Introduction
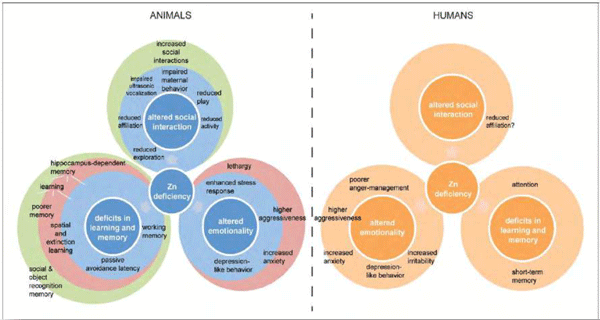
Trace minerals are essential elements in the maintenance of life for man and animal. However, what is not well understood or appreciated is how absorption and retention of these minerals can affect animal and human wellbeing, psychological status, health, and performance. While required in small amounts, just milligrams per day, trace minerals play critical roles as catalytic, regulatory, and structural components in multiple enzyme and protein systems. Zinc, for example, is a key component in over 300 enzyme systems and a structural part of over 3000 proteins. Yet as a “type-II nutrient”, zinc must be consumed on a daily basis to maintain normal bodily function. While trace minerals are key to cellular functions, their role prior to absorption, in maintenance and growth of the gut microbiome, is also important. Mineral source, amount and presence of dietary components, such as fiber, phytase, or even folic acid may lead to imbalances and/or insufficiencies, resulting in microbiome alteration with concomitant effects on health and neural function (Sauer and Grabrucker, 2019). Interestingly, as much as 20% of ingested zinc may be required by intestinal bacteria for growth and function (Smith et al., 1972). An insufficient intake of zinc has been linked to depression, mental lethargy, and cognitive impairment in human and animal models (Figure 1).
Mineral Absorption
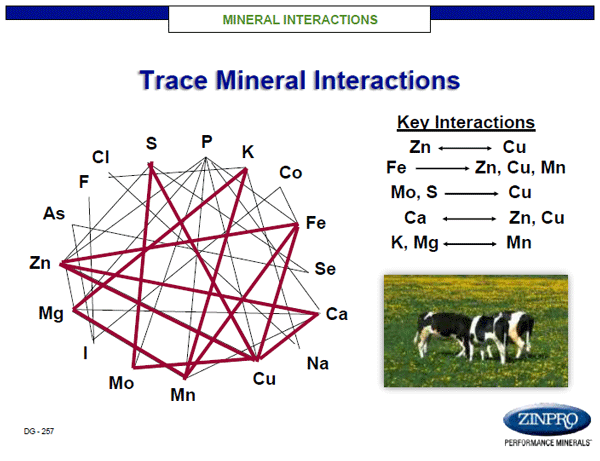
It is well established that both macro and trace minerals are capable of antagonizing absorption and retention of other minerals (Figure 2; NRC, 2001). The interrelationship between minerals and their requirements emphasize the importance of achieving a proper balance between amount and bio-efficacy. For example, when man or animal with normal zinc status consume excess zinc, the storage protein metallothionine is upregulated in intestinal enterocytes. This is a normal response, as metallothionine will hold absorbed zinc until it is either further moved into circulation or lost back into the gut for excretion via cellular turnover. The challenge is that metallothionine has a greater affinity for copper than zinc, thus copper movement into circulation may be antagonized by high zinc intake. The balance between zinc and copper is important in establishment and maintenance of gastro-intestinal (GI) microbiota and gut epithelial integrity, along with the potential for production of inflammatory cytokines (Sauer and Grabrucker, 2019). In a review, Vela et al. (2015) reported low levels of zinc and elevated copper have been linked to alterations in GI microbiota, increased intestinal epithelium permeability, and immune system abnormalities all increasing the severity of symptoms associated with autism spectrum disorder (ASD). It is well established that zinc plays a key role in these systems and mothers consuming an insufficient supply of zinc are at greater risk for GI tract dysbiosis, with an increased risk of skin challenges, immune depression, psychological depression, and higher risk of birth defects.
