Evaluation of Bone Marrow Adipose Tissue and Bone Mineralization on Broiler Chickens Affected by Wooden Breast Myopathy
Author details:
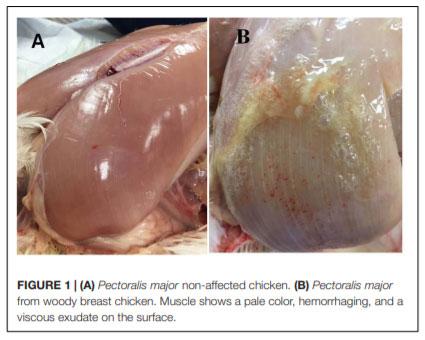
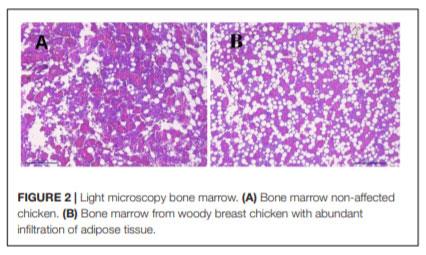
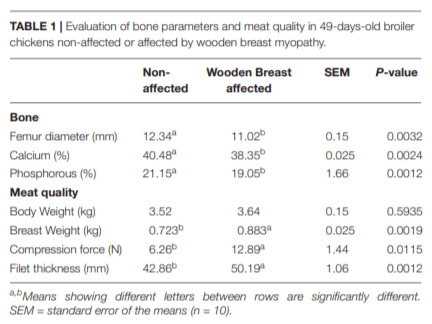
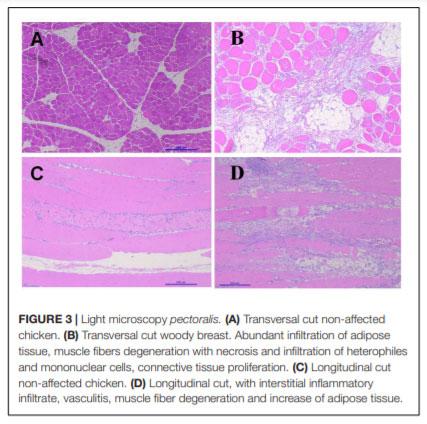
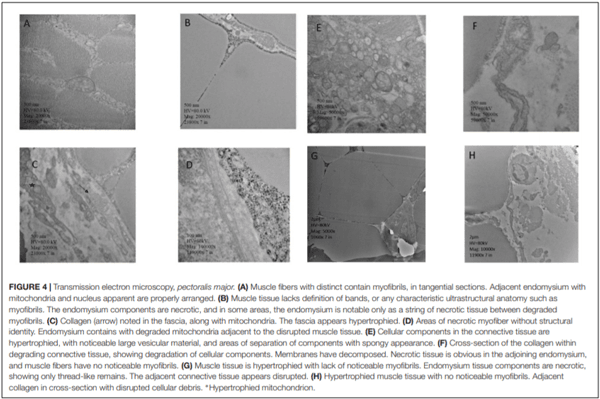
In humans, alterations in bone metabolism have been associated with myopathies. We postulate the hypothesis that perhaps similar pathologies can also be associated in modern chickens. Hence, this study aimed to assess the fat infiltration in bone marrow and its repercussion on broiler chicken affected by Wooden Breast (WB) myopathy. Ten Cobb 500 live birds with extreme rigidity of the Pectoralis major (PM) muscle were selected as WB affected chickens by physical examination of the muscle at 49 days of age, whereas ten chickens healthy with no physical signs of hardness in the breast muscle were considered to be unaffected. Macroscopic lesions in affected chickens included areas of firm and inflamed muscle with pale appearance, hemorrhaging, and viscous exudate on the surface. Bone marrow and sections of the PM muscle were collected and analyzed for light microscopy. Additionally, transmission electron microscopy was conducted in affected or unaffected muscle. Chickens affected with WB showed significant reductions (P < 0.05) in femur diameter, calcium, and phosphorous percentage but increased breast weight, compression force and filet thickness when compared with non-affected chickens. Interestingly, bone marrow from WB chicken had subjectively, more abundant infiltration of adipose tissue, when compared with non-affected chickens. Histology of the Pectoralis major of birds with WB showed abundant infiltration of adipose tissue, muscle fibers degeneration with necrosis and infiltration of heterophils and mononuclear cells, connective tissue proliferation, and vasculitis. Ultrastructural changes of WB muscle revealed lack definition of bands in muscle tissue, or any normal ultrastructural anatomy such as myofibrils. The endomysium components were necrotic, and in some areas, the endomysium was notable only as a string of necrotic tissue between degraded myofibrils. The fascia appeared hypertrophied, with large areas of necrosis and myofiber without structural identity with degraded mitochondria adjacent to the disrupted muscle tissue. As far as we know, this is the first study that describes a subjective increase in adipose tissue in the bone marrow of chickens affected with WB when compared with non-affected chickens, and reduced bone mineralization.
