Manage Wet Litter
Strategies to Manage Wet Litter
Published: September 21, 2010
By: Stephen R. Collett (The University of Georgia, College of Veterinary Medicine, USA)
Abstract
Poultry litter becomes wet when the rate of water addition (urine/faeces/spillage) exceeds the rate of removal (evaporation). Anti-nutritional factors, toxins, pathogens and nutrient imbalances may cause wet litter directly by altering normal digestive physiology or indirectly by disturbing normal gut ecology.
Poultry litter becomes wet when the rate of water addition (urine/faeces/spillage) exceeds the rate of removal (evaporation). Anti-nutritional factors, toxins, pathogens and nutrient imbalances may cause wet litter directly by altering normal digestive physiology or indirectly by disturbing normal gut ecology.
Poor quality ingredients and those with excess oligosaccharides or minerals can cause a nutritionally induced polydypsia, polyuria and diarrhoea which may increase water output sufficiently to cause wet litter. If the situation persists for long enough the ensuing inflammatory response causes mild to severe gastro enteritis which further increases water output. The damage so caused to the cytoskeleton of the gastrointestinal tract reduces the surface area for nutrient absorption and allows the opportunity for pathogen proliferation, gut colonization and even invasion and systemic disease.
It is necessary to use an integrated and holistic approach in manipulating the microbial populations and host inflammatory response to managing the gut ecology. Antibiotics, enzymes, drinking water acidification, probiotics, prebiotics, immune modulators and mycotoxin binders have all shown promise in this regard.
Seeding the gastro-intestinal tract with bacteria at hatch and subsequently managing the gastro-intestinal environment, whilst at the same time reducing anti-nutritional factors and the use of precision diet formulation has proved advantageous in minimising the incidence of wet litter in birds whilst maintaining and improving bird performance.
Introduction
Commercial poultry housing and management practices have been designed to keep birds within their comfort-zone at all times. Apart from satisfying the primary concern for bird welfare this also minimises homeostatic activity and ensures the most efficient partitioning of energy for production. Under these carefully controlled conditions water balance is kept positive (growth) or neutral.
Commercial poultry housing and management practices have been designed to keep birds within their comfort-zone at all times. Apart from satisfying the primary concern for bird welfare this also minimises homeostatic activity and ensures the most efficient partitioning of energy for production. Under these carefully controlled conditions water balance is kept positive (growth) or neutral.
Water balance is compromised in healthy animals when a dietary stress exceeds homeostatic mechanism capacity and in disease when the integrity and or function of the cells responsible for water/solute transport are adversely affected. If under such circumstances urine and faecal water loss increases to the point where the rate of water addition to the litter exceeds the rate of removal, the litter moisture content rises until it exceeds desirable levels (25%) and at this point it is deemed "wet".
Apart from being an indicator of gastrointestinal upset and feed conversion inefficiencies, wet litter also creates unfavourable house environment conditions.
Water Balance
Water movement across a biological membrane is solute concentration dependent and in living organisms; water balance is controlled by a combination of active and passive solute transfer, a process referred to as osmoregulation. Since water is the biological transport medium for nutrient absorption and waste excretion, water balance is governed by solute and water movement across the gastrointestinal and renal tubule cytoskeleton.
Water movement across a biological membrane is solute concentration dependent and in living organisms; water balance is controlled by a combination of active and passive solute transfer, a process referred to as osmoregulation. Since water is the biological transport medium for nutrient absorption and waste excretion, water balance is governed by solute and water movement across the gastrointestinal and renal tubule cytoskeleton.
Water balance is a crucial part of homeostasis and it involves equilibrating intake and synthesis (metabolic water) with excretion via the kidney (urine) and gastrointestinal tract (faeces) and insensible loss via the skin and respiratory tract (evaporation). Assuming house environment control is efficient and birds remain healthy, insensible (evaporative) water loss is minimised and excretory (urine and faecal) water loss is diet dependent. Dietary mineral content, anion-cation balance and several feed ingredient characteristics will affect water intake and feed passage time thus altering urine and faecal moisture.
Since feed is generally low in moisture (10%) and metabolic water production is limited by diet formulation (~ 0.14g/kcal of dietary energy) moisture intake is primarily controlled by drinking (~ 80%) . Water consumption is requirement-driven and the thirst centre is stimulated by cellular dehydration (osmoreceptors), extracellular dehydration (mechanoreceptors) and angiotensin II secretion (reninangiotensin axis).
Since feed is generally low in moisture (10%) and metabolic water production is limited by diet formulation (~ 0.14g/kcal of dietary energy) moisture intake is primarily controlled by drinking (~ 80%) . Water consumption is requirement-driven and the thirst centre is stimulated by cellular dehydration (osmoreceptors), extracellular dehydration (mechanoreceptors) and angiotensin II secretion (reninangiotensin axis).
Insensible moisture loss (skin and respiratory tract) accounts for 50-80% of total loss but seldom contributes directly to litter moisture since at thermoneutrality, evaporative loss is minimised and water in the vapour form is removed from the house relatively easily . Water loss as vapour does however increases relative humidity (RH) thus reducing the air's litter drying capacity and could cause saturation and condensation. Once condensed, water requires additional energy (heat of evaporation) and effort (air temperature/humidity control) to remove, so homeostatic stress that increases liquid water loss (faecal and urinary) therefore poses a greater risk to litter moisture control.
Urinary excretion is somewhat unique in the avian species since firstly the ureters open into the coprodeum and secondly the urine passes retrograde up the colon to the caecae before being evacuated via the cloaca with the faeces. The content of the urine is significantly altered during its passage through the coprodeum, colon and caecae.
Urine excretion is controlled by several hormones including arginine vasotocin (antidiuretic), renin/angiotensin (diuretic/antidiuretic and natiuretic/antinatiuretic), aldosterone (antinatiuretic), atrial natriuretic peptide (diuretic and naturietic), parathyroid hormone (calcium mobilization and excretion of phosphorus).
At standard temperatures broilers will consume approximately 1.75 to 2 times more water than feed (by weight) so it is crucial that the poultry house ventilation system design and operation is efficient enough to prevent the litter moisture content from exceeding an optimal 25%.. At stocking densities of 34kg/m2 huge amounts of water are added to the litter on a daily basis (Fig 1). At six weeks of age for example, 20,000 birds will excrete approximately 2.5t of water into the litter in one day. Relatively minor changes in water excretion rates can, very rapidly compromise litter moisture control.
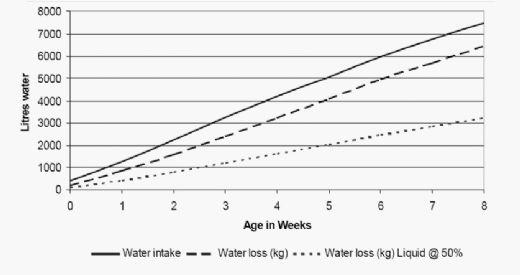
Figure 1. Increase in total water intake, total water loss and water loss as liquid (urine and faecal) for a 20.000 bird house (modified from Broiler performance Cobb 500; laughlin 2006; Goldstein and Skadhauge 2000, assuming Cobb drinking 1.75 times feed intake and water retention of 25% of weight gain).
Figure 1. Increase in total water intake, total water loss and water loss as liquid (urine and faecal) for a 20.000 bird house (modified from Broiler performance Cobb 500; laughlin 2006; Goldstein and Skadhauge 2000, assuming Cobb drinking 1.75 times feed intake and water retention of 25% of weight gain).
Water Imbalance
Urine output increases above normal (polyuria) when intake exceeds requirement (overcorrection of dehydration), the need for solute excretion exceeds normal (mineral and protein loading) or when urine concentrating mechanisms are compromised (nephrotoxins such as ochratoxin, citrinin and oosporin). Although an increase in urine output can cause wet litter, the condition is often incorrectly interpreted as an increase in true faecal water consequent to diarrhoea or enteritis.
Several feed ingredient characteristics can alter faecal water content directly by increasing ingesta osmolarity, or reducing transit time and absorptive surface area/function thus compromising water absorption and stimulating intake. The resultant increase in faecal water is termed diarrhoea. This is different to enteritis, where inflammation of the gastrointestinal lining negatively affects digestion and the consequent reduction in nutrient and water absorption cause faecal water to increases above normal. The latter is usually associated with pathological conditions involving gut microbiota or feed toxins while the former is usually physiological in nature. However any physiological perturbation that negatively affects nutrient assimilation (intake, digestion and absorption) will increase nutrient through-flow and could alter the microbial population of the distal gut sufficiently to cause wet litter. Ultimately, bird health and wellbeing, food safety and the efficient conversion of feed to food depends on the harmonious balance of the complex gastrointestinal tract ecology.
Urine output increases above normal (polyuria) when intake exceeds requirement (overcorrection of dehydration), the need for solute excretion exceeds normal (mineral and protein loading) or when urine concentrating mechanisms are compromised (nephrotoxins such as ochratoxin, citrinin and oosporin). Although an increase in urine output can cause wet litter, the condition is often incorrectly interpreted as an increase in true faecal water consequent to diarrhoea or enteritis.
Several feed ingredient characteristics can alter faecal water content directly by increasing ingesta osmolarity, or reducing transit time and absorptive surface area/function thus compromising water absorption and stimulating intake. The resultant increase in faecal water is termed diarrhoea. This is different to enteritis, where inflammation of the gastrointestinal lining negatively affects digestion and the consequent reduction in nutrient and water absorption cause faecal water to increases above normal. The latter is usually associated with pathological conditions involving gut microbiota or feed toxins while the former is usually physiological in nature. However any physiological perturbation that negatively affects nutrient assimilation (intake, digestion and absorption) will increase nutrient through-flow and could alter the microbial population of the distal gut sufficiently to cause wet litter. Ultimately, bird health and wellbeing, food safety and the efficient conversion of feed to food depends on the harmonious balance of the complex gastrointestinal tract ecology.
Strategies to Prevent Wet Litter
Preventing Polyuria
Chickens on grain based diets (approximately 30 meq Na+/kg) excrete a lot of NH4 +, phosphate and K+ while Na+ and Cl- together only account for just over 10% of the total osmolality. The relatively high levels of potassium in soybean (and molasses) can also be sufficient to induce a polydipsia, polyuria and wet litter.
Preventing Polyuria
Chickens on grain based diets (approximately 30 meq Na+/kg) excrete a lot of NH4 +, phosphate and K+ while Na+ and Cl- together only account for just over 10% of the total osmolality. The relatively high levels of potassium in soybean (and molasses) can also be sufficient to induce a polydipsia, polyuria and wet litter.
