87: 2512-2516. https://doi.org/10.3382/ ps.2008-00190
Silva JHV, Araújo JA, Goulart CC, Costa FGP, Sakomura NK, Martins TDD, 2008. Calcium:available phosphorus ratio and phytase levels for semi heavy laying hens in the first and second posture cycle. Rev Bras Zootec 37 (12): 2166-2172. https://doi.org/10.1590/S1516-35982008001200013
Tamin NM, Angel R, Christman M, 2004. Influence of dietary calcium and phytase on phytate phosphorus hydrolyses in broiler chickens. Poult Sci 83 (8): 1358-1367. https://doi. org/10.1093/ps/83.8.1358
Valério SR, Oliveira RFM, Donzele JL, Albino LFT, Orlando UAD, Vaz RGMV, 2003. Digestible lysine levels in diets maintaining or not the relationship of amino acids for broilers from 1 to 21 days of age kept under heat stress. Rev Bras Zootec 32 (2): 361-371.
Xie M, Wang SX, Hou SS, Huang W, 2009. Interaction between dietary calcium and non-phytate phosphorus on growth performance and bone ash in early White Pekin ducklings. Anim Feed Sci Technol 151: 161-166. https://doi. org/10.1016/j.anifeedsci.2009.01.005
Yan F, Kersey JH, Fritts CA, Waldroup PW, Stilborn HL, Crum RC Jr, Rice DW, Raboy V, 2000. Evaluation of normal yellow dent corn and high available phosphorus corn in combination with reduced dietary phosphorus and phytase supplementation for broilers grown to market weights in litter pens. Poult Sci 79 (9): 1282-1289. https://doi.org/10.1093/ ps/79.9.1282










.jpg&w=3840&q=75)