Keywords: bone marrow adipose tissue, broiler chickens, wooden breast, histology, electron microscopy.
Abdalla, B. A., Chen, J., Nie, Q., and Zhang, X. (2018). Genomic insights into the multiple factors controlling abdominal fat deposition in a chicken model. Front. Genet. 9:262. doi: 10.3389/fgene.2018.00262
Adams, G. R. (2002). Invited review: autocrine/paracrine IGF-I and skeletal muscle adaptation. J. Appl. Physiol. 93, 1159–1167. doi: 10.1152/japplphysiol.01264. 2001
Akhmedov, D., and Berdeaux, R. (2013). The e?ects of obesity on skeletal muscle regeneration. Front. Physiol. 4:371. doi: 10.3389/fphys.2013.00371
Alnahhas, N., Berri, C., Chabault, M., Chartrin, P., Boulay, M., Bourin, M. C., et al. (2016). Genetic parameters of white striping in relation to body weight, carcass composition, and meat quality traits in two broiler lines divergently selected for the ultimate pH of the pectoralis major muscle. BMC Genet. 17:61. doi: 10.1186/s12863-016-0369-2
Awano, H., Blaeser, A., Wu, B., Lu, P., Keramaris-Vrantsis, E., and Lu, Q. (2015). Dystroglycanopathy muscles lacking functional glycosylation of alpha-dystroglycan retain regeneration capacity. Neuromuscul. Disord. 25, 474–484. doi: 10.1016/j.nmd.2015.03.004
Baxter, M. F., Merino-Guzman, R., Latorre, J. D., Maha?ey, B. D., Yang, Y., Teague, K. D., et al. (2017). Optimizing fluorescein isothiocyanate dextran measurement as a biomarker in a 24-h feed restriction model to induce gut permeability in broiler chickens. Front. Vet. Sci. 4:56. doi: 10.3389/fvets.2017.00056
Bijnen, M., Josefs, T., Cuijpers, I., Maalsen, C. J., van de Gaar, J., Vroomen, M., et al. (2018). Adipose tissue macrophages induce hepatic neutrophil recruitment and macrophage accumulation in mice. Gut 67, 1317–1327. doi: 10.1136/gutjnl-2016-313654
Blem, C. R. (1976). Patterns of lipid storage and utilization in birds. Am. Zool. 16, 671–684. doi: 10.1093/icb/16.4.671
Bornelöv, S., Seroussi, E., Yosefi, S., Benjamini, S., Miyara, S., Ruzal, M., et al. (2018). Comparative omics and feeding manipulations in chicken indicate a shift of the endocrine role of visceral fat towards reproduction. BMC Genomics 19:295. doi: 10.1186/s12864-018-4675-0
Bourneuf, E., Hérault, F., Chicault, C., Carré, W., Assaf, S., Monnier, A., et al. (2006). Microarray analysis of di?erential gene expression in the liver of lean and fat chickens. Gene 372, 162–170. doi: 10.1016/j.gene.2005.12.028
Burt, D. (2007). Emergence of the chicken as a model organism: implications for agriculture and biology. Poult. Sci. 86, 1460–1471. doi: 10.1093/ps/86.7.1460 Carotenuto, F., Coletti, D., Di Nardo, P., and Teodori, L. (2016). a-linolenic acid reduces TNF-induced apoptosis in C2C12 myoblasts by regulating expression of apoptotic proteins. Eur. J. Transl. Myol. 26:6033. doi: 10.4081/ejtm.2016. 6033
Cawthorn, W. P., and Scheller, E. L. (2017). Bone marrow adipose tissue: formation, function, and impact on health and disease. Front. Endocrinol. 8:112. doi: 10.3389/fendo.2017.00112