Most diets will have added salt and since sodium is the primary extracellular cation maintenance of sodium balance by the kidney is crucial to control of extracellular fluid volume and blood pressure . When the Na+ concentration in the ration is raised threefold there is a corresponding increase in urine Na+ and Cl- (<50% of osmotic space) at the expense of the NH4 + while other solute concentrations change very little . Minor sodium excess is controlled by reducing intestinal uptake but as the concentration in the diet increases renal naturesis follows . Elevated sodium excretion necessitates concomitant loss of an equivalent anion (usually Cl-) and water.
Provided the drinking water solute concentration does not exceed the regulatory capacity of the kidney the bird is able to cope with relatively high dietary salt concentrations by increasing urine output . The polyuria induced by sodium excretion is exacerbated by chloride induced osmotic diuresis and can to a degree be countered by partial replacement of salt derived sodium with sodium bicarbonate thereby reducing chlorine intake.
Dietary calcium and phosphorus levels are regulated by stringent maximum and minimum specification constraints because both the amount and ratio of these minerals is important to productivity . Calcium is reabsorbed from the glomerular filtrate very efficiently (>98%) by a mechanism that operates close to maximum while phosphorus is in contrast reabsorbed fairly inefficiently (40%) . Parathyroid hormone (PTH) secretion in response to low blood Ca enhances intestinal absorption and reduces renal excretion . Elevated blood Ca proportionally increases the calcium concentration of the glomerular filtrate which easily exceeds reabsoption capacity and consequently increases Ca excretion . PTH increases phosphorus excretion by inhibiting renal tubular reabsorption and secretion.
Excess calcium excretion can cause renal pathology (calcinosis/urolithiasis) resulting in compromised water retention and diuresis/wet litter . Dolomitic limestone contains relatively high levels of magnesium (8-10%) and apart from competing with calcium for absorption the Mg excretion can cause diuresis and wet litter.
Mycotoxicoses are difficult to quantify as there appears to be significant interaction and potentiation but nephrotoxins such as ochratoxin (especially type A), citrinin and oosporin can compromise renal function causing polyuria/polydypsia and wet litter . The avoidance of contaminated ingredients, the dilution of contaminated ingredients with non-contaminated ingredients or the addition of mycotoxin binders are all potential ways of limiting or preventing toxicity.
Gut Health management to prevent Diarrhoea and Enteritis
The integrity of the gastrointestinal absorptive membrane determines the efficiency of the assimilation process. Development and maintenance of the structure (anatomy) and function (physiology) has to be integrated with gut microbial community evolution since that is the primary determinant of gut ecological evolution. Detailed study of gut ecology with modern molecular techniques has helped elucidate details of the gastrointestinal environment that challenges previous dogma . Similarly gene sequencing of purported commensals and host cascade reaction modulation studies have opened our minds to the significance of the vast cellular communication process responsible for balancing this eco system . As this knowledge base expands, it becomes easier to design strategies to maintain gut health through science based gut ecology management.
The integrity of the gastrointestinal absorptive membrane determines the efficiency of the assimilation process. Development and maintenance of the structure (anatomy) and function (physiology) has to be integrated with gut microbial community evolution since that is the primary determinant of gut ecological evolution. Detailed study of gut ecology with modern molecular techniques has helped elucidate details of the gastrointestinal environment that challenges previous dogma . Similarly gene sequencing of purported commensals and host cascade reaction modulation studies have opened our minds to the significance of the vast cellular communication process responsible for balancing this eco system . As this knowledge base expands, it becomes easier to design strategies to maintain gut health through science based gut ecology management.
Colonization of the gut with pioneer bacteria species that are able to modulate gene expression in the host gut epithelia to assist in creating favourable conditions for the evolution of a stable climax (steady state) community provides a natural form of defence against pathogen challenge. The speed with which this "climax flora" develops appears to be important with respect to future/sustained resilience . With this objective in mind it is possible to identify several management opportunities to enhance gut health and bird productivity including; seeding the hatchling gut with favorable flora; early modification of the gut environment to promote climax flora development; pathogen exclusion (competitive and selective); immune modulation; and ingredient/nutrient management.
Seeding of the gut
It was demonstrated many years ago that a "mature" gut microbial community can reduce the prevalence of wet litter by making it more difficult for pathogens to infiltrate . Steps to control gut health in broilers should ideally start at the parent flock level because the first organisms to gain access to the receptive and uninhabited environment of the hatchling gut originate from the shell. Manipulation of parent gut flora can have a beneficial effect on offspring resistance to pathogen colonization because even low doses of beneficial bacteria can significantly improve resistance to pathogen colonization if introduced at hatch . These organisms also create conditions that shape development of the climax flora.
It was demonstrated many years ago that a "mature" gut microbial community can reduce the prevalence of wet litter by making it more difficult for pathogens to infiltrate . Steps to control gut health in broilers should ideally start at the parent flock level because the first organisms to gain access to the receptive and uninhabited environment of the hatchling gut originate from the shell. Manipulation of parent gut flora can have a beneficial effect on offspring resistance to pathogen colonization because even low doses of beneficial bacteria can significantly improve resistance to pathogen colonization if introduced at hatch . These organisms also create conditions that shape development of the climax flora.
Similarly, dosing day-old chicks with competitive exclusion products can reduce pathogen infection rate (95% to <5%) following low grade (<104CFU) challenge. The proviso being, that the process of gut colonization is allowed to proceed for at least 4 hours before challenge . Administration of Enterococcus faecium, Lactobacillus reuteri , a combination of Lactobacillus spp, B. bifidum, and Aspergillus oryzae , or a combination of Lactobacillus acidophilus, Bacillus subtilis and Enterococcus faecium significantly improved broiler performance under research conditions. Some companies have successfully utilized continuously fed probiotics to enhance gut health and feed efficiency on a commercial scale (Kounev, Z. 2004, personal communication).
Lactobacillus johnsonii and a mixture of Lactobacillus acidophilus and Streptococcus faecium have shown promise in reducing the impact of low grade necrotic enteritis on performance when used as probiotics . In one particular study a probiotic matched an antibiotic in protecting against necrotic enteritis induced mortality but the same organisms (Lactobacillus acidophilus and Enterococcus faecium) proved to be less successful in an earlier study.
It would appear that by selecting specific pioneer species as probiotic candidates it is possible to create a gut environment that accelerates the establishment of favourable and stable climax flora communities. With this strategy the emphasis subtly shifts from working against, to working with the natural ecology of the gut.
Gut environment management
Acidification
Meta analysis and literature review indicate that water and feed acidification have an important role to play in the avoidance of wet litter through gut flora management. Although a fairly well researched and accepted practice in pig production organic acids use is rapidly becoming popular in the poultry industry as a means of relinquishing dependence on antibiotics.
The beneficial effects of commercial acid preparations are thought to arise from the antibacterial properties of ionization. Organic acids are able to diffuse across the bacterial cell membrane rapidly when in the undissociated form. Once internalized the neutral pH of the cytoplasm causes dissociation, thus raising the intracellular concentration of both protons and anions. Bacterial proton-motive forces are exhausted in pursuance of homeostasis and the resultant rise in cytoplasm pH interferes with bacterial cell physiology. At low concentrations organic acids have a bacteriostatic effect but at high concentrations they become bactericidal (when acid concentration causes internal pH to rise to the point where denaturation of bacterial protein and DNA occurs.)
Acidification
Meta analysis and literature review indicate that water and feed acidification have an important role to play in the avoidance of wet litter through gut flora management. Although a fairly well researched and accepted practice in pig production organic acids use is rapidly becoming popular in the poultry industry as a means of relinquishing dependence on antibiotics.
The beneficial effects of commercial acid preparations are thought to arise from the antibacterial properties of ionization. Organic acids are able to diffuse across the bacterial cell membrane rapidly when in the undissociated form. Once internalized the neutral pH of the cytoplasm causes dissociation, thus raising the intracellular concentration of both protons and anions. Bacterial proton-motive forces are exhausted in pursuance of homeostasis and the resultant rise in cytoplasm pH interferes with bacterial cell physiology. At low concentrations organic acids have a bacteriostatic effect but at high concentrations they become bactericidal (when acid concentration causes internal pH to rise to the point where denaturation of bacterial protein and DNA occurs.)
Acid ionization varies considerably according to type, concentration and mix of acids used and is further modified by the pH, buffering capacity and water activity of the feed, water and gut content (Chung and Goeffert, 1970; Krause et al., 1994). Since their activity in the gastrointestinal tract is so variable, systematic studies of the effects of a variety of acidifiers are not available. It is not possible to determine the overall responses to complex acidification strategies or to compare these with other supplementation strategies using objective analytical process (Rosen, 2003).
Nutrient balance - Intake, absorption and excretion
The low pH of the upper gastrointestinal tract provides a competitive advantage to the acidophilic organisms and is by contrast, relatively hostile to many of the potential pathogens such as Clostridium perfringens and Salmonella sp. While the lower part of the digestive tract is alkaline (pH 7-8) and more hospitable to these potential pathogens their replication rate is presumably limited by nutrient availability. The flora of the lower gastrointestinal tract spend the majority of their existence in intense competition for a limited source of nutrients . Under such conditions evolution occurs very rapidly and continuously through mutation, selection and takeover. Any factor that reduces digestion efficiency in the upper gastrointestinal tract changes the nutrient supply to the lower tract and will likely favour specific stationary phase mutants. Since many of the problem organisms such as Clostridium perfringens are proteolytic, it is logical to expect that protein through-flow would provide a competitive advantage to these organisms and increase the propensity for enteritis and wet litter.
The low pH of the upper gastrointestinal tract provides a competitive advantage to the acidophilic organisms and is by contrast, relatively hostile to many of the potential pathogens such as Clostridium perfringens and Salmonella sp. While the lower part of the digestive tract is alkaline (pH 7-8) and more hospitable to these potential pathogens their replication rate is presumably limited by nutrient availability. The flora of the lower gastrointestinal tract spend the majority of their existence in intense competition for a limited source of nutrients . Under such conditions evolution occurs very rapidly and continuously through mutation, selection and takeover. Any factor that reduces digestion efficiency in the upper gastrointestinal tract changes the nutrient supply to the lower tract and will likely favour specific stationary phase mutants. Since many of the problem organisms such as Clostridium perfringens are proteolytic, it is logical to expect that protein through-flow would provide a competitive advantage to these organisms and increase the propensity for enteritis and wet litter.
While daily feed intake in broilers has increased tremendously, feed transit time in the small intestine has remained fairly constant . Since feed transit-time is inversely proportional to intake this means that retention time in the proventriculus and gizzard has declined. This is perhaps not surprising since the primary function of the proventriculus and gizzard (feed processing) is performed at the feed mill. What is perhaps significant is that the process of feed acidification and pepsin activation is likely also compromised.
