Identification and Comparative Expression Analysis of Interleukin 2/15 Receptor b Chain in Chickens Infected with E. tenella
Author details:
1 College of Veterinary Medicine and Research Institute of Life Science, Gyeongsang National University, Jinju, Korea, 2 Departments of Infectious Diseases and Avian Diseases, College of Veterinary Medicine and Korea Zoonosis Research Institute, Chonbuk National University, Jeonju, Korea, 3 National Institute of Animal Science, RDA, Cheonan, Chungnam, Korea, 4 Animal Parasitic Diseases Laboratory, Animal and Natural Resources Institute, Agricultural Research Service, United States Department of Agriculture, Beltsville, Maryland, United States of America.
Background: Interleukin (IL) 2 and IL15 receptor b chain (IL2/15Rb, CD122) play critical roles in signal transduction for the biological activities of IL2 and IL15. Increased knowledge of non-mammalian IL2/15Rb will enhance the understanding of IL2 and IL15 functions.
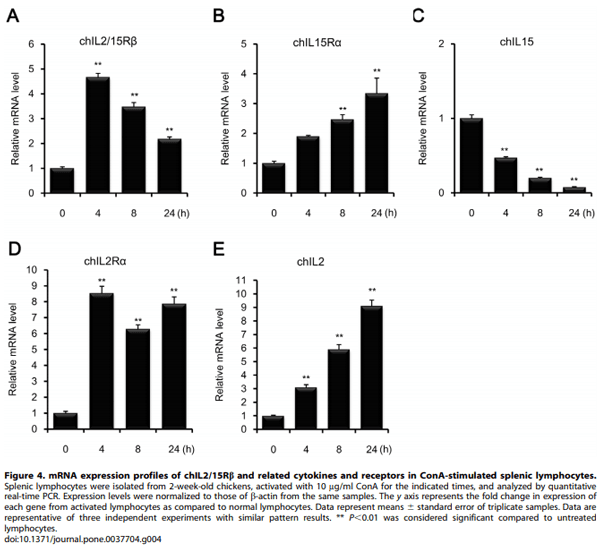
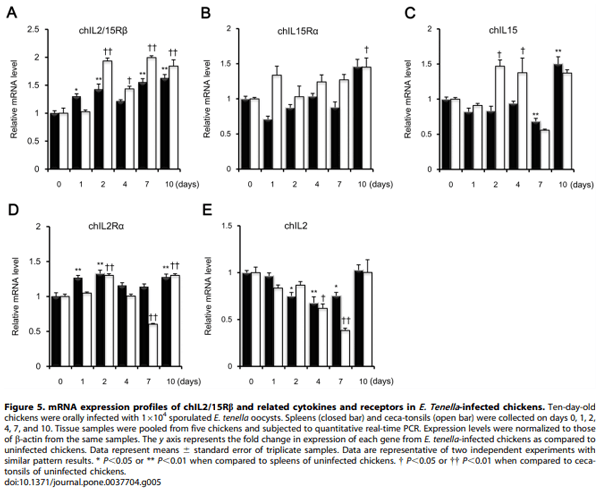
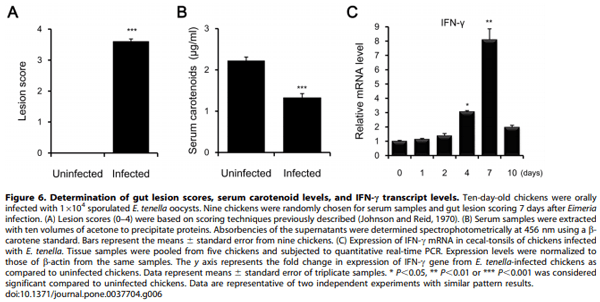
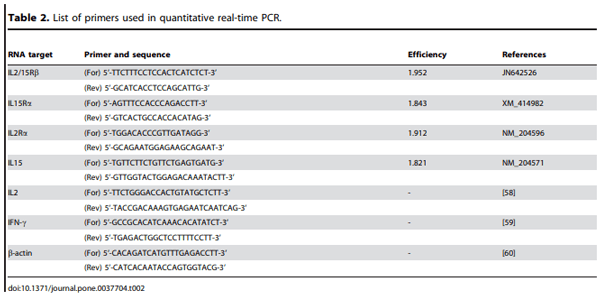
Methology/Principal Findings: Chicken IL2/15Rb (chIL2/15Rb) cDNA was cloned using 59/39-RACE. The predicted protein sequence contained 576 amino acids and typical features of the type-I cytokine receptor family. COS-7 cells transfected with chIL2/15Rb produced proteins of approximately 75 and 62.5 kDa