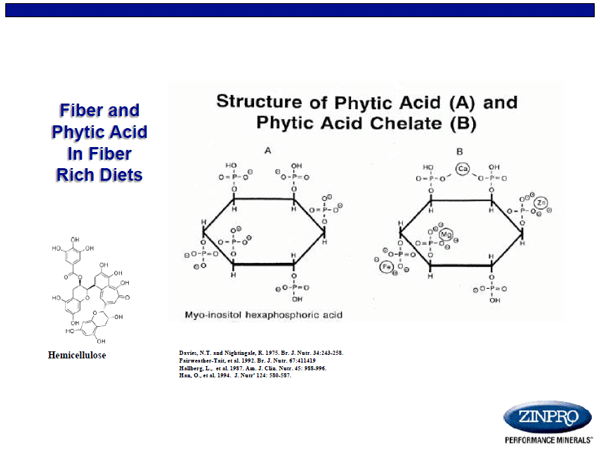
Fiber, phytase (Figure 3), and the presence of divalent cations such as calcium, iron, and copper are well established as potential antagonists to mineral absorption from the GI tract, specifically zinc, having been shown to reduce mineral availability in many species of animals and man (NRC, 2005). In humans, it is common for women to consume calcium supplements to mitigate the effects of osteoporosis or drink water naturally high in calcium. Similarly, women often take iron supplements to prevent anemia and to maintain sufficient red blood cell numbers during pregnancy to ensure proper fetal development. However, supplements meant to aid in offspring health and wellbeing may not always be as they seem. Supplementation of pregnant women with iron and folate is a common practice for prevention of gestational anemia and fetal neural tube defects. Work by Ghishan et al. (1986) demonstrated that zinc and folic acid have a mutually inhibitory effect at the intestinal level. This means that women and/or animals consuming folic acid may be at greater risk for zinc insufficiency, thus suffering subsequent effects of altered mineral status, with potential unfortunate effects on their offspring.
Early works at Zinpro Corporation found that sheep consuming a daily allocation of folic acid and zinc demonstrated a marked decline in circulating zinc status. These findings led us to further investigate the effects of folate supplementation on zinc retention. Our work, and that of Milne et al. (1984) demonstrated that folic acid supplementation resulted in increased fecal zinc excretion and a significant decline in urinary zinc, indicating a reduction in zinc retention and utilization. These findings are problematic, as Hagmeyer et al. (2015) reported that insufficient zinc intake/status is not only associated with skin lesions, growth retardation, poor appetite, delayed wound healing, cell-mediated immune dysfunction, and abnormal neurosensory changes, additionally, behavioral alterations have been consistently reported. Interestingly, recent findings in humans have documented an association between folate consumption, zinc insufficiency and depression, emotional instability, increased anxiety and aggression, irritability, and defects in social behavior. It is important to note that low serum zinc levels have been reported in Autism Spectrum Disorder (ASD), Attention Deficit Hyperactivity Disorder (ADHD), and mood disorders such as depression and schizophrenia (Hagmeyer et al., 2015).
Epigenetic Effects
Epigenetic effects are alterations inheritable phenotype without changes to the DNA sequence. For example, subclinical zinc deficiency in pregnant and lactating mothers is a possible cause for an immature and dysfunctional intestine in newborns (Vela et al., 2015). In a review, Vela et al. (2015) reported that maternal zinc status had teratogenic and pathological effects leading to developmental and functional effects in man and animal. Research indicated that low maternal zinc can result in impaired zinc absorption, leading to imbalance in zinc and copper, and behavior alterations in brain development in offspring.