Any factor that accelerates feed passage couldl potentially reduce the efficiency of digestion and absorption since this process requires time. Several feed ingredient characteristics will affect passage time including viscosity, particle size, digestibility (starch), and lipid or protein content. To prevent nutrient through-flow from causing wet litter the nutritionist should consider ingredient blend in addition to nutrient specification. Some ingredient characteristics such as water content, viscosity and non-digestible nutrient composition can be enhanced with concomitant enzyme and or osmolyte usage . Water soluble non starch polysaccharides (NSP) adversely affect digestibility by stimulating mucus production and increasing ingesta viscosity. Grains such as wheat, rye and barley are rich in water soluble NSPs and there is ample research to demonstrate that the use of exogenous enzymes improves digestibility . The toxin producing capability of Clostridium perfringens is also highly dependent on the form (mono vs disaccharide) carbohydrate substrate.
All fats and oils have the potential to become oxidised and the resulting rancid fats have compromised digestibility and can cause gastrointestinal disturbance and wet litter directly (steatorrhoea) or indirectly by affecting gut flora (oxidative). Unprotected fatty acids released by oil seed processing (grinding or chemical extraction) are very susceptible to oxidative rancidity.
All fats and oils have the potential to become oxidised and the resulting rancid fats have compromised digestibility and can cause gastrointestinal disturbance and wet litter directly (steatorrhoea) or indirectly by affecting gut flora (oxidative). Unprotected fatty acids released by oil seed processing (grinding or chemical extraction) are very susceptible to oxidative rancidity.
The growth enhancing effect of dietary enzymes is comparable to that of the antibiotics when tested in controlled experimental conditions, suggesting a common mechanism of action, i.e. manipulation of the gut ecology. This may be one of the explanations why there is individual and ingredient variation in response to enzyme supplementation. Apart from the direct feed efficiency implication of reduced digestion and absorption, the through flow of undigested nutrients impacts downstream gut ecology. Potentially toxic compounds such as ammonia, amines, phenols and indoles are generated by the proteolytic and ureolytic activity of the caecal flora on non-digested nutrients that make their way through to the caecal pouches. These toxic compounds affect flora ecology in the rabbit and the same is likely true for the broiler. The morphology of the ileocaecal junction is such that only fluid or very small (dissolved or suspended) particles enter the caecal pouch when intra luminal pressure increases during convergence of rectal retro peristaltic and ileal peristaltic contractions . Since the retro peristaltic contractions of the colon/rectum are almost continuous, 87-97% of the caecal fluid originates from the urine. Urine derived uric acid reaching the caeca by retro peristalsis and undigested protein from the upper gastrointestinal tract provide a source of nitrogen for microbial amino acid synthesis. Subsequent degradation of caecal microbial protein by for caecal microbial communities.
The amount of nitrogen reaching the caeca is influenced by the amount of protein in the diet, the efficiency of protein digestion /absorption in the upper gastrointestinal tract and the state of nitrogen balance. Exogenous enzymes added to the diet to promote protein digestion affect caecal flora communities by reducing the amount of protein nitrogen reaching the caeca. Conversely any physiological perturbation that negatively affects nutrient assimilation (intake, digestion and absorption) will increase caecal nitrogen either by increasing nutrient through flow or body protein turnover rate (nitrogen excretion via the urine).
Volatile fatty acids (VFA) are by products of uric acid degradation by caecal flora and despite passive absorption, caecal VFA concentrations (acetate>propionate>butyrate) are very high (125nM) . Since these weak organic acids have antibacterial activity they likely play an important role in balancing the caecal ecology.
Antimicrobials
Antibiotics have been an integral part of poultry feed for the past 50 years . Decades of research and field use have established the efficiency of antibiotics as growth promoters and in-feed antibiotics have been shown to subtly change the composition of the normal flora. PCR-denaturant gradient gel electrophoresis (DGGE) studies indicate that while antibiotics definitely change the gut flora profile, microbial populations become more homogeneous. DGGE studies are useful for tracking changes in gut flora profiles but give no detail on actual composition. The use of 16S clone libraries have made it possible to study specific organism profile changes in the gut. It would appear that lactobacilli and clostridia are the two groups most significantly affected by antibiotics.
Antibiotics have been an integral part of poultry feed for the past 50 years . Decades of research and field use have established the efficiency of antibiotics as growth promoters and in-feed antibiotics have been shown to subtly change the composition of the normal flora. PCR-denaturant gradient gel electrophoresis (DGGE) studies indicate that while antibiotics definitely change the gut flora profile, microbial populations become more homogeneous. DGGE studies are useful for tracking changes in gut flora profiles but give no detail on actual composition. The use of 16S clone libraries have made it possible to study specific organism profile changes in the gut. It would appear that lactobacilli and clostridia are the two groups most significantly affected by antibiotics.
Antibiotic growth promotion strategies have focused on manipulation of the gut ecology of the small intestine to improve feed efficiency but the impact on caecal flora, house flora and seeding of the hatchling gut of the next placement has been ignored. The gut flora changes elicited by a growth promotant are dependent on its antibacterial properties, rate of absorption, method of inactivation/metabolism, route of excretion, the degree of luminal (micro organism) enzyme inactivation or adsorption to ingesta. Many antibiotics are excreted via the urine in an active form, whether metabolized or not. Parenteral antimicrobials or those that are absorbed from the gastrointestinal tract after oral administration are concentrated in the urine and subsequently transported back to the caeca as illustrated by the high concentration of antibiotic in the caecal wall. Orally administered antimicrobials that or not absorbed also reach the caeca when the intraluminal pressure at the ileocaecal junction increases and liquid is forced into the caecal pouches.
The extensive reviews on in-feed antibiotic use and those covering the various alternatives, have reported on research investigating the response to first-time-one-off use of growth promoter strategies in controlled trials under carefully monitored experimental conditions. Broiler production is, in contrast, a continuous system. Broiler gut flora determines the composition of the litter/house flora which in turn acts as the seed stock for the gut flora of the next placement . While the small-intestine ecology influences the efficiency of digestion and absorption it is the caecal/colon/rectal flora that gives rise to the house flora. Although there are literally thousands of growth-promotant trials demonstrating their efficacy (or lack thereof) the literature is devoid of data showing the long-term effect of such programs. While the use of a growth promoter can alter the gut flora within a couple of weeks it takes several grow-out cycles to change the house flora . This is by no means a new concept, both rotation and shuttle programs have been used for decades to avoid the lack of response to growth promoting antibiotics following persistent use.
Just like penicillin many of the mycotoxins that commonly contaminate poultry feed likely have antimicrobial properties. Mycotoxin research has focused on host toxicity but it is possible that gut flora destabilization and feed efficiency is affected long before symptoms of toxicity appear. Molecular profiling using 16SrRNA library studies will likely provide more insight into the impact that mycotoxin contamination of feed has on gut flora and help with the understanding of exactly how these novel strategies help to control gut health.
Selective exclusion
Pathogen attachment to the intestinal epithelium is a pivotal first step in the colonisation of the gut and depends on, amongst other things, flagella, type 1 fimbriae and pillus receptors for specific host cell docking sites . Adherence has also been associated with mannose resistant haemagglutinins. Scanning electron microscope studies of the caecal epithelium have shown that the organisms of the gut flora form a tightly adherent mat over the gut surface. These organisms are attached to each other and the epithelia by a series of fibrils, which effectively prevents pathogenic organisms from gaining access to epithelial receptors. The adhesive flagella of enteropathogenic E. coli (EPEC) have been shown to be induced by animal cells. While competitive exclusion relies on the ability of live organisms to compete for attachment sites it is also possible to block attachment sites with decoy molecules and change gut flora communities
Pathogen attachment to the intestinal epithelium is a pivotal first step in the colonisation of the gut and depends on, amongst other things, flagella, type 1 fimbriae and pillus receptors for specific host cell docking sites . Adherence has also been associated with mannose resistant haemagglutinins. Scanning electron microscope studies of the caecal epithelium have shown that the organisms of the gut flora form a tightly adherent mat over the gut surface. These organisms are attached to each other and the epithelia by a series of fibrils, which effectively prevents pathogenic organisms from gaining access to epithelial receptors. The adhesive flagella of enteropathogenic E. coli (EPEC) have been shown to be induced by animal cells. While competitive exclusion relies on the ability of live organisms to compete for attachment sites it is also possible to block attachment sites with decoy molecules and change gut flora communities
Immune response
Any immune response bears a production cost. An appropriate immune response, adequate to contain infectious disease and minimize its impact on productivity, is the cost of health. An inappropriate, excessive or inadequate immune response will depress performance unnecessarily, so in a performance driven broiler industry the prevention of wet litter should include an immune modulation strategy.
Any immune response bears a production cost. An appropriate immune response, adequate to contain infectious disease and minimize its impact on productivity, is the cost of health. An inappropriate, excessive or inadequate immune response will depress performance unnecessarily, so in a performance driven broiler industry the prevention of wet litter should include an immune modulation strategy.
The gastrointestinal environment is loaded with a plethora of antigens of feed and micro-organism origin, the majority of which pose no threat of infectious disease. An inappropriate adaptive immune response to non-pathogen derived antigens is prevented by the innate immune system.
With systemic challenge, most (70%) of the negative impact on growth rate and feed efficiency is attributed to reduced feed intake, while the inefficiencies of catabolism and nutrient absorption account for the rest (30%) . Low level antigen recognition at the gut/ingesta interface probably seldom stimulates systemic/fever response but pro-inflammatory mediators released in response to antigen stimulation of this nature can damage host tissue, thereby causing localized inflammatory disease and reduced feed efficiency.
Antigen induced inflammation of the gut cytoskeleton stimulates an increase in mucus secretion, paracellular permeability, and feed passage (peristalsis). The cascade of events that follows is self perpetuating and provides additional advantage to organisms such as Clostridium perfringens that are capable of rapid multiplication thus increasing the propensity for wet litter . Both endogenous and exogenous anti-inflammatory agents modulate the immune response and help in the maintenance of gut health by preserving the integrity of the host enteron/environment interface and reducing the systemic (fever) response.
Local immune response is more pronounced in the lower gastrointestinal tract and high molecular weight proteins are more immunogenic so, apart from the obvious inefficiencies of nutrient wastage arising from poor digestibility or rapid feed passage, undigested proteins reaching the caeca are strongly inflammatory and thus further reduce feed efficiency. High protein diets, essential to attain broiler muscle tissue accretion rates, increase the risk of downstream gut health challenges by increasing the chance of protein throughflow. Peptic digestion is already marginal because selection for growth rate has reduced feed retention time in the crop and gizzard, thus reducing enzyme/nutrient contact time. This is especially so with soluble protein because liquids pass through the digestive tract 15% faster than solids.