under normal and tunicamycin-treated conditions, respectively. The genomic structure of chIL2/15Rb was similar to its mammalian counterparts. chIL2/15Rb transcripts were detected in the lymphoblast cell line CU205 and in normal lymphoid organs and at moderate levels in bursa samples. Expression profiles of chIL2/15Rb and its related cytokines and receptors were examined in ConA-stimulated splenic lymphocytes and in ceca-tonsils of Eimeria tenella-infected chickens using quantitative real-time PCR. Expression levels of chIL2/15Rb, chIL2Ra, and chIL15Ra were generally elevated in ceca-tonsils and ConA-activated splenic lymphocytes. However, chIL2 and chIL15 expression levels were differentially regulated between the samples. chIL2 expression was upregulated in ConA-activated splenic lymphocytes, but not in ceca-tonsils. In constrast, chIL15 expression was upregulated in ceca-tonsils, but not in ConA-activated splenic lymphocytes.
Conclusions/Significance: We identified an avian form of IL2/15Rb and compared its gene expression pattern with those of chIL2, chIL15, chIL2Ra, and chIL15Ra. Our observations suggest that chIL15 and its receptors, including chIL2/15Rb, play important roles in mucosal immunity to intestinal intracellular parasites such as Eimeria.
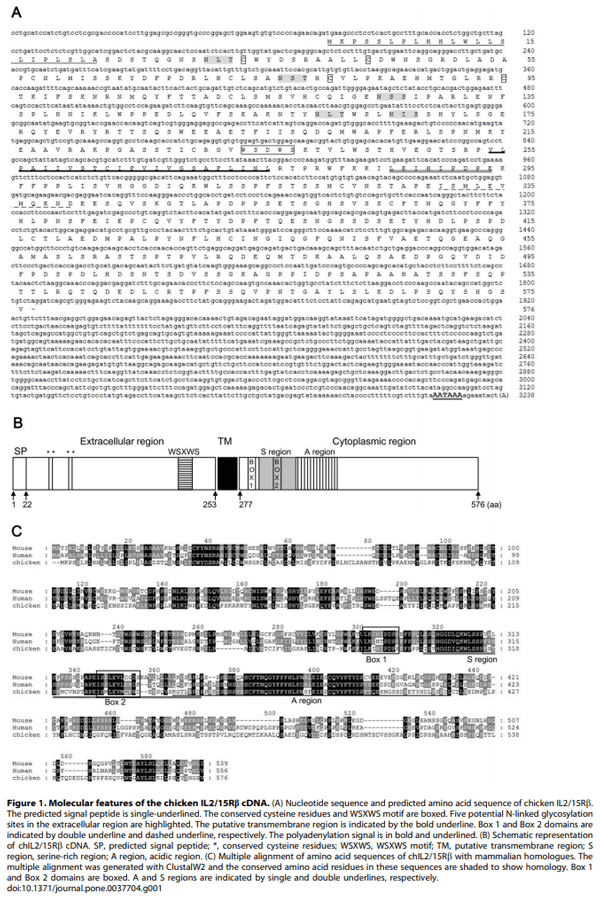
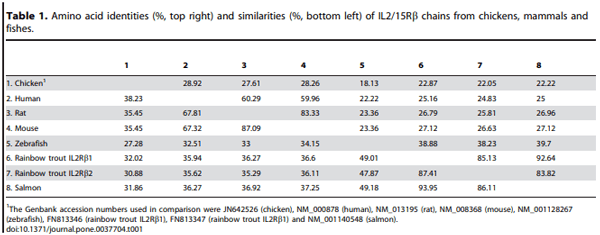
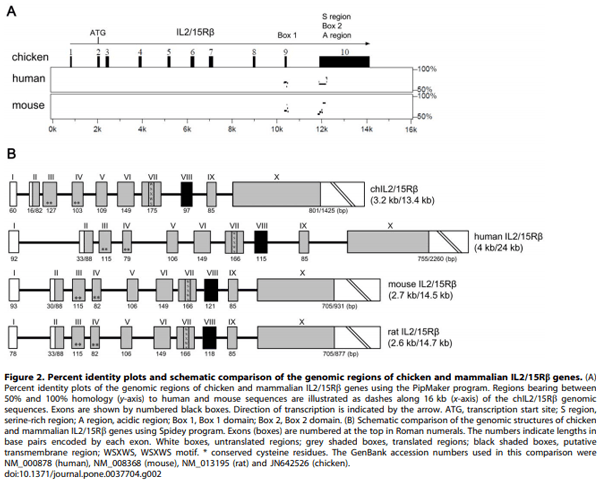
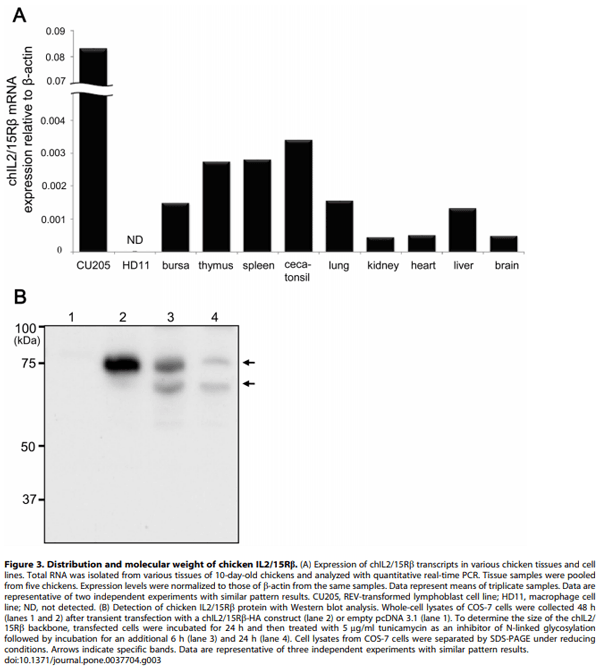