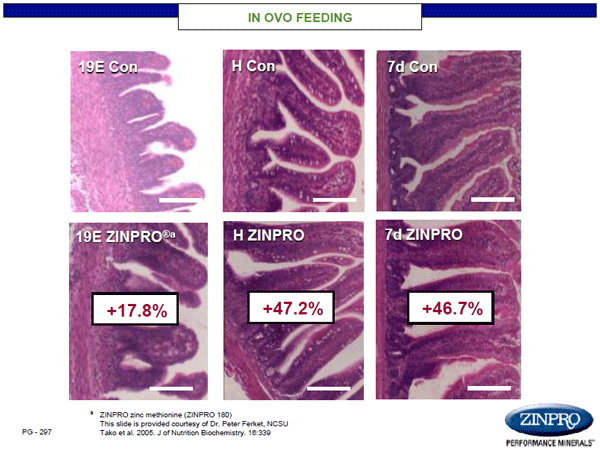
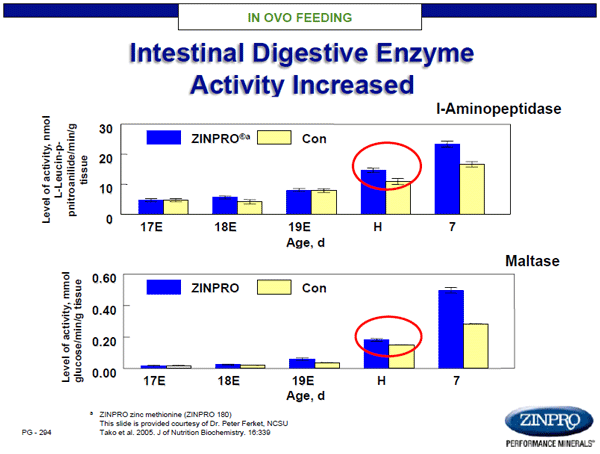
Unfortunately, mineral insufficiency creates a cascade of effects that are not easily delineated. For example, insufficient zinc often leads to an alteration in GI tract permeability, i.e. gut leakage, which results in inflammation and localization of inflammatory cytokines with immune cell migration. At the same time, GI microbiota may begin to shift at the phylum and genus level, altering digestive functionality, yield of end products, and disruption of communication between the gut and central nervous system (Strati et al., 2017). It is well established that individuals with ASD often suffer with GI co-morbidities, as a result of altered gut microbiota. Thus, maternal nutrition has direct effects on GI development, establishment of a normal microbiota, and GI communication with the brain in offspring. Notably, early works at Zinpro Corporation showed that chicken eggs injected with zinc methionine complex not only survived to hatch, but also showed marked improvements in GI tract development and function (Figures 4 & 5; Ferket, NCSU). While the chicks had morphological improvements in GI epithelial integrity and function, these changes resulted in > 7% enhancement in body weight growth through 14 days-of-age.
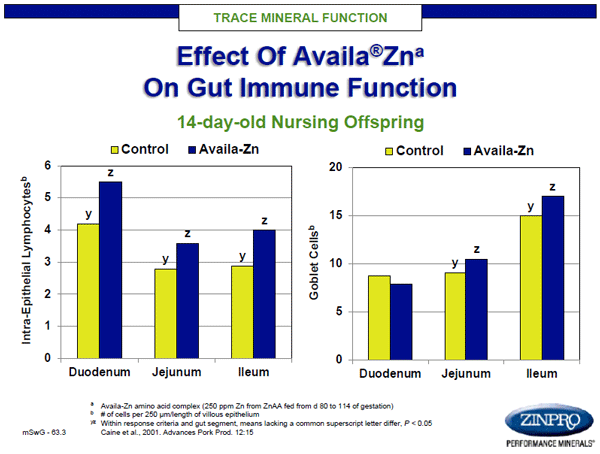
Similarly, researchers found that feeding sows zinc, as zinc amino acid complex (ZnAA), during the last trimester of pregnancy resulted in improved intestinal development of baby pigs (Figure 6; Caine et al., 2009). Offspring of sows fed ZnAA had increased villous height and villus height:crypt depth ratio in the jejunum, and greater goblet cell numbers in the ileum. These works demonstrate that maternal nutrition plays key roles in offspring morphological development and may also alter neural development and function.