The nature and extent of the inflammatory response is influenced by several nutritional factors. Dietary polyunsaturated fatty acids (PUFA) for example provide the building blocks for cell membrane synthesis and indirectly determine the type of immune response since the cell membrane lipids provide the substrate for immune system communication molecule synthesis . Cereal grains are high in linoleic acid (n-6 PUFA precursor for arachidonic acid) which generates prostaglandins, leukotreins and thromboxanes while fish oil is high in n-3 PUFA which generates Interleukin-1 and prostaglandin-E.
Just as the cost of an excessive or inappropriate immune response negatively impacts performance so too does an inadequate immune response. An inadequate immune response is usually recognized as an increase in flock mortality but has a negative economic impact long before flock mortality rises. Specific infectious diseases nutritional deficiencies, toxicity, and stress are all factors that can induce sufficient immune suppression to cause an inadequate response.
Stress only impacts performance measurably once the aggregate of each individual stress exceeds the host's coping mechanisms . The degree to which an adverse stimulus or stress will negatively impact bird performance is directly proportional to the existing stress load, so good animal husbandry, nutrition and bio-security are prerequisites to preventing wet litter.
Immune modulation can be used to carefully manage the balance between disease resistance and tolerance in order to maintain productivity.
Conclusion
Urine output increases above normal (polyuria) when intake exceeds requirement (overcorrection of dehydration), the need for solute excretion exceeds normal (mineral and protein loading) or when urine concentrating mechanisms are compromised. Polyuria can be avoided through careful diet formulation and ingredient management.
Urine output increases above normal (polyuria) when intake exceeds requirement (overcorrection of dehydration), the need for solute excretion exceeds normal (mineral and protein loading) or when urine concentrating mechanisms are compromised. Polyuria can be avoided through careful diet formulation and ingredient management.
Gut microbial imbalance is a fundamental cause of wet litter and there are several opportunities for intervention to enhance gut health and productivity by managing this ecosystem:
1. Seeding of the hatchling gut begins with vertical transmission of parent gut flora but is effectively modified with early administration of effective probiotics or competitive exclusion products. To be successful they must initiate the development of a primary flora which will rapidly evolve into a stable and favorable climax flora by creating suitable gut conditions and excluding unfavorable organisms.
2. Preparing the gut environment (pH, redox potential) for early transition from primary to climax flora through water/feed acidification. Candidates need to be weak acids that are buffered to withstand the neutralizing effect of minerals dissolved in the drinking water and have dissociation characteristics that make them active in the small intestine.
3. Excluding pathogens from colonizing the gut by competitive and selective exclusion. It is important that the selective exclusion product is compatible with (does not exclude) the organisms used for competitive exclusion or as a probiotic.
4. Enhancing resilience by stimulating protective immune response while suppressing the acute phase or fever response.
5. Decreasing nutrient through flow by enhancing nutrient digestion and absorption (exogenous enzyme addition and nutrient modification, feeding and lighting programs, careful use of antibiotics) to avoid caecal flora upset.
References
Akester, A. R., R. S. Anderson, et al. (1967). "A radiographic study of urine flow in the domestic fowl." Br Poult Sci 8(3): 209-12.
Almirall, M. and E. Esteve-Garcia (1994). "Rate of passage of barley diets with chromium oxide: influence of age and poultry strain and effect of beta-glucanase supplementation." Poult Sci 73(9): 1433-40.
Amann, R. I., W. Ludwig, et al. (1995). "Phylogenetic identification and in situ detection of individual microbial cells without cultivation." Microbiol Rev 59(1): 143-69.
Anadon, A., M. R. Martinez-Larranaga, et al. (1995). "Pharmacokinetics and residues of enrofloxacin in chickens." Am J Vet Res 56(4): 501-6.
Anderson, D., V. McCracken, et al. (2000). "Gut microbiology and growth-promoting antibiotics in swine." Nutritional Abstracts and Reviews Series B: Livestock Feeds and Feeding 70(2): 101-108.
Annison, E. F., K. J. Hill, et al. (1968). "Volatile fatty acids in the digestive tract of the fowl." Br J Nutr 22(2): 207-16.
Arnason, S. S., G. E. Rice, et al. (1986). "Plasma levels of arginine vasotocin, prolactin, aldosterone and corticosterone during prolonged dehydration in the domestic fowl: effect of dietary NaCl." J Comp Physiol [B] 156(3): 383-97.
Arnason, S. S. and E. Skadhauge (1991). "Steady-state sodium absorption and chloride secretion of colon and coprodeum, and plasma levels of osmoregulatory hormones in hens in relation to sodium intake." J Comp Physiol [B] 161(1): 1-14.
Austin, D. J., M. Kakehashi, et al. (1997). "The transmission dynamics of antibiotic-resistant bacteria: the relationship between resistance in commensal organisms and antibiotic consumption." Proc R Soc Lond B Biol Sci 264(1388): 1629-38.
Austin, D. J., K. G. Kristinsson, et al. (1999). "The relationship between the volume of antimicrobial consumption in human communities and the frequency of resistance." Proc Natl Acad Sci U S A 96(3): 1152-6.
Avellaneda, G., J. Lu, et al. (2003). The Impact Of Growth-Promoting Antibiotics On Total Poultry Microbiota As Well As Enterococcus Population Present On Poultry Carcass. Congress of the World Veterinary Poultry Association July Program and Abstracts, Poultry Disease Research Center, University of Georgia.
Barnes, E. M. (1977). "Ecological concepts of the anaerobic flora in the avian intestine." Am J Clin Nutr 30(11): 1793-8.
Barnes, E. M. and C. S. Impey (1970). "The isolation and properties of the predominant anaerobic bacteria in the caeca of chickens and turkeys." Br Poult Sci 11(4): 467-81.
Barnes, E. M., G. C. Mead, et al. (1972). "The intestinal flora of the chicken in the period 2 to 6 weeks of age, with particular reference to the anaerobic bacteria." Br Poult Sci 13(3): 311-26.
Bedford, M. (1996). "Interaction between ingested feed and the digestive system in poultry." Journal of Applied Poultry Research 5: 86-95.
Bedford, M. (2000). "Removal of antibiotic growth promoters from poultry diets: implications and strategies to minimise subsequent problems." World's Poultry Science Journal. Volume Number. 56( 347-365).
Bilgin, N., F. Claesens, et al. (1992). "Kinetic properties of Escherichia coli ribosomes with altered forms of S12." J Mol Biol 224(4): 1011-27.
Bjorkman, J., D. Hughes, et al. (1998). "Virulence of antibiotic-resistant Salmonella typhimurium." Proc Natl Acad Sci U S A 95(7): 3949-53.
Bjornhag, D. and I. Sperber (1977). "Transport of various food components through the digestive tract of turkeys, geese and guinea fowl." Swedish Journal of Agricultural Science 7: 57-66.
Bonhoeffer, S., M. Lipsitch, et al. (1997). "Evaluating treatment protocols to prevent antibiotic resistance." Proc Natl Acad Sci U S A 94(22): 12106-11.
Bouma, J. E. and R. E. Lenski (1988). "Evolution of a bacteria/plasmid association." Nature 335(6188): 351-2.
Branch, W. J. and J. H. Cummings (1978). "Comparison of radio-opaque pellets and chromium sesquioxide as inert markers in studies requiring accurate faecal collections." Gut 19(5): 371-6.
Brandtzaeg, P. (1989). "Overview of the mucosal immune system." Curr Top Microbiol Immunol 146: 13-25.
Brandtzaeg, P. E. (2002). "Current understanding of gastrointestinal immunoregulation and its relation to food allergy." Ann N Y Acad Sci 964: 13-45.
Cherrington, C. A., M. Hinton, et al. (1990). "Effect of short-chain organic acids on macromolecular synthesis in Escherichia coli." J Appl Bacteriol 68(1): 69-74.
Cherrington, C. A., M. Hinton, et al. (1991). "Organic acids: chemistry, antibacterial activity and practical applications." Adv Microb Physiol 32: 87-108.
Choct, M. and G. Annison (1992). "The inhibition of nutrient digestion by wheat pentosans." Br J Nutr 67(1): 123-32.
Clark, N. B. and L. L. Mok (1986). "Renal excretion in gull chicks: effect of parathyroid hormone and calcium loading." Am J Physiol 250(1 Pt 2): R41-50.
Classen, H. L. (1996). "Cereal grain starch and exogenous enzymes in poultry diets." Animal Feed Science Technology 62: 21-27.
Clench, M. H., V. M. Pineiro-Carrero, et al. (1989). "Migrating myoelectric complex demonstrated in four avian species." Am J Physiol 256(3 Pt 1): G598-603.
Collett, S. (2005). Strategies for Improving Gut Health in Commercial Operations. 3rd International Poultry Broiler Nutritionists Conference - Poultry Beyond 2010 Auckland, New Zealand.
Collett, S. and K. Dawson (2001). Alternatives to subtherapeutic antibiotics: What are the options? How effective are they?2nd International Poultry Broiler Nutritionist's Conference. Poultry beyond 2005, Sheraton Rotorua, New Zealand.
Collier, C. T., J. D. van der Klis, et al. (2003). "Effects of tylosin on bacterial mucolysis, Clostridium perfringens colonization, and intestinal barrier function in a chick model of necrotic enteritis." Antimicrob Agents Chemother 47(10): 3311-7.
DANMAP (2002). Use of antimicrobial agents and occurance of antimicrobial resistance in bacteria from food animals and humans in Denmark: ISSN 1600-2032.
Davidson, P. (2001). Chemical preservatives and natural antimicrobial compounds. Food Microbiology - Fundamentals and Frontiers. M. Doyle, L. Beuchat and T. Montville. Washington, DC, American Society for Microbiology: 593-627.
Denbow, D. (2000). Gastrointestinal Anatomy and Physiology. Sturkies Avian Physiology. G. Wihittow. New York, Academic Press: 299-325.
Duke, G. E. (1982). "Gastrointestinal motility and its regulation." Poult Sci 61(7): 1245-56. Elson, C. O. and Y. Cong (2002). "Understanding immune-microbial homeostasis in intestine." Immunol Res 26(1-3): 87-94.
Ferrando, C., P. Vergara, et al. (1987). "Study of the rate of passage of food with chromiummordanted plant cells in chickens (Gallus gallus)." Q J Exp Physiol 72(3): 251-9.
Finkel, S. E. and R. Kolter (1999). "Evolution of microbial diversity during prolonged starvation." Proc Natl Acad Sci U S A 96(7): 4023-7.
Finucane, M., K. A. Dawson, et al. (1999). "Effects of Mannanoligosaccharide and BMD on Gut Microflora of Turkey Poults." Poultry Sci. 78 (Suppl. 1): 77.
Fritsche, K. L., N. A. Cassity, et al. (1991). "Effect of dietary fat source on antibody production and lymphocyte proliferation in chickens." Poult Sci 70(3): 611-7.