1. Ma A, Koka R, Burkett P (2006) Diverse functions of IL-2, IL-15, and IL-7 in lymphoid homeostasis. Annu Rev Immunol 24: 657–679.
2. Bodnar A, Nizsaloczki E, Mocsar G, Szaloki N, Waldmann TA, et al. (2008) A biophysical approach to IL-2 and IL-15 receptor function: localization, conformation and interactions. Immunol Lett 116: 117–125.
3. Gaffen SL (2001) Signaling domains of the interleukin 2 receptor. Cytokine 14, 63–77.
4. Burchill MA, Yang J, Vang KB, Farrar MA (2007) Interleukin-2 receptor signaling in regulatory T cell development and homeostasis. Immunol Lett 114: 1–8.
5. Rochman Y, Spolski R, Leonard WJ (2009) New insights into the regulation of T cells by cc family cytokines. Nat Rev Immunol 9: 480–490.
6. Tanaka T, Tsudo M, Karasuyama H, Kitamura F, Kono T, et al. (1991) A novel monoclonal antibody against murine IL-2 receptor b-chain. Characterization of receptor expression in normal lymphoid cells and EL-4 cells. J Immunol 147: 2222–2228.
7. David D, Bani L, Moreau JL, Demaison C, Sun K, et al. (1998) Further analysis of interleukin-2 receptor subunit expression on the different human peripheral blood mononuclear cell subsets. Blood 91: 165–172.
8. Sharon M, Siegel JP, Tosato G, Yodoi J, Gerrard TL, et al. (1988) The human interleukin 2 receptor b chain (p70). Direct identification, partial purification, and patterns of expression on peripheral blood mononuclear cells. J Exp Med 167: 1265–1270.
