I. INTRODUCTION
Calcium (Ca) is an essential nutrient for poultry for biological processes such as bone development and egg shell formation (De Vries et al., 2010). Laying hens must be provided with Ca at approximately 4% of the total feed volume. Very little Ca is provided within the cereal grain component of the diet, therefore diets must be supplemented with sources of Ca. Dietary Ca is provided to layers as either a calcium carbonate grit or flour, such as limestone, with a small proportion being provided by the inorganic P source and also from meat and bone meal, if used in the diet. Although Ca is a necessary dietary component, the addition of limestone can result in a reduction of the digestibility of key nutrients in the diet, in particular phytate-P (Selle et al., 2009).
Commercial poultry diets are typically corn- and/or wheat-soy based and contain relatively high levels of phytate-P which has limited availability for poultry (Cowieson et al., 2004; Selle et al., 2009). Phytate carries a strong negative charge, and thus has a high affinity for divalent cations (Angel et al., 2002). Due to the high inclusion levels of Ca in the diet, Ca2+ is the dominant cation in the diet and chelates with phytate, forming Ca-phytate complexes (Selle et al., 2009). The formation of Ca-phytate complex reduces the bioavailability of both Ca and P, effecting skeletal health and egg quality. To improve the digestibility of phytate-P and other nutrients as well as to maintain skeletal integrity, physical and temporal separation of Ca from phytate may be necessary.
Laying hens are known to possess a specific appetite for Ca (Wood-Gush and Kare 1966; Hughes and Wood-Gush 1971; Joshua and Mueller 1979). This specific appetite is thought to originate from the ancestral Red Jungle Fowl, whereby the bird would have foraged consuming seeds, roots, insects and soil/silicates to satisfy nutritional requirements. This study aims to exploit the putative Ca specific appetite of layers to investigate the effect of diets with reduced dietary Ca concentration in conjunction with a separate source of Ca on bird performance and egg quality.
II. MATERIALS AND METHODS
All experimental procedures conducted had approval from The University of Sydney Animal Ethics Committee. A total of 80 Isa Brown laying hens (approximately 35-42 weeks of age) were randomly allocated to one of four dietary treatments, with 20 replicate birds per treatment. Dietary treatments were formulated to four different Ca concentrations (1, 2, 3 or 4%.), were wheat-soy based and formulated to an AME of 11.72 MJ/kg. Birds had access to a separate Ca source, approximately 95% CaCO3 with an average particle size of 1.4 mm (AB Grit, Omya Australia). Birds had ad libitum access to feed, water and the separate Ca source throughout the study. Birds were housed in individual cages (23cm W x 45cm D x 45cm H) in an environmentally controlled shed and were exposed to a lighting regime of 16h:8h (light:dark). Prior to the commencement of the study, birds were placed into cages two weeks beforehand and fed the 4% Ca. This was to ensure birds were habituated to their environment and reduce stress which could have affected the study’s results.
Individual bird body weight and plasma Ca an P concentrations was measured at the start and end of the trial. Approximately 3 ml of blood was collected from the brachial vein from each bird. Samples were refrigerated overnight before being spun to separate the plasma that were analysed for Ca an P. Feed intake, separate Ca source intake and egg production were recorded daily. Egg quality (egg weight, albumin height, dry eggshell weight, eggshell thickness and Haugh units) was measured three times per week. All data were analysed using REML including random effects in GenStat (14th Edition, VSN International).
III. RESULTS AND DISCUSSION
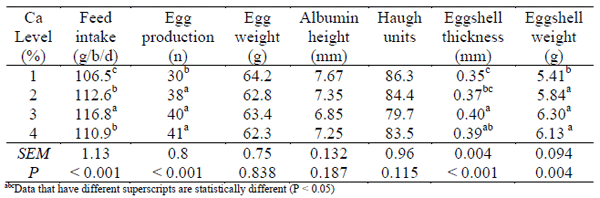
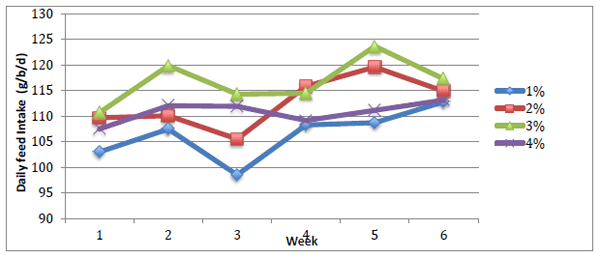
Feed and separate Ca intake were analysed using weekly values while egg quality parameters were analysed using treatment averages. Feed intake increased over time (Figure 1) and overall was significantly higher in birds fed 3% Ca and lowest in birds fed 1% Ca (Table 1). Birds fed 2 or 4% Ca diets were intermediary. Separate Ca intake (Figure 2) decreased with increasing dietary Ca concentration (P < 0.05). Body weight was not influenced by diet but birds were significantly lighter at the conclusion of the study (P = 0.003) with birds fed 1% Ca losing approximately 172g over the study. Birds fed 1% Ca had numerically lower plasma Ca and P concentrations at the conclusion of the study when compared to the start of the study while birds from the other groups had increased plasma Ca concentrations. Birds fed the 2% Ca diet had significantly lower plasma Ca concentration than birds fed 4% Ca.