Geyra, A., Z. Uni, et al. (2001). "Enterocyte dynamics and mucosal development in the posthatch chick." Poult Sci 80(6): 776-82.
Gidenne, T. (1997). "Caeco-colic digestion in the growing rabbit: impact of nutritional factors and related disturbances." Livstock Production Science 51(73-78).
Giron, J. A., A. G. Torres, et al. (2002). "The flagella of enteropathogenic Escherichia coli mediate adherence to epithelial cells." Mol Microbiol 44(2): 361-79.
Godwin, D. and J. H. Slater (1979). "The influence of the growth environment on the stability of a drug resistance plasmid in Escherichia coli K12." J Gen Microbiol 111(1): 201-10.
Goldstein, D. and E. Skadhauge (2000). Renal and extrarenal regulation of body fluid compartments. Sturkie's Avian Physiology, Academic press.
Gong, J., R. J. Forster, et al. (2002). "Diversity and phylogenetic analysis of bacteria in the mucosa of chicken ceca and comparison with bacteria in the cecal lumen." FEMS Microbiol Lett 208(1): 1-7.
Gray, D. A. (1993). "Plasma atrial natriuretic factor concentrations and renal actions in the domestic fowl." J Comp Physiol [B] 163(6): 519-23.
Gray, D. A., R. J. Naude, et al. (1988). "Plasma arginine vasotocin and angitensin II in the water deprived ostrich (Struthio camelus)." Comp. Biochem. Physiol. 89(2): 251-256.
Helling, R. B., T. Kinney, et al. (1981). "The maintenance of Plasmid-containing organisms in populations of Escherichia coli." J Gen Microbiol 123(Pt 1): 129-41.
Hooge, D. (2003). Broiler chicken performance may improve with MOS. Feedstuffs: 11-13.
Idris, U., J. I. Lu, et al. (2003). Factors Affecting Epidemiology Of Antibiotic-Resistant Campylobacter Jejuni And Campylobarcter Coli. Program and Abstracts, Congress of the World Veterinary Poultry Association.
Iji, P. (1999). "The impact of cereal non-starch polysaccharides on intestinal development and function in broiler chickens." World's Poultry Science Journal 55: 375-387.
Imhof, M. and C. Schlotterer (2001). "Fitness effects of advantageous mutations in evolving Escherichia coli populations." Proc Natl Acad Sci U S A 98(3): 1113-7.
Kanosue, K., H. Schmid, et al. (1990). "Differential osmoresponsiveness of periventricular neurons in duck hypothalamus." Am J Physiol 258(4 Pt 2): R973-81.
Kaufman, S., H. P. Kaesermann, et al. (1980). "The mechanism of drinking induced by parenteral hyperonocotic solutions in the pigeon and in the rat." J Physiol 301: 91-9.
Kellerup, S. U., J. E. Parker, et al. (1965). "Effects of restricted water consumption on broiler chicks." Poultry Science 44: 78-83.
Kelly, D., J. I. Campbell, et al. (2004). "Commensal anaerobic gut bacteria attenuate inflammation by regulating nuclear-cytoplasmic shuttling of PPAR-gamma and RelA." Nat Immunol 5(1): 104- 12.
Klasing, K. C. (1998). "Nutritional modulation of resistance to infectious diseases." Poult Sci 77(8): 1119-25.
Klipper, E., D. Sklan, et al. (2001). "Response, tolerance and ignorance following oral exposure to a single dietary protein antigen in Gallus domesticus." Vaccine 19(20-22): 2890-7.
Klipper, E., D. Sklan, et al. (2004). "Maternal antibodies block induction of oral tolerance in newly hatched chicks." Vaccine 22(3-4): 493-502.
Knarreborg, A., M. A. Simon, et al. (2002). "Effects of dietary fat source and subtherapeutic levels of antibiotic on the bacterial community in the ileum of broiler chickens at various ages." Appl Environ Microbiol 68(12): 5918-24.
Knoll, U., G. Glunder, et al. (1999). "Comparative study of the plasma pharmacokinetics and tissue concentrations of danofloxacin and enrofloxacin in broiler chickens." J Vet Pharmacol Ther 22(4): 239-46.
Kocher, A., R. Hughes, et al. (1997). Beta-gluconase reduces but does not eliminate variation in AME of barley varieties. Australian Poultry Science Symposium.
Koike, T. I., L. R. Pryor, et al. (1979). "Effects of salt infusion on plasma immunoreactive vasotocin in conscious chickens." Gen. Comp. Endocrinology 37: 451-458.
Korver, D. and K. Klasing (1995). "n-3 polyunsaturated fatty acids improve growth rate of broiler chickens and decrease interleukin-1 production." Poultry Science 74:(Supplement): 15.
Korver, D. R. and K. C. Klasing (1997). "Dietary fish oil alters specific and inflammatory immune responses in chicks." J Nutr 127(10): 2039-46.
Kubena, L. F., R. H. Bailey, et al. (2001). "Cecal volatile fatty acids and broiler chick susceptibility to Salmonella typhimurium colonization as affected by aflatoxins and T-2 toxin." Poult Sci 80(4): 411-7.
Lan, P. T., H. Hayashi, et al. (2002). "Phylogenetic analysis of cecal microbiota in chicken by the use of 16S rDNA clone libraries." Microbiol Immunol 46(6): 371-82.
Leeson, S. and J. D. Summers (2005). Commercial poultry nutrition. Canada, University books. Levin, B. R., V. Perrot, et al. (2000). "Compensatory mutations, antibiotic resistance and the population genetics of adaptive evolution in bacteria." Genetics 154(3): 985-97.
Levy, S. B. (2002). "The 2000 Garrod lecture. Factors impacting on the problem of antibiotic resistance." J Antimicrob Chemother 49(1): 25-30.
Lilijebjelke, K., C. Hofacre, et al. (2003). "Molecular Epidemiology of Salmonella on Poultry Farms In NE Georgia." Program and Abstracts, Congress of the World Veterinary Poultry Association. Lillehoj, H. S. and J. M. Trout (1996). "Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites." Clin Microbiol Rev 9(3): 349-60.
Lu, J., U. Idris, et al. (2003). "Diversity and succession of the intestinal bacterial community of the maturing broiler chicken." Appl Environ Microbiol 69(11): 6816-24.
Marder, J. and J. Ben-Asher (1983). "Cutaneous water evaporation--I. Its significance in heatstressed birds." Comp Biochem Physiol A 75(3): 425-31.
Marder, J. and P. Raber (1989). "Beta-adrenergic control of trans-cutaneous evaporative cooling mechanisms in birds." J. Comp. Physiol. 159: 97-103.
Massad, E., S. Lundberg, et al. (1993). "Modeling and simulating the evolution of resistance against antibiotics." Int J Biomed Comput 33(1): 65-81.
McFall-Ngai, M. J. (2002). "Unseen forces: the influence of bacteria on animal development." Dev Biol 242(1): 1-14.
Mongin (1981).
Morild, I., R. Monwinckel, et al. (1985). "The juxtaglomerular apparatus in the avian kidney." Cell Tissue Res. 240: 209-214.
Neish, A. S. (2002). "The gut microflora and intestinal epithelial cells: a continuing dialogue." Microbes Infect 4(3): 309-17.
Nguyen, T. N., Q. G. Phan, et al. (1989). "Effects of carriage and expression of the Tn10 tetracycline-resistance operon on the fitness of Escherichia coli K12." Mol Biol Evol 6(3): 213-25.
Oyofo, B. A., J. R. DeLoach, et al. (1989). "Prevention of Salmonella typhimurium colonization of broilers with D-mannose." Poult Sci 68(10): 1357-60.
Parks, C. W., J. L. Grimes, et al. (2001). "The effect of mannanoligosaccharides, bambermycins, and virginiamycin on performance of large white male market turkeys." Poult Sci 80(6): 718-23.
Refstie, S., B. Svihus, et al. (1999). "Nutrient digestibility in Atlantic salmon and broiler chickens related to viscosity and non-starch polysaccharide content in different soyabean products." Animal Feed Science Technology 79: 331-345.
Rice, G. E. and E. Skadhauge (1982). "Caecal water and electrolyte absorption and the effects of acetate and glusose, in dehydrated, low NaCl diet hens." J. Comp. Physiol. 147: 61-64.
Roberts, J. R. and M. R. Hughes (1983). "Glomerular filtration rate and drinking rate in Japanese quial, Coturnix coturnix japonica in response to acclimation to saline drinking water." Canadian Journal of Zoology 61: 2394.
Rosen, G. (2000). "Enzymes for broilers: A multi-factorial assessment." Fd. Intl. 21(12): 14-18.
Rosen, G. (2000). Multi-factorial assessment of exogenous enzymes in broiler pronutrition: Target and problems. Proceedings of the 3rd European Symposium on Feed Enzymes, Noordwijkerhout, Netherlands.
Rosen, G., Vol. II, p141. (1996). Proceedings of the World's Poultry Science Society, New Delhi.
Rosen, G. D. (1995). "Antibacterials in poultry and pig nutrition. In Biotechnology in the Animal Feeds and Animal Feeding Ed by R.J. Wallace and A. Chesson. VCH Verlagsgesellschaft mbH D-69461 Weinheim." 8: 143-172.
Rosen, G. D. (2001). "Multi-factorial efficacy evaluation of alternatives to antimicrobials in pronutrition." British Poultry Science 42(S1): S104-S105.
Salanitro, J. P., I. G. Blake, et al. (1978). "Bacteria isolated from the duodenum, ileum, and cecum of young chicks." Appl Environ Microbiol 35(4): 782-90.
Salanitro, J. P., I. G. Blake, et al. (1974). "Studies on the cecal microflora of commercial broiler chickens." Appl Microbiol 28(3): 439-47.
Schildknect, E., L. Rakebrand, et al. (2003). Changes In Anticoccidial Sensitivity Profiles Of Coccidia From Broiler Chickens Raised On Built-Up Litter For Eight Production Cycles Following A Coccidiosis Challenge. International Poultry Scientific Forum Abstracts.
Schildknect, E., L. Rakebrand, et al. (2003). Changes In Live Performance Of Broiler Chickens Raised On Built-Up Litter For Eight Production Cycles Following A Coccidiosis. International Poultry Scientific Forum Abstracts, Atlanta, Georgia.
Schrag, S. J. and V. Perrot (1996). "Reducing antibiotic resistance." Nature 381(6578): 120-1.
Schrag, S. J., V. Perrot, et al. (1997). "Adaptation to the fitness costs of antibiotic resistance in Escherichia coli." Proc R Soc Lond B Biol Sci 264(1386): 1287-91.
Schwalbe, R. S., C. W. Hoge, et al. (1990). "In vivo selection for transmissible drug resistance in Salmonella typhi during antimicrobial therapy." Antimicrob Agents Chemother 34(1): 161-3.