9. Wang G, Lu C, Liu H, Jin W, Jiao X, et al. (2001) Immunohistochemical localization of interleukin-2 and its receptor subunits a, b and c in the main olfactory bulb of the rat. Brain Res 893: 244–252.
10. Nielsen OH, Ciardelli T, Wu Z, Langholz E, Kirman I (1995) Circulating soluble interleukin-2 receptor a and b chain in inflammatory bowel disease. Am J Gastroenterol 90: 1301–1306.
11. Honda M, Kitamura K, Takeshita T, Sugamura K, Tokunaga T (1990) Identification of a soluble IL-2 receptor b -chain from human lymphoid cell line cells. J Immunol 145: 4131–4135.
12. Montes de Oca P, Malarde V, Proust R, Dautry-Varsat A, Gesbert F (2010) Ectodomain shedding of interleukin-2 receptor b and generation of an intracellular functional fragment. J Biol Chem 285: 22050–22058.
13. Suzuki H, Duncan GS, Takimoto H, Mak TW (1997) Abnormal development of intestinal intraepithelial lymphocytes and peripheral natural killer cells in mice lacking the IL-2 receptor b chain. J Exp Med 185: 499–505.
14. Gilmour KC, Fujii H, Cranston T, Davies EG, Kinnon C, et al. (2001) Defective expression of the interleukin-2/interleukin-15 receptor b subunit leads to a natural killer cell-deficient form of severe combined immunodeficiency. Blood 98: 877–879.
15. Suzuki H, Kundig TM, Furlonger C, Wakeham A, Timms E, et al. (1995) Deregulated T cell activation and autoimmunity in mice lacking interleukin-2 receptor b. Science 268: 1472–1476.
16. Malek TR, Yu A, Vincek V, Scibelli P, Kong L (2002) CD4 regulatory T cells prevent lethal autoimmunity in IL-2Rb-deficient mice. Implications for the nonredundant function of IL-2. Immunity 17: 167–178.
17. Kurreeman FA, Daha NA, Chang M, Catanese JJ, Begovich AB, et al. (2009) Association of IL2RA and IL2RB with rheumatoid arthritis: a replication study in a Dutch population. Ann Rheum Dis 68: 1789–1790.
18. Tanaka T, Kitamura F, Nagasaka Y, Kuida K, Suwa H, et al. (1993) Selective long-term elimination of natural killer cells in vivo by an anti-interleukin 2 receptor b chain monoclonal antibody in mice. J Exp Med 178: 1103–1107.
19. Yokoyama S, Watanabe N, Sato N, Perera PY, Filkoski L, et al. (2009) Antibody-mediated blockade of IL-15 reverses the autoimmune intestinal damage in transgenic mice that overexpress IL-15 in enterocytes. Proc Natl Acad Sci U S A 106: 15849–15854.
20. Sundick RS, Gill-Dixon C (1997) A cloned chicken lymphokine homologous to both mammalian IL-2 and IL-15. J Immunol 159: 720–725.
21. Lillehoj HS, Min W, Choi KD, Babu US, Burnside J, et al. (2001) Molecular, cellular, and functional characterization of chicken cytokines homologous to mammalian IL-15 and IL-2. Vet Immunol Immunopathol 82: 229–244.
22. Teng QY, Zhou JY, Wu JJ, Guo JQ, Shen HG (2006) Characterization of chicken interleukin 2 receptor a chain, a homolog to mammalian CD25. FEBS Lett 580: 4274–4281.
23. Li G, Lillehoj HS, Min W (2001) Production and characterization of monoclonal antibodies reactive with the chicken interleukin-15 receptor alpha chain. Vet Immunol Immunopathol 82: 215–227.
24. Min W, Lillehoj HS, Fetterer RH (2002) Identification of an alternatively spliced isoform of the common cytokine receptor c chain in chickens. Biochem Biophys Res Commun 299: 321–327.