Table 1 - Effect of dietary calcium concentrations on feed intake, egg quality
The influence of dietary Ca on egg quality results are presented in Table 1. Egg production increased with increasing dietary Ca and birds fed 1% Ca diets laid significantly less eggs than birds fed the other diets. Eggshells from birds fed 1% Ca diets were significantly thinner than those from birds fed diets containing 3 and 4% Ca. Dry egg shell weight was lowest in birds fed diets containing 1% Ca when compared to the other groups (P < 0.05). No significant difference between groups was observed for egg weight, albumin height or Haugh units.
Figure 1 - Average weekly feed intake of diets over the experimental period. Diet P < 0.001, Week P = 0.004, SEM 1.132.
Figure 2 - Average weekly separate calcium source intake over the experimental period. Diet P = 0.007, Week P = 0.142, SEM 3.371.
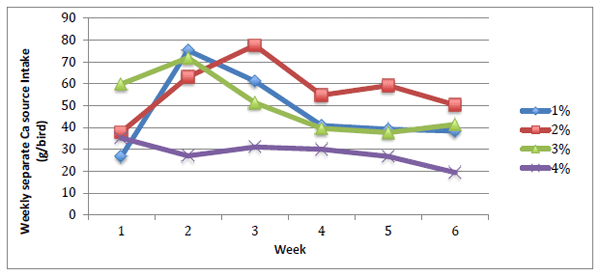
Poultry have been shown to possess a specific appetite for Ca (Wood-Gush and Kare 1966; Hughes and Wood-Gush 1971; Joshua and Mueller 1979) and the results of the present study are in general agreement with this. Birds decreased their consumption of the separate Ca source with increasing dietary Ca. This is consistent with the findings of Griminger and Lutz (1964) who showed birds consumed less supplemental Ca when fed high Ca diets, while birds had a high intake of supplemental Ca when fed low Ca diets. The results of the present study also show that some birds fed low Ca diets did not adapt to separate Ca choice feeding, particularly those fed 1% Ca diets and it is possible that 1% dietary Ca was too low in the context of this study. Similarly, Salim (1981) showed that some hens refused to eat a Ca supplement when dietary Ca was as low as 0.5%. The results did show that feeding diets with 2% Ca is feasible, however, birds may take time to find an equilibrium. It appears that birds fed diets with 1-3% Ca were able to recognise the Ca deficiency in week 1 and over compensated for this in weeks 2 and 3 before reaching an equilibrium point in week 4. If the 1% dietary Ca groups is excluded from the results from week three onwards, the consumption of the separate Ca source follows the assumption that birds on a low Ca diet will increase their separate Ca intake accordingly.
Birds fed 1% Ca diets produced significantly less eggs, with thinner and lighter eggshell weights and may be described as being Ca deficient. Ca deficiency decreases egg production and a reduces the thickness of the eggshell (Scott et al., 1971). Several birds in this study fed the 1% Ca diet ceased egg production and one bird in particular regularly ate its own egg shell, possibly in an attempt to obtain Ca.
It can be concluded that the Ca-specific appetite is not universally or uniformly expressed in laying hens. Though some birds were able to regulate to a common Ca intake and maintain production when fed reduced dietary Ca, others were unable to adapt to this new feeding regime. Further work is required to explore the optimum time, ontogenetically, to introduce a separate Ca source and to consider groups of birds where birds with reduced ability to self-regulate may learn from the early adopters. The effect of separating Ca from phytate on nutrient digestibility in layers is currently being assessed. In the future it may be possible to spatially separate Ca from the basal diet and exploit the Ca-specific appetite of layer hens to specifically improve the digestibility of phytate-P and amino acids. This would enhance the profitability and sustainability of the egg industry.
ACKNOWLEDGMENTS: This study was funded by the Australian Egg Corporation Limited and the senior author is in receipt of a scholarship from the Poultry CRC. Professor Peter Thomson (University of Sydney) is thanked for his assistance with statistical analysis
REFERENCES
Angel R, Tamim NM, Applegate TJ, Dhandu AS & Ellestad LE (2002) Journal of Applied Poultry Research 11: 471-480.
Cowieson AJ, Acamovic T & Bedford MR (2004) British Poultry Science 45: 101-108.
De Vries S, Kwakkel RP & Dijkstra J (2010) In: Phosphorus and Calcium utilisation and Requirements in Farm Animals (DMSS Vitti, E Kebreab) CABI International.
Griminger P & Lutz H (1964) Poultry Science 43: 710-716.
Hughes BO & Wood-Gush DGM (1971) Animal Behaviour 19: 490-499.
Joshua IG & Mueller WJ (1979) British Poultry Science 20: 481-490.
Salim AA (1981) PhD Dissertation, University of Nebraska, Lincoln, NE
Scott ML, Hull SJ & Mullenhoff PA (1971) Poultry Science 50: 1055-1063.
Selle PH, Cowieson AJ & Ravindran V (2009) Livestock Science 124: 126-141.
Wood-Gush DGM & Kare MR (1966) British Poultry Science 7: 285-290.