Schwalbe, R. S., W. J. Ritz, et al. (1990). "Selection for vancomycin resistance in clinical isolates of Staphylococcus haemolyticus." J Infect Dis 161(1): 45-51.
Sell, J., J. Eastwood, et al. (1983). "Influence of supplemental fat on metabolizable energy and ingesta transit time in laying hens." Nutrition Rep. Intern. 28: 487-495.
Shane, S. M., R. G. Young, et al. (1969). "Renal and parathyroid changes produced by high calcium intake in growing pullets." Avian Diseases 13: 558-567.
Sharon, N. and H. Lis (1993). "Carbohydrates in cell recognition." Sci Am 268(1): 82-9.
Sibbald, I. R. (1979). "Passage of feed through the adult rooster." Poult Sci 58(2): 446-59.
Sklan, D., D. Dubrov, et al. (1975). "51Cr-EDTA, 91Y and 141Ce as nonabsorbed reference substances in the gastrointestinal tract of the chicken." J Nutr 105(12): 1549-52.
Sklan, D. and S. Hurwitz (1980). "Protein digestion and absorption in young chicks and turkeys." J Nutr 110(1): 139-44.
Spring, P. (1996). Effects of mannanoligosaccharide on different caecal parameters and on the attachment of enteric pathogens in poultry. Zurich, Swiss Fed. Inst. Technology.
Stappenbeck, T. S., L. V. Hooper, et al. (2002). "Developmental regulation of intestinal angiogenesis by indigenous microbes via Paneth cells." Proc Natl Acad Sci U S A 99(24): 15451- 5.
Stavric, S., T. Gleeson, et al. (1987). "Effect of environmental temperature on the susceptibility of young chickens to Salmonella typhimurium." Australian Veterinary Journal 55: 413.
Stewart, F. M., R. Antia, et al. (1998). "The population genetics of antibiotic resistance. II: Analytic theory for sustained populations of bacteria in a community of hosts." Theor Popul Biol 53(2): 152-65.
Sudo, S. and G. Duke (1980). "Kinetics of absorption of volatile fatty acids from the ceca of domestic turkeys. Comp. Biochem. Physiol." 67: 231-237.
Sullivan, A., C. Edlund, et al. (2001). "Effect of antimicrobial agents on the ecological balance of human microflora." Lancet Infect Dis 1(2): 101-14.
Swamy, H. V., T. K. Smith, et al. (2002). "Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on production and metabolism in broilers." Poult Sci 81(7): 966-75.
Swamy, H. V., T. K. Smith, et al. (2002). "Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on swine performance, brain regional neurochemistry, and serum chemistry and the efficacy of a polymeric glucomannan mycotoxin adsorbent." J Anim Sci 80(12): 3257-67.
Tahara, T., S. Yoshioka, et al. (1997). "Isolation and partial characterization of bacteriocins produced by Lactobacillus gasseri JCM 2124." FEMS Microbiol Lett 148(1): 97-100.
Takei, Y., Y. Okawara, et al. (1988). "Water intake induced by water deprivation in the quail, Coturnix coturnix japonica." J Comp Physiol [B] 158(5): 519-25.
Turkov, M. I., V. V. Belushkin, et al. (1990). "[The effect of small concentrations of antibiotics blocking the synthesis of bacterial cell wall on the permeability of Escherichia coli cell wall for plasmid DNA]." Genetika 26(3): 443-7.
Uden, P., P. E. Colucci, et al. (1980). "Investigation of chromium, cerium and cobalt as markers in digesta. Rate of passage studies." J Sci Food Agric 31(7): 625-32.
Ueda, Y., S. Hachimura, et al. (2001). "Apoptosis of antigen-specific T cells induced by oral administration of antigen: comparison of intestinal and non-intestinal immune organs." Biosci Biotechnol Biochem 65(5): 1170-4.
Uni, Z., A. Geyra, et al. (2000). "Small intestinal development in the young chick: crypt formation and enterocyte proliferation and migration." Br Poult Sci 41(5): 544-51.
Uni, Z., Y. Noy, et al. (1999). "Posthatch development of small intestinal function in the poult." Poult Sci 78(2): 215-22.
Vaughan, E. E., F. Schut, et al. (2000). "A molecular view of the intestinal ecosystem." Curr Issues Intest Microbiol 1(1): 1-12.
Vergara, P., C. Ferrando, et al. (1989). "Factors determining gastrointestinal transit time of several markers in the domestic fowl." Q J Exp Physiol 74(6): 867-74.
Washburn, K. W. (1991). "Efficiency of feed utilization and rate of feed passage through the digestive system." Poult Sci 70(3): 447-52.
Weurding, R. E., A. Veldman, et al. (2001). "Starch digestion rate in the small intestine of broiler chickens differs among feedstuffs." J Nutr 131(9): 2329-35.
Wideman, R. F. J. (1987). "Renal regulation of avian calcium and phosphorus metabolism." Jurnal of Nutrition 117: 808-814.
Wideman, R. F. J., J. A. Closser, et al. (1985). "Urolithiasis in pullets and laying hens: role of dietary calcium and phosphorus " Poult Sci 64(2300-2307).
Wise, R. (1999). "A review of the mechanisms of action and resistance of antimicrobial agents." Can Respir J 6 Suppl A: 20A-2A.
Woo, P. C., A. P. To, et al. (2003). "Facilitation of horizontal transfer of antimicrobial resistance by transformation of antibiotic-induced cell-wall-deficient bacteria." Med Hypotheses 61(4): 503-8.
Xu, J., M. K. Bjursell, et al. (2003). "A genomic view of the human-Bacteroides thetaiotaomicron symbiosis." Science 299(5615): 2074-6.
Xu, J. and J. I. Gordon (2003). "Inaugural Article: Honor thy symbionts." Proc Natl Acad Sci U S A 100(18): 10452-9.
Zambrano, M. M., D. A. Siegele, et al. (1993). "Microbial competition: Escherichia coli mutants that take over stationary phase cultures." Science 259(5102): 1757-60.
Zhu, W. M., W. Liu, et al. (2000). "Isolation and characterization of a new bacteriocin from Lactobacillus gasseri KT7." J Appl Microbiol 88(5): 877-86.
Zinser, E. R. and R. Kolter (2004). "Escherichia coli evolution during stationary phase." Res Microbiol 155(5): 328-36.
Akester, A. R., R. S. Anderson, et al. (1967). "A radiographic study of urine flow in the domestic fowl." Br Poult Sci 8(3): 209-12.
Almirall, M. and E. Esteve-Garcia (1994). "Rate of passage of barley diets with chromium oxide: influence of age and poultry strain and effect of beta-glucanase supplementation." Poult Sci 73(9): 1433-40.
Amann, R. I., W. Ludwig, et al. (1995). "Phylogenetic identification and in situ detection of individual microbial cells without cultivation." Microbiol Rev 59(1): 143-69.
Anadon, A., M. R. Martinez-Larranaga, et al. (1995). "Pharmacokinetics and residues of enrofloxacin in chickens." Am J Vet Res 56(4): 501-6.
Anderson, D., V. McCracken, et al. (2000). "Gut microbiology and growth-promoting antibiotics in swine." Nutritional Abstracts and Reviews Series B: Livestock Feeds and Feeding 70(2): 101-108.
Annison, E. F., K. J. Hill, et al. (1968). "Volatile fatty acids in the digestive tract of the fowl." Br J Nutr 22(2): 207-16.
Arnason, S. S., G. E. Rice, et al. (1986). "Plasma levels of arginine vasotocin, prolactin, aldosterone and corticosterone during prolonged dehydration in the domestic fowl: effect of dietary NaCl." J Comp Physiol [B] 156(3): 383-97.
Arnason, S. S. and E. Skadhauge (1991). "Steady-state sodium absorption and chloride secretion of colon and coprodeum, and plasma levels of osmoregulatory hormones in hens in relation to sodium intake." J Comp Physiol [B] 161(1): 1-14.
Austin, D. J., M. Kakehashi, et al. (1997). "The transmission dynamics of antibiotic-resistant bacteria: the relationship between resistance in commensal organisms and antibiotic consumption." Proc R Soc Lond B Biol Sci 264(1388): 1629-38.
Austin, D. J., K. G. Kristinsson, et al. (1999). "The relationship between the volume of antimicrobial consumption in human communities and the frequency of resistance." Proc Natl Acad Sci U S A 96(3): 1152-6.
Avellaneda, G., J. Lu, et al. (2003). The Impact Of Growth-Promoting Antibiotics On Total Poultry Microbiota As Well As Enterococcus Population Present On Poultry Carcass. Congress of the World Veterinary Poultry Association July Program and Abstracts, Poultry Disease Research Center, University of Georgia.
Barnes, E. M. (1977). "Ecological concepts of the anaerobic flora in the avian intestine." Am J Clin Nutr 30(11): 1793-8.
Barnes, E. M. and C. S. Impey (1970). "The isolation and properties of the predominant anaerobic bacteria in the caeca of chickens and turkeys." Br Poult Sci 11(4): 467-81.
Barnes, E. M., G. C. Mead, et al. (1972). "The intestinal flora of the chicken in the period 2 to 6 weeks of age, with particular reference to the anaerobic bacteria." Br Poult Sci 13(3): 311-26.
Bedford, M. (1996). "Interaction between ingested feed and the digestive system in poultry." Journal of Applied Poultry Research 5: 86-95.
Bedford, M. (2000). "Removal of antibiotic growth promoters from poultry diets: implications and strategies to minimise subsequent problems." World's Poultry Science Journal. Volume Number. 56( 347-365).
Bilgin, N., F. Claesens, et al. (1992). "Kinetic properties of Escherichia coli ribosomes with altered forms of S12." J Mol Biol 224(4): 1011-27.
Bjorkman, J., D. Hughes, et al. (1998). "Virulence of antibiotic-resistant Salmonella typhimurium." Proc Natl Acad Sci U S A 95(7): 3949-53.
Bjornhag, D. and I. Sperber (1977). "Transport of various food components through the digestive tract of turkeys, geese and guinea fowl." Swedish Journal of Agricultural Science 7: 57-66.
Bonhoeffer, S., M. Lipsitch, et al. (1997). "Evaluating treatment protocols to prevent antibiotic resistance." Proc Natl Acad Sci U S A 94(22): 12106-11.
Bouma, J. E. and R. E. Lenski (1988). "Evolution of a bacteria/plasmid association." Nature 335(6188): 351-2.
Branch, W. J. and J. H. Cummings (1978). "Comparison of radio-opaque pellets and chromium sesquioxide as inert markers in studies requiring accurate faecal collections." Gut 19(5): 371-6.
Brandtzaeg, P. (1989). "Overview of the mucosal immune system." Curr Top Microbiol Immunol 146: 13-25.
