I. INTRODUCTION
There is widespread interest in the successful development of reduced-crude protein (CP) diets, focusing on alternative dietary strategies to reduce CP levels whilst maintaining acceptable growth performance (Greenhalgh et al., 2020). Reduced CP diets have the potential to provide advantages environmentally by decreasing outputs of nitrogen and ammonia, improving bird welfare by enhancing litter quality and lowering incidences of foot-pad dermatitis and improving flock health by lessening undigested protein entering the large intestine to fuel the proliferation of potential pathogens (Greenhalgh et al., 2020).
Additionally, reduced-CP diets provide economic advantages by lessening feed ingredient costs. Modest reductions in CP are already being realised by inclusions of unbound (synthetic or crystalline) methionine, lysine and threonine, which have been routinely included in poultry diets for decades (Kidd et al., 2013). Of considerable interest are branched-chain amino acids (BCAA); isoleucine, leucine and valine as they are regulators of muscle protein synthesis (Yoshizawa., 2004), accounting for 40% of the dietary essential amino acids in body protein (Ospina-Rojas et al., 2017; Ferrando et al., 1995). Additionally, either isoleucine or valine may be the fourth limiting amino acid in reduced-CP diets, but they may also be antagonised by leucine. BCAA antagonisms in poultry were probably first reported by Mathieu and Scott (1968) and subsequently confirmed by Smith and Austic (1978) and other researchers. While BCAA antagonisms are not thought to be an issue in conventional diets (Waldroup et al., 2002), this may not apply to reduced-CP diets. Characterisation of said BCAA antagonisms include depressed feed intake, poor weight gain and increased FCR (Farran et al., 2003).
The Texas A&M optimal digestible amino acid ratios suggest ratios of 69 isoleucine, 109 leucine and 80 valine relative to lysine (100) (Wu, 2014); however, leucine levels in practical broiler diets almost invariably exceed 109 and may be in the order of 130. That requirements for amino acids which are typically present in more than adequate amounts in broiler diets may not be adequately identified was raised by Waldroup (2007) and this caveat certainly applies to leucine.
As an amino acid, leucine is a ‘building-block’ for protein deposition but when plasma and intracellular levels of leucine exceed the minimum needed for protein accretion, the metabolic roles of this amino acid declare themselves (Harper et al., 1984). Given in sufficient amounts, high concentrations of leucine have been shown to activate mTOR signalling, promoting protein deposition as well as supressing protein degradation in skeletal muscle (Li et al., 2011). Thus, there is the implication that broiler chickens may benefit from dietary leucine inclusions that are higher than standard recommendations. However, it follows that this will probably necessitate higher inclusions of isoleucine and valine to counteract antagonistic BCAA interactions (D’Mello & Lewis, 1970). Therefore, the present study was designed to test the hypothesis that elevated leucine inclusions will benefit the performance of broiler chickens offered wheat-based, reduced-CP diets (190 g/kg) from 7 to 28 days post-hatch provided there are concomitant elevations in isoleucine and valine inclusions.
II. MATERIALS AND METHODS
This study fully complied with the guidelines (2019/1667) specifically approved by the Research Integrity and Ethics Administration of The University of Sydney. The experimental design consisted of a 3 × 3 factorial array of dietary treatments, which were offered to 378 offsex male Ross 308 broiler chickens from 7 to 28 days post-hatch. The dietary treatments as shown in Table 1 comprised three inclusion levels of digestible leucine (12.71, 15.02 and17.33 g/kg) and three inclusion levels of combinative, digestible isoleucine and valine (g/kg) (17.21, 20.32 and 23.45 g/kg). All nine diets were wheat-based and formulated to contain 190 g/kg crude protein and 11.55 g/kg digestible lysine with an energy density of 12.90 MJ/kg metabolisable energy (ME), and the dietary electrolyte balance was maintained constant at 230 mEq/kg. Weight gain, feed intake, FCR and abdominal fat-