Mineral Form and Bioavailability
Maternal nutrition is only part of the story, as mineral source is also a key component in meeting the nutritional requirements of man and animals. In general, inorganic minerals are less bioavailable and metabolically active than “organic” forms of trace minerals. A mineral is deemed organic when it is complexed or chelated to an amino acid, protein fragment, or other carbon-containing material. Unfortunately, much confusion and misinformation exists in regard to the true chemical structure and bio-efficacy of various organic mineral sources.
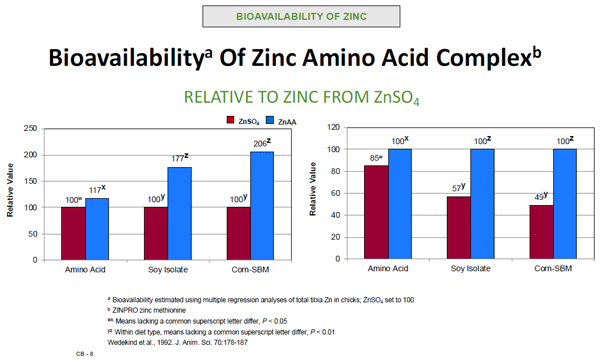
With regard to this particular discussion on mineral supplementation and its impact on GI tract integrity and prevention of GI microbiota dybiosis, Sauer and Grabrucker (2019) reported ZnAA was taken up by cells, not through classical zinc importer proteins, but rather by amino acid transporters. They noted ZnAA showed significant advantages over inorganic mineral salt supplements, as they did not compete with other metals for GI epithelium transporters and were less accessible for folic acid or phytic acid antagonism. This work is supported by earlier work demonstrating ZnAA was markedly less influenced by dietary antagonists, and therefore more bioavailable for absorption (Wedekind et al., 1994).
In unpublished work utilizing zinc knockout mice, workers at Vanderbilt University were able to create an ASD-type disorder in mouse pups by controlling maternal zinc supplementation. The pups were identified to have alterations in GI zinc and copper balance, impaired zinc uptake, and microbiome dysbiosis. Interestingly, expression of ASD in the pups was then reversed through supplementation with ZnAA. These findings were not totally unexpected, as work by Sauer and Grabrucker (2019) demonstrated that ZnAA was absorbed differently than inorganic zinc sources, and positively impacted GI permeability, GI microbiota, and reduced neuroinflammatory markers.
Conclusion
Human and animal research demonstrates that maternal mineral nutrition affects not only maternal health and wellbeing, but also that of her offspring. Maternal mineral insufficiencies have been shown to elicit epigenetic effects on offspring, impacting GI tract development and integrity, microbiota diversity, behavior impairment, and susceptibility to ASD, ADHD, and depression. Selection of bioavailable mineral sources, such as metal amino acid complexes, has shown to improve GI tract integrity, re-establish normal gut microbiota, and by enhancing mineral status improve neural function, behavior, and sense of wellbeing.
Figure 1. Comparative overview of behavioral alterations associated with zinc deficiency in animals and humans. Hagmeyer et al. 2015. Front. In Behav. Neurosci. 8:443
Figure 2. Macro and trace mineral interactions in man and animals.
Figure 3. Structure of phytic acid and its influence on mineral complexation and bioavailability in diets.
Figure 4. Impact of in ovo feeding with ZINPRO® zinc methionine on intestinal integrity of chicks.
Figure 5. Impact of in ovo feeding with ZINPRO® zinc methionine on intestinal digestive integrity of chicks.
Figure 6. GI tract integrity and functional effects in newborn baby pigs after late gestation maternal supplementation with zinc amino acid complex.
Figure 7. Impact of zinc source and antagonists on relative bioavailability.







.jpg&w=3840&q=75)