Brandtzaeg, P. E. (2002). "Current understanding of gastrointestinal immunoregulation and its relation to food allergy." Ann N Y Acad Sci 964: 13-45.
Cherrington, C. A., M. Hinton, et al. (1990). "Effect of short-chain organic acids on macromolecular synthesis in Escherichia coli." J Appl Bacteriol 68(1): 69-74.
Cherrington, C. A., M. Hinton, et al. (1991). "Organic acids: chemistry, antibacterial activity and practical applications." Adv Microb Physiol 32: 87-108.
Choct, M. and G. Annison (1992). "The inhibition of nutrient digestion by wheat pentosans." Br J Nutr 67(1): 123-32.
Clark, N. B. and L. L. Mok (1986). "Renal excretion in gull chicks: effect of parathyroid hormone and calcium loading." Am J Physiol 250(1 Pt 2): R41-50.
Classen, H. L. (1996). "Cereal grain starch and exogenous enzymes in poultry diets." Animal Feed Science Technology 62: 21-27.
Clench, M. H., V. M. Pineiro-Carrero, et al. (1989). "Migrating myoelectric complex demonstrated in four avian species." Am J Physiol 256(3 Pt 1): G598-603.
Collett, S. (2005). Strategies for Improving Gut Health in Commercial Operations. 3rd International Poultry Broiler Nutritionists Conference - Poultry Beyond 2010 Auckland, New Zealand.
Collett, S. and K. Dawson (2001). Alternatives to subtherapeutic antibiotics: What are the options? How effective are they?2nd International Poultry Broiler Nutritionist's Conference. Poultry beyond 2005, Sheraton Rotorua, New Zealand.
Collier, C. T., J. D. van der Klis, et al. (2003). "Effects of tylosin on bacterial mucolysis, Clostridium perfringens colonization, and intestinal barrier function in a chick model of necrotic enteritis." Antimicrob Agents Chemother 47(10): 3311-7.
DANMAP (2002). Use of antimicrobial agents and occurance of antimicrobial resistance in bacteria from food animals and humans in Denmark: ISSN 1600-2032.
Davidson, P. (2001). Chemical preservatives and natural antimicrobial compounds. Food Microbiology - Fundamentals and Frontiers. M. Doyle, L. Beuchat and T. Montville. Washington, DC, American Society for Microbiology: 593-627.
Denbow, D. (2000). Gastrointestinal Anatomy and Physiology. Sturkies Avian Physiology. G. Wihittow. New York, Academic Press: 299-325.
Duke, G. E. (1982). "Gastrointestinal motility and its regulation." Poult Sci 61(7): 1245-56. Elson, C. O. and Y. Cong (2002). "Understanding immune-microbial homeostasis in intestine." Immunol Res 26(1-3): 87-94.
Ferrando, C., P. Vergara, et al. (1987). "Study of the rate of passage of food with chromiummordanted plant cells in chickens (Gallus gallus)." Q J Exp Physiol 72(3): 251-9.
Finkel, S. E. and R. Kolter (1999). "Evolution of microbial diversity during prolonged starvation." Proc Natl Acad Sci U S A 96(7): 4023-7.
Finucane, M., K. A. Dawson, et al. (1999). "Effects of Mannanoligosaccharide and BMD on Gut Microflora of Turkey Poults." Poultry Sci. 78 (Suppl. 1): 77.
Fritsche, K. L., N. A. Cassity, et al. (1991). "Effect of dietary fat source on antibody production and lymphocyte proliferation in chickens." Poult Sci 70(3): 611-7.
Geyra, A., Z. Uni, et al. (2001). "Enterocyte dynamics and mucosal development in the posthatch chick." Poult Sci 80(6): 776-82.
Gidenne, T. (1997). "Caeco-colic digestion in the growing rabbit: impact of nutritional factors and related disturbances." Livstock Production Science 51(73-78).
Giron, J. A., A. G. Torres, et al. (2002). "The flagella of enteropathogenic Escherichia coli mediate adherence to epithelial cells." Mol Microbiol 44(2): 361-79.
Godwin, D. and J. H. Slater (1979). "The influence of the growth environment on the stability of a drug resistance plasmid in Escherichia coli K12." J Gen Microbiol 111(1): 201-10.
Goldstein, D. and E. Skadhauge (2000). Renal and extrarenal regulation of body fluid compartments. Sturkie's Avian Physiology, Academic press.
Gong, J., R. J. Forster, et al. (2002). "Diversity and phylogenetic analysis of bacteria in the mucosa of chicken ceca and comparison with bacteria in the cecal lumen." FEMS Microbiol Lett 208(1): 1-7.
Gray, D. A. (1993). "Plasma atrial natriuretic factor concentrations and renal actions in the domestic fowl." J Comp Physiol [B] 163(6): 519-23.
Gray, D. A., R. J. Naude, et al. (1988). "Plasma arginine vasotocin and angitensin II in the water deprived ostrich (Struthio camelus)." Comp. Biochem. Physiol. 89(2): 251-256.
Helling, R. B., T. Kinney, et al. (1981). "The maintenance of Plasmid-containing organisms in populations of Escherichia coli." J Gen Microbiol 123(Pt 1): 129-41.
Hooge, D. (2003). Broiler chicken performance may improve with MOS. Feedstuffs: 11-13.
Idris, U., J. I. Lu, et al. (2003). Factors Affecting Epidemiology Of Antibiotic-Resistant Campylobacter Jejuni And Campylobarcter Coli. Program and Abstracts, Congress of the World Veterinary Poultry Association.
Iji, P. (1999). "The impact of cereal non-starch polysaccharides on intestinal development and function in broiler chickens." World's Poultry Science Journal 55: 375-387.
Imhof, M. and C. Schlotterer (2001). "Fitness effects of advantageous mutations in evolving Escherichia coli populations." Proc Natl Acad Sci U S A 98(3): 1113-7.
Kanosue, K., H. Schmid, et al. (1990). "Differential osmoresponsiveness of periventricular neurons in duck hypothalamus." Am J Physiol 258(4 Pt 2): R973-81.
Kaufman, S., H. P. Kaesermann, et al. (1980). "The mechanism of drinking induced by parenteral hyperonocotic solutions in the pigeon and in the rat." J Physiol 301: 91-9.
Kellerup, S. U., J. E. Parker, et al. (1965). "Effects of restricted water consumption on broiler chicks." Poultry Science 44: 78-83.
Kelly, D., J. I. Campbell, et al. (2004). "Commensal anaerobic gut bacteria attenuate inflammation by regulating nuclear-cytoplasmic shuttling of PPAR-gamma and RelA." Nat Immunol 5(1): 104- 12.
Klasing, K. C. (1998). "Nutritional modulation of resistance to infectious diseases." Poult Sci 77(8): 1119-25.
Klipper, E., D. Sklan, et al. (2001). "Response, tolerance and ignorance following oral exposure to a single dietary protein antigen in Gallus domesticus." Vaccine 19(20-22): 2890-7.
Klipper, E., D. Sklan, et al. (2004). "Maternal antibodies block induction of oral tolerance in newly hatched chicks." Vaccine 22(3-4): 493-502.
Knarreborg, A., M. A. Simon, et al. (2002). "Effects of dietary fat source and subtherapeutic levels of antibiotic on the bacterial community in the ileum of broiler chickens at various ages." Appl Environ Microbiol 68(12): 5918-24.
Knoll, U., G. Glunder, et al. (1999). "Comparative study of the plasma pharmacokinetics and tissue concentrations of danofloxacin and enrofloxacin in broiler chickens." J Vet Pharmacol Ther 22(4): 239-46.
Kocher, A., R. Hughes, et al. (1997). Beta-gluconase reduces but does not eliminate variation in AME of barley varieties. Australian Poultry Science Symposium.
Koike, T. I., L. R. Pryor, et al. (1979). "Effects of salt infusion on plasma immunoreactive vasotocin in conscious chickens." Gen. Comp. Endocrinology 37: 451-458.
Korver, D. and K. Klasing (1995). "n-3 polyunsaturated fatty acids improve growth rate of broiler chickens and decrease interleukin-1 production." Poultry Science 74:(Supplement): 15.
Korver, D. R. and K. C. Klasing (1997). "Dietary fish oil alters specific and inflammatory immune responses in chicks." J Nutr 127(10): 2039-46.
Kubena, L. F., R. H. Bailey, et al. (2001). "Cecal volatile fatty acids and broiler chick susceptibility to Salmonella typhimurium colonization as affected by aflatoxins and T-2 toxin." Poult Sci 80(4): 411-7.
Lan, P. T., H. Hayashi, et al. (2002). "Phylogenetic analysis of cecal microbiota in chicken by the use of 16S rDNA clone libraries." Microbiol Immunol 46(6): 371-82.
Leeson, S. and J. D. Summers (2005). Commercial poultry nutrition. Canada, University books. Levin, B. R., V. Perrot, et al. (2000). "Compensatory mutations, antibiotic resistance and the population genetics of adaptive evolution in bacteria." Genetics 154(3): 985-97.
Levy, S. B. (2002). "The 2000 Garrod lecture. Factors impacting on the problem of antibiotic resistance." J Antimicrob Chemother 49(1): 25-30.
Lilijebjelke, K., C. Hofacre, et al. (2003). "Molecular Epidemiology of Salmonella on Poultry Farms In NE Georgia." Program and Abstracts, Congress of the World Veterinary Poultry Association. Lillehoj, H. S. and J. M. Trout (1996). "Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites." Clin Microbiol Rev 9(3): 349-60.
Lu, J., U. Idris, et al. (2003). "Diversity and succession of the intestinal bacterial community of the maturing broiler chicken." Appl Environ Microbiol 69(11): 6816-24.
Marder, J. and J. Ben-Asher (1983). "Cutaneous water evaporation--I. Its significance in heatstressed birds." Comp Biochem Physiol A 75(3): 425-31.
Marder, J. and P. Raber (1989). "Beta-adrenergic control of trans-cutaneous evaporative cooling mechanisms in birds." J. Comp. Physiol. 159: 97-103.
Massad, E., S. Lundberg, et al. (1993). "Modeling and simulating the evolution of resistance against antibiotics." Int J Biomed Comput 33(1): 65-81.
McFall-Ngai, M. J. (2002). "Unseen forces: the influence of bacteria on animal development." Dev Biol 242(1): 1-14.
Mongin (1981).
Morild, I., R. Monwinckel, et al. (1985). "The juxtaglomerular apparatus in the avian kidney." Cell Tissue Res. 240: 209-214.
Neish, A. S. (2002). "The gut microflora and intestinal epithelial cells: a continuing dialogue." Microbes Infect 4(3): 309-17.