25. Jeong J, Lee C, Yoo J, Koh PO, Kim YH, et al. (2011) Molecular identification of duck and quail common cytokine receptor c chain genes. Vet Immunol Immunopathol 140: 159–165.
26. Hatakeyama M, Tsudo M, Minamoto S, Kono T, Doi T, et al. (1989) Interleukin-2 receptor beta chain gene: generation of three receptor forms by cloned human a and b chain cDNA’s. Science 244: 551–556.
27. Kono T, Doi T, Yamada G, Hatakeyama M, Minamoto S, et al. (1990) Murine interleukin 2 receptor b chain: dysregulated gene expression in lymphoma line EL-4 caused by a promoter insertion. Proc Natl Acad Sci U S A 87: 1806–1810.
28. Page TH, Dallman MJ (1991) Molecular cloning of cDNAs for the rat interleukin 2 receptor a and b chain genes: differentially regulated gene activity in response to mitogenic stimulation. Eur J Immunol 21: 2133–2138.
29. Wang T, Huang W, Costa MM, Secombes CJ (2011) The gamma-chain cytokine/receptor system in fish: more ligands and receptors. Fish Shellfish Immunol 31: 673–687.
30. Nelson BH, Willerford DM (1998) Biology of the interleukin-2 receptor. Adv Immunol 70: 1–81.
31. Miyazaki T, Kawahara A, Fujii H, Nakagawa Y, Minami Y, et al. (1994) Functional activation of Jak1 and Jak3 by selective association with IL-2 receptor subunits. Science 266: 1045–1047.
32. Usacheva A, Sandoval R, Domanski P, Kotenko SV, Nelms K, et al. (2002) Contribution of the Box 1 and Box 2 motifs of cytokine receptors to Jak1 association and activation. J Biol Chem 277: 48220–48226.
33. Codias EK, Olosz F, Malek TR (2000) Genomic organization and 5’ regulatory region of the mouse IL-2 receptor b -chain gene (IL-2Rb). Immunogenetics 51: 508–512.
34. Shibuya H, Yoneyama M, Nakamura Y, Harada H, Hatakeyama M, et al. (1990) The human interleukin-2 receptor b -chain gene: genomic organization, promoter analysis and chromosomal assignment. Nucleic Acids Res 18: 3697–3703.
35. Hatakeyama M, Kono T, Kobayashi N, Kawahara A, Levin SD, et al. (1991) Interaction of the IL-2 receptor with the src-family kinase p56lck: identification of novel intermolecular association. Science 252: 1523–1528.
36. Kono T, Minami Y, Taniguchi T (1993) The interleukin-2 receptor complex and signal transduction: role of the beta-chain. Semin Immunol 5: 299–307.
37. Min W, Lillehoj HS (2002) Isolation and characterization of chicken interleukin17 cDNA. J Interferon Cytokine Res 22: 1123–1128.
38. Gobel TW, Kaspers B, Stangassinger M (2001) NK and T cells constitute two major, functionally distinct intestinal epithelial lymphocyte subsets in the chicken. Int Immunol 13: 757–762.
39. Cox GW, Mathieson BJ, Giardina SL, Varesio L (1990) Characterization of IL2 receptor expression and function on murine macrophages. J Immunol 145: 1719–1726.
40. Matsui K (1999) Role of interleukin-2 receptor expression on macrophages from Salmonella-infected mice. FEMS Immunol Med Microbiol 24: 97–103.
41. Tsudo M, Kozak RW, Goldman CK, Waldmann TA (1986) Demonstration of a non-Tac peptide that binds interleukin 2: a potential participant in a multichain interleukin 2 receptor complex. Proc Natl Acad Sci U S A 83: 9694–9698.
42. Asao H, Takeshita T, Nakamura M, Nagata K, Sugamura K (1990) The evidence for the IL-2 receptor b chain processing from p70 to p75. Int Immunol 2: 469–472.
43. Hilton LS, Bean AG, Kimpton WG, Lowenthal JW (2002) Interleukin-2 directly induces activation and proliferation of chicken T cells in vivo. J Interferon Cytokine Res 22: 755–763.