Collins, K. H., Herzog, W., MacDonald, G. Z., Reimer, R. A., Rios, J. L., Smith, I. C., et al. (2018). Obesity, metabolic syndrome, and musculoskeletal disease: common inflammatory pathways suggest a central role for loss of muscle integrity. Front. Physiol. 9:112. doi: 10.3389/fphys.2018.00112
De Boer, A. A., Monk, J. M., Liddle, D. M., Hutchinson, A. L., Power, K. A., Ma, D. W., et al. (2016). Fish-oil-derived n-3 polyunsaturated fatty acids reduce NLRP3 inflammasome activity and obesity-related inflammatory cross-talk between adipocytes and CD11bC macrophages. J. Nutr. Biochem. 34, 61–72. doi: 10.1016/j.jnutbio.2016.04.004
Dodson, M. V., Hausman, G. J., Guan, L., Du, M., Rasmussen, T. P., Poulos, S. P., et al. (2010). Lipid metabolism, adipocyte depot physiology and utilization of meat animals as experimental models for metabolic research. Int. J. Biol. Sci. 6:691. doi: 10.7150/ijbs.6.691
Engin, A (2017). “Adipose tissue hypoxia in obesity and its impact on preadipocytes and macrophages: hypoxia hypothesis,” in Obesity and Lipotoxicity, eds A. B.
Engin and A. Engin (Cham: Springer), 305–326. doi: 10.1007/978-3-319-48382-5_13
Engin, A. B (2017). “Adipocyte-macrophage cross-talk in obesity,” in Obesity and Lipotoxicity, eds A. B. Engin and A. Engin (Cham: Springer), 327–343. doi: 10.1007/978-3-319-48382-5_14
Espinosa-Diez, C., Miguel, V., Mennerich, D., Kietzmann, T., Sánchez-Pérez, P., Cadenas, S., et al. (2015). Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 6, 183–197. doi: 10.1016/j.redox.2015.07.008
Fu, X., Zhu, M., Zhang, S., Foretz, M., Viollet, B., and Du, M. (2016). Obesity impairs skeletal muscle regeneration through inhibition of AMPK. Diabetes 65, 188–200. doi: 10.2337/db15-0647
Halevy, O., Geyra, A., Barak, M., Uni, Z., and Sklan, D. (2000). Early posthatch starvation decreases satellite cell proliferation and skeletal muscle growth in chicks. J. Nutr. 130, 858–864. doi: 10.1093/jn/130.4.858
Han, T., Tajar, A., and Lean, M. (2011). Obesity and weight management in the elderly. Br. Med. Bull. 97, 169–196. doi: 10.1093/bmb/ldr002
Hardouin, P., Rharass, T., and Lucas, S. (2016). Bone marrow adipose tissue: to be or not to be a typical adipose tissue? Front. Endocrinol. 7:85. doi: 10.3389/fendo. 2016.00085
Hausman, G., Dodson, M., Ajuwon, K., Azain, M., Barnes, K., Guan, L., et al. (2009). Board-invited review: the biology and regulation of preadipocytes and adipocytes in meat animals. J. Anim. Sci. 87, 1218–1246. doi: 10.2527/jas.2008-1427
Huang, X., and Ahn, D. U. (2018). The Incidence of muscle abnormalities in broiler breast meat-a review. Korean J. Food Sci. Anim. Resour. 38, 835–850. doi: 10.5851/kosfa.2018.e2
Ingalls, A. M., Dickie, M. M., and Shell, G. (1996). Obese, a new mutation in the house mouse. Obes. Res. 4:101. doi: 10.1002/j.1550-8528.1996.tb00519.x
International, A. O. A. C. (2000). “Animal feeds,” in O?cial Methods of Analysis of AOAC International, ed. W. Horwaitz (Gaithersburg, MD: AOAC International), 1–54.
Johnston, D. W. (1971). The absence of brown adipose tissue in birds. Comp. Biochem. Physiol. 40, l107–l108.