Nguyen, T. N., Q. G. Phan, et al. (1989). "Effects of carriage and expression of the Tn10 tetracycline-resistance operon on the fitness of Escherichia coli K12." Mol Biol Evol 6(3): 213-25.
Oyofo, B. A., J. R. DeLoach, et al. (1989). "Prevention of Salmonella typhimurium colonization of broilers with D-mannose." Poult Sci 68(10): 1357-60.
Parks, C. W., J. L. Grimes, et al. (2001). "The effect of mannanoligosaccharides, bambermycins, and virginiamycin on performance of large white male market turkeys." Poult Sci 80(6): 718-23.
Refstie, S., B. Svihus, et al. (1999). "Nutrient digestibility in Atlantic salmon and broiler chickens related to viscosity and non-starch polysaccharide content in different soyabean products." Animal Feed Science Technology 79: 331-345.
Rice, G. E. and E. Skadhauge (1982). "Caecal water and electrolyte absorption and the effects of acetate and glusose, in dehydrated, low NaCl diet hens." J. Comp. Physiol. 147: 61-64.
Roberts, J. R. and M. R. Hughes (1983). "Glomerular filtration rate and drinking rate in Japanese quial, Coturnix coturnix japonica in response to acclimation to saline drinking water." Canadian Journal of Zoology 61: 2394.
Rosen, G. (2000). "Enzymes for broilers: A multi-factorial assessment." Fd. Intl. 21(12): 14-18.
Rosen, G. (2000). Multi-factorial assessment of exogenous enzymes in broiler pronutrition: Target and problems. Proceedings of the 3rd European Symposium on Feed Enzymes, Noordwijkerhout, Netherlands.
Rosen, G., Vol. II, p141. (1996). Proceedings of the World's Poultry Science Society, New Delhi.
Rosen, G. D. (1995). "Antibacterials in poultry and pig nutrition. In Biotechnology in the Animal Feeds and Animal Feeding Ed by R.J. Wallace and A. Chesson. VCH Verlagsgesellschaft mbH D-69461 Weinheim." 8: 143-172.
Rosen, G. D. (2001). "Multi-factorial efficacy evaluation of alternatives to antimicrobials in pronutrition." British Poultry Science 42(S1): S104-S105.
Salanitro, J. P., I. G. Blake, et al. (1978). "Bacteria isolated from the duodenum, ileum, and cecum of young chicks." Appl Environ Microbiol 35(4): 782-90.
Salanitro, J. P., I. G. Blake, et al. (1974). "Studies on the cecal microflora of commercial broiler chickens." Appl Microbiol 28(3): 439-47.
Schildknect, E., L. Rakebrand, et al. (2003). Changes In Anticoccidial Sensitivity Profiles Of Coccidia From Broiler Chickens Raised On Built-Up Litter For Eight Production Cycles Following A Coccidiosis Challenge. International Poultry Scientific Forum Abstracts.
Schildknect, E., L. Rakebrand, et al. (2003). Changes In Live Performance Of Broiler Chickens Raised On Built-Up Litter For Eight Production Cycles Following A Coccidiosis. International Poultry Scientific Forum Abstracts, Atlanta, Georgia.
Schrag, S. J. and V. Perrot (1996). "Reducing antibiotic resistance." Nature 381(6578): 120-1.
Schrag, S. J., V. Perrot, et al. (1997). "Adaptation to the fitness costs of antibiotic resistance in Escherichia coli." Proc R Soc Lond B Biol Sci 264(1386): 1287-91.
Schwalbe, R. S., C. W. Hoge, et al. (1990). "In vivo selection for transmissible drug resistance in Salmonella typhi during antimicrobial therapy." Antimicrob Agents Chemother 34(1): 161-3.
Schwalbe, R. S., W. J. Ritz, et al. (1990). "Selection for vancomycin resistance in clinical isolates of Staphylococcus haemolyticus." J Infect Dis 161(1): 45-51.
Sell, J., J. Eastwood, et al. (1983). "Influence of supplemental fat on metabolizable energy and ingesta transit time in laying hens." Nutrition Rep. Intern. 28: 487-495.
Shane, S. M., R. G. Young, et al. (1969). "Renal and parathyroid changes produced by high calcium intake in growing pullets." Avian Diseases 13: 558-567.
Sharon, N. and H. Lis (1993). "Carbohydrates in cell recognition." Sci Am 268(1): 82-9.
Sibbald, I. R. (1979). "Passage of feed through the adult rooster." Poult Sci 58(2): 446-59.
Sklan, D., D. Dubrov, et al. (1975). "51Cr-EDTA, 91Y and 141Ce as nonabsorbed reference substances in the gastrointestinal tract of the chicken." J Nutr 105(12): 1549-52.
Sklan, D. and S. Hurwitz (1980). "Protein digestion and absorption in young chicks and turkeys." J Nutr 110(1): 139-44.
Spring, P. (1996). Effects of mannanoligosaccharide on different caecal parameters and on the attachment of enteric pathogens in poultry. Zurich, Swiss Fed. Inst. Technology.
Stappenbeck, T. S., L. V. Hooper, et al. (2002). "Developmental regulation of intestinal angiogenesis by indigenous microbes via Paneth cells." Proc Natl Acad Sci U S A 99(24): 15451- 5.
Stavric, S., T. Gleeson, et al. (1987). "Effect of environmental temperature on the susceptibility of young chickens to Salmonella typhimurium." Australian Veterinary Journal 55: 413.
Stewart, F. M., R. Antia, et al. (1998). "The population genetics of antibiotic resistance. II: Analytic theory for sustained populations of bacteria in a community of hosts." Theor Popul Biol 53(2): 152-65.
Sudo, S. and G. Duke (1980). "Kinetics of absorption of volatile fatty acids from the ceca of domestic turkeys. Comp. Biochem. Physiol." 67: 231-237.
Sullivan, A., C. Edlund, et al. (2001). "Effect of antimicrobial agents on the ecological balance of human microflora." Lancet Infect Dis 1(2): 101-14.
Swamy, H. V., T. K. Smith, et al. (2002). "Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on production and metabolism in broilers." Poult Sci 81(7): 966-75.
Swamy, H. V., T. K. Smith, et al. (2002). "Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on swine performance, brain regional neurochemistry, and serum chemistry and the efficacy of a polymeric glucomannan mycotoxin adsorbent." J Anim Sci 80(12): 3257-67.
Tahara, T., S. Yoshioka, et al. (1997). "Isolation and partial characterization of bacteriocins produced by Lactobacillus gasseri JCM 2124." FEMS Microbiol Lett 148(1): 97-100.
Takei, Y., Y. Okawara, et al. (1988). "Water intake induced by water deprivation in the quail, Coturnix coturnix japonica." J Comp Physiol [B] 158(5): 519-25.
Turkov, M. I., V. V. Belushkin, et al. (1990). "[The effect of small concentrations of antibiotics blocking the synthesis of bacterial cell wall on the permeability of Escherichia coli cell wall for plasmid DNA]." Genetika 26(3): 443-7.
Uden, P., P. E. Colucci, et al. (1980). "Investigation of chromium, cerium and cobalt as markers in digesta. Rate of passage studies." J Sci Food Agric 31(7): 625-32.
Ueda, Y., S. Hachimura, et al. (2001). "Apoptosis of antigen-specific T cells induced by oral administration of antigen: comparison of intestinal and non-intestinal immune organs." Biosci Biotechnol Biochem 65(5): 1170-4.
Uni, Z., A. Geyra, et al. (2000). "Small intestinal development in the young chick: crypt formation and enterocyte proliferation and migration." Br Poult Sci 41(5): 544-51.
Uni, Z., Y. Noy, et al. (1999). "Posthatch development of small intestinal function in the poult." Poult Sci 78(2): 215-22.
Vaughan, E. E., F. Schut, et al. (2000). "A molecular view of the intestinal ecosystem." Curr Issues Intest Microbiol 1(1): 1-12.
Vergara, P., C. Ferrando, et al. (1989). "Factors determining gastrointestinal transit time of several markers in the domestic fowl." Q J Exp Physiol 74(6): 867-74.
Washburn, K. W. (1991). "Efficiency of feed utilization and rate of feed passage through the digestive system." Poult Sci 70(3): 447-52.
Weurding, R. E., A. Veldman, et al. (2001). "Starch digestion rate in the small intestine of broiler chickens differs among feedstuffs." J Nutr 131(9): 2329-35.
Wideman, R. F. J. (1987). "Renal regulation of avian calcium and phosphorus metabolism." Jurnal of Nutrition 117: 808-814.
Wideman, R. F. J., J. A. Closser, et al. (1985). "Urolithiasis in pullets and laying hens: role of dietary calcium and phosphorus " Poult Sci 64(2300-2307).
Wise, R. (1999). "A review of the mechanisms of action and resistance of antimicrobial agents." Can Respir J 6 Suppl A: 20A-2A.
Woo, P. C., A. P. To, et al. (2003). "Facilitation of horizontal transfer of antimicrobial resistance by transformation of antibiotic-induced cell-wall-deficient bacteria." Med Hypotheses 61(4): 503-8.
Xu, J., M. K. Bjursell, et al. (2003). "A genomic view of the human-Bacteroides thetaiotaomicron symbiosis." Science 299(5615): 2074-6.
Xu, J. and J. I. Gordon (2003). "Inaugural Article: Honor thy symbionts." Proc Natl Acad Sci U S A 100(18): 10452-9.
Zambrano, M. M., D. A. Siegele, et al. (1993). "Microbial competition: Escherichia coli mutants that take over stationary phase cultures." Science 259(5102): 1757-60.
Zhu, W. M., W. Liu, et al. (2000). "Isolation and characterization of a new bacteriocin from Lactobacillus gasseri KT7." J Appl Microbiol 88(5): 877-86.
Zinser, E. R. and R. Kolter (2004). "Escherichia coli evolution during stationary phase." Res Microbiol 155(5): 328-36.
Related topics
Authors:

Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post
22 de septiembre de 2010
Dear Sir,
Its very elaborative article on WET Litter.Article shows a systematic approach on the topic presentation which will benefit to all the technical persons concerned.
I expect much more technical articles to be posted in future on the Engormix from your side, with the help of which more literacy will be created on the subject matter.
Thanks & regards,
Dr Jaydip Mulik (M.V.Sc.DCT)

Kemin Industries, Inc
22 de septiembre de 2010
Very good article and thanks Dr.Stephen R. Collet. One of the very valid point you brought in this article is the mycotoxin and its role in litter quality. There are n number of research papers stated that pathogenic microorganism with mycotoxins will have much adverse effect when compare to only mycotoxins or microorganism. At low level, mycotoxins will cause only the performance impact and along with other causative factors it will create much higher rious impact.
Once again thank you for the good article!
R Kannan
Product manager - Kemin







You may be interested in
.jpg&w=3840&q=75)