44. Vervelde L, Vermeulen AN, Jeurissen SH (1996) In situ characterization of leucocyte subpopulations after infection with Eimeria tenella in chickens. Parasite Immunol 18: 247–256.
45. Yun CH, Lillehoj HS, Choi KD (2000) Eimeria tenella infection induces local gamma interferon production and intestinal lymphocyte subpopulation changes. Infect Immun 68: 1282–1288.
46. Reinecker HC, MacDermott RP, Mirau S, Dignass A, Podolsky DK (1996) Intestinal epithelial cells both express and respond to interleukin 15. Gastroenterology 111: 1706–1713.
47. Hirose K, Suzuki H, Nishimura H, Mitani A, Washizu J, et al. (1998) Interleukin-15 may be responsible for early activation of intestinal intraepithelial lymphocytes after oral infection with Listeria monocytogenes in rats. Infect Immun 66: 5677–5683.
48. Inagaki-Ohara K, Nishimura H, Mitani A, Yoshikai Y (1997) Interleukin-15 preferentially promotes the growth of intestinal intraepithelial lymphocytes bearing cd T cell receptor in mice. Eur J Immunol 27: 2885–2891.
49. Ohta N, Hiroi T, Kweon MN, Kinoshita N, Jang MH, et al. (2002) IL-15- dependent activation-induced cell death-resistant Th1 type CD8 ab + NK1.1+ T cells for the development of small intestinal inflammation. J Immunol 169: 460–468.
50. Yun CH, Lillehoj HS, Lillehoj EP (2000) Intestinal immune responses to coccidiosis. Dev Comp Immunol 24: 303–324.
51. Min W, Lillehoj HS, Burnside J, Weining KC, Staeheli P, et al. (2001) Adjuvant effects of IL-1b, IL-2, IL-8, IL-15, IFN-a, IFN-c TGF- b4 and lymphotactin on DNA vaccination against Eimeria acervulina. Vaccine 20: 267–274.
52. Ma D, Ma C, Pan L, Li G, Yang J, et al. (2011) Vaccination of chickens with DNA vaccine encoding Eimeria acervulina 3–1E and chicken IL-15 offers protection against homologous challenge. Exp Parasitol 127: 208–214.
53. Yoo J, Kang SH, Jeong J, Kim WH, Kim S, et al. (2011) Effects of simple and disposable chicken cages for experimental eimeria infections. Korean J Parasitol 49: 299–302.
54. Schat KA, Pratt WD, Morgan R, Weinstock D, Calnek BW (1992) Stable transfection of reticuloendotheliosis virus-transformed lymphoblastoid cell lines. Avian Diseases 36: 432–439.
55. Beug H, von Kirchbach A, Doderlein G, Conscience JF, Graf T (1979) Chicken hematopoietic cells transformed by seven strains of defective avian leukemia viruses display three distinct phenotypes of differentiation. Cell 18: 375–390.
56. Johnson J, Reid WM (1970) Anticoccidial drugs: lesion scoring techniques in battery and floor-pen experiments with chickens. Exp Parasitol 28: 30–36.
57. Matthews JO, Ward TL, Southern LL (1997) Interactive effects of betaine and monensin in uninfected and Eimeria acervulina-infected chicks. Poult Sci 76: 1014–1019.
58. Liu H, Zhang M, Han H, Yuan J, Li Z (2010) Comparison of the expression of cytokine genes in the bursal tissues of the chickens following challenge with infectious bursal disease viruses of varying virulence. Virol J 7: 364.
59. Berndt A, Wilhelm A, Jugert C, Pieper J, Sachse K, et al. (2007) Chicken cecum immune response to Salmonella enterica serovars of different levels of invasiveness. Infect Immun 75: 5993–6007.
60. De Boever S, Vangestel C, De Backer P, Croubels S, Sys SU (2008) Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet Immunol Immunopathol 122: 312–317.
61. Schwartz S, Zhang Z, Frazer KA, Smit A, Riemer C, et al. (2000) PipMaker-A Web Server for Aligning Two Genomic DNA Sequences. Genome Res 10: 577–586.









.jpg&w=3840&q=75)