Karalaki, M., Fili, S., Philippou, A., and Koutsilieris, M. (2009). Muscle regeneration: cellular and molecular events. In Vivo 23, 779–796.
King, J. R. (1967). Adipose tissue composition in experimentally induced fat deposition in the White-crowned Sparrow. Comp. Biochem. Physiol. 21, 393– 403. doi: 10.1016/0010-406x(67)90801-8
Knowles, T. G., Kestin, S. C., Haslam, S. M., Brown, S. N., Green, L. E., Butterworth, A., et al. (2008). Leg disorders in broiler chickens: prevalence, risk factors and prevention. PLoS One 3:e1545. doi: 10.1371/journal.pone.0001545
Kozakowska, M., Pietraszek-Gremplewicz, K., Jozkowicz, A., and Dulak, J. (2015). The role of oxidative stress in skeletal muscle injury and regeneration: focus on antioxidant enzymes. J. Muscle Res. Cell. Motil. 36, 377–393. doi: 10.1007/ s10974-015-9438-9
Kuang, S., Kuroda, K., Le Grand, F., and Rudnicki, M. A. (2007). Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell 129, 999–1010. doi: 10.1016/j.cell.2007.03.044
Kuttappan, V. A., Bottje, W., Ramnathan, R., Hartson, S. D., Coon, C. N., Kong, B.-W., et al. (2017). Proteomic analysis reveals changes in carbohydrate and protein metabolism associated with broiler breast myopathy. Poult. Sci. 96, 2992–2999. doi: 10.3382/ps/pex069
Kuttappan, V. A., Hargis, B. M., and Owens, C. M. (2016). White striping and woody breast myopathies in the modern poultry industry: a review. Poult. Sci. 95, 2724–2733. doi: 10.3382/ps/pew216
Laumonier, T., and Menetrey, J. (2016). Muscle injuries and strategies for improving their repair. J. Exp. Orthop. 3:15. doi: 10.1186/s40634-016-0051-7 Leng, L., Zhang, H., Dong, J. Q., Wang, Z. P., Zhang, X. Y., Wang, S. Z., et al. (2016).
Selection against abdominal fat percentage may increase intramuscular fat content in broilers. J. Anim. Breed. Genet. 133, 422–428. doi: 10.1111/jbg.12204
Lepper, C., Partridge, T. A., and Fan, C.-M. (2011). An absolute requirement for Pax7-positive satellite cells in acute injury-induced skeletal muscle regeneration. Development 138, 3639–3646. doi: 10.1242/dev.067595
Livingston, M., Ferket, P., Brake, J., and Livingston, K. (2018). Dietary amino acids under hypoxic conditions exacerbates muscle myopathies including wooden breast and white stripping. Poult. Sci. 98, 1517–1527. doi: 10.3382/ps/pey463
Marchesi, J. A. P., Ibelli, A. M. G., Peixoto, J. O., Cantão, M. E., Pandolfi, J. R. C., Marciano, C. M. M., et al. (2018). Whole transcriptome analysis of the pectoralis major muscle reveals molecular mechanisms involved with white striping in broiler chickens. Poult. Sci. 98, 590–601. doi: 10.3382/ps/pey429
Matsuzawa, Y., Shimomurn, I., Nakumura, Y., Keno, Y., Kotani, K., Tokunaga, K., et al. (1995). Pathophysiology and pathogenesis of visceral fat obesity. Obes. Res. 3, 187s–194s. doi: 10.1002/j.1550-8528.1995.tb00462.x
Meloche, K. J., Dozier, W. A. III, Brandebourg, T. D., and Starkey, J. D. (2018). Skeletal muscle growth characteristics and myogenic stem cell activity in broiler chickens a?ected by wooden breast. Poult. Sci. 97, 4401–4414. doi: 10.3382/ps/ pey287
Meyer, G. A., and Ward, S. R. (2016). Developmental biology and regenerative medicine: addressing the vexing problem of persistent muscle atrophy in the chronically torn human rotator cu?. Phys. Ther. 96, 722–733. doi: 10.2522/ptj. 20150029
Miska, K. B., and Fetterer, R. H. (2019). Expression of amino acid and sugar transporters, aminopeptidase, and the di-and tri-peptide transporter PepT1; di?erences between modern fast growing broilers and broilers not selected for rapid growth. Poult. Sci. 98, 2272–2280. doi: 10.3382/ps/pey583
Mudalal, S., Lorenzi, M., Soglia, F., Cavani, C., and Petracci, M. (2015). Implications of white striping and wooden breast abnormalities on quality traits of raw and marinated chicken meat. Animal 9, 728–734. doi: 10.1017/ S175173111400295X
Mutryn, M. F., Brannick, E. M., Fu, W., Lee, W. R., and Abasht, B. (2015). Characterization of a novel chicken muscle disorder through di?erential gene expression and pathway analysis using RNA-sequencing. BMC Genomics 16:399. doi: 10.1186/s12864-015-1623-0
Ouali, A., Gagaoua, M., Boudida, Y., Becila, S., Boudjellal, A., Herrera-Mendez, C. H., et al. (2013). Biomarkers of meat tenderness: present knowledge and perspectives in regards to our current understanding of the mechanisms involved. Meat Sci. 95, 854–870. doi: 10.1016/j.meatsci.2013.05.010
Rambold, A. S., and Pearce, E. L. (2018). Mitochondrial dynamics at the interface of immune cell metabolism and function. Trends Immunol. 39, 6–18. doi: 10. 1016/j.it.2017.08.006
Resnyk, C. W., Carré, W., Wang, X., Porter, T. E., Simon, J., Le Bihan-Duval, E., et al. (2017). Transcriptional analysis of abdominal fat in chickens divergently selected on bodyweight at two ages reveals novel mechanisms controlling adiposity: validating visceral adipose tissue as a dynamic endocrine and metabolic organ. BMC Genomics 18:626. doi: 10.1186/s12864-017-4035-5
Riondino, S., Roselli, M., Palmirotta, R., Della-Morte, D., Ferroni, P., Guadagni, F., et al. (2014). Obesity and colorectal cancer: role of adipokines in tumor initiation and progression. World J. Gastroenterol. 20:5177. doi: 10.3748/wjg. v20.i18.5177
Saely, C. H., Geiger, K., and Drexel, H. (2012). Brown versus white adipose tissue: a mini-review. Gerontology 58, 15–23. doi: 10.1159/000321319
SAS Institute Inc. (2002). 9.4 User’s Guide. Cary, NC: SAS Documentation.
Schwarz, K., Siddiqi, N., Singh, S., Neil, C. J., Dawson, D., Frenneaux, M. P., et al.
(2014). The breathing heart - mitochondrial respiratory chain dysfunction in cardiac disease. Int. J. Cardiol. 171, 134–143. doi: 10.1016/j.ijcard.2013.12.014
Soglia, F., Gao, J., Mazzoni, M., Puolanne, E., Cavani, C., Petracci, M., et al. (2017). Superficial and deep changes of histology, texture and particle size distribution in broiler wooden breast muscle during refrigerated storage. Poult. Sci. 96, 3465–3472. doi: 10.3382/ps/pex115
Soglia, F., Mudalal, S., Babini, E., Di Nunzio, M., Mazzoni, M., Sirri, F., et al. (2015). Histology, composition, and quality traits of chicken Pectoralis major muscle a?ected by wooden breast abnormality. Poult. Sci. 95, 651–659. doi: 10.3382/ps/pev353
Soglia, F., Zeng, Z., Gao, J., Puolanne, E., Cavani, C., Petracci, M., et al. (2018). Evolution of proteolytic indicators during storage of broiler wooden breast meat. Poult. Sci. 97, 1448–1455. doi: 10.3382/ps/pex398
Stern, C. D. (2005). The chick: a great model system becomes even greater. Dev. Cell 8, 9–17. doi: 10.1016/s1534-5807(04)00425-3
Thomas, D., and Apovian, C. (2017). Macrophage functions in lean and obese adipose tissue. Metabolism 72, 120–143. doi: 10.1016/j.metabol.2017.04.005 Tumová,° E., and Teimouri, A. (2010). Fat deposition in the broiler chicken: a review. Sci. Agric. Biochem. 41, 121–128. doi: 10.3382/ps/pex184
Velleman, S. (2018). Association of posthatch muscle growth with the progression of breast muscle myopathies. J. Anim. Sci. 96:96. doi: 10.1093/jas/sky073.178 Velleman, S. G., and Clark, D. L. (2015). Histopathologic and myogenic gene expression changes associated with wooden breast in broiler breast muscles. Avian Dis. 59, 410–418. doi: 10.1637/11097-042015-reg.1
Velleman, S. G., Clark, D. L., and Tonniges, J. R. (2017). Fibrillar collagen organization associated with broiler wooden breast fibrotic myopathy. Avian Dis. 61, 481–490. doi: 10.1637/11738-080217-Reg.1
Velleman, S. G., McFarland, D. C., Li, Z., Ferrin, N. H., Whitmoyer, R., Dennis, J. E., et al. (1997). Alterations in sarcomere structure, collagen organization, mitochondrial activity, and protein metabolism in the avian low score normal muscle weakness. Dev. Growth Di?er. 39, 563–570. doi: 10.1046/j.1440-169x. 1997.t01-4-00003.x
Wang, G., Kim, W. K., Cline, M. A., and Gilbert, E. R. (2017). Factors a?ecting adipose tissue development in chickens: a review. Poult. Sci. 96, 3687–3699. doi: 10.3382/ps/pex184
Wang, H., Li, H., Wang, Q., Wang, Y., Han, H., and Shi, H. (2006). Microarray analysis of adipose tissue gene expression profiles between two chicken breeds. J. Biosci. 31, 565–573. doi: 10.1007/bf02708408
Wang, H. B., Li, H., Wang, Q. G., Zhang, X. Y., Wang, S. Z., Wang, Y. X., et al. (2007). Profiling of chicken adipose tissue gene expression by genome array. BMC Genomics 8:193. doi: 10.1186/1471-2164-8-193
Wen, C., Jiang, X., Ding, L., Wang, T., and Zhou, Y. (2017). E?ects of dietary methionine on breast muscle growth, myogenic gene expression and IGF-I signaling in fast-and slow-growing broilers. Sci. Rep. 7:1924. doi: 10.1038/ s41598-017-02142-z
Wensveen, F. M., Valentic,´ S., Šestan, M., Turk Wensveen, T., and Polic,´ B. (2015). The “Big Bang” in obese fat: events initiating obesity-induced adipose tissue inflammation. Eur. J. Immunol. 45, 2446–2456. doi: 10.1002/eji. 201545502
Wideman, R. F., and Hamal, K. R. (2011). Idiopathic pulmonary arterial hypertension: an avian model for plexogenic arteriopathy and serotonergic vasoconstriction. J. Pharmacol. Toxicol. Methods 63, 283–295. doi: 10.1016/j. vascn.2011.01.002
Zambonelli, P., Zappaterra, M., Soglia, F., Petracci, M., Sirri, F., Cavani, C., et al. (2016). Detection of di?erentially expressed genes in broiler pectoralis major muscle a?ected by white striping-wooden breast myopathies. Poult. Sci. 95, 2771–2785. doi: 10.3382/ps/pew268
Zhang, B., and Coon, C. N. (1997). The relationship of various tibia bone measurements in hens. Poult. Sci. 76, 1698–1701. doi: 10.1093/ps/76.12.1698 Zhang, B. B., Zhou, G., and Li, C. (2009). AMPK: an emerging drug target for diabetes and the metabolic syndrome. Cell Metab. 9, 407–416. doi: 10.1016/j. cmet.2009.03.012
Zhuo, Q., Yang, W., Chen, J., and Wang, Y. (2012). Metabolic syndrome meets osteoarthritis. Nat. Rev. Rheumatol. 8, 729–737. doi: 10.1038/nrrheum.2012. 135
Zoico, E., Rossi, A., Di Francesco, V., Sepe, A., Olioso, D., Pizzini, F., et al. (2009). Adipose tissue infiltration in skeletal muscle of healthy elderly men: relationships with body composition, insulin resistance, and inflammation at the systemic and tissue level. J. Gerontol. A Biol. Sci. Med. Sci. 65, 295–299. doi: 10.1093/gerona/glp155















.jpg&w=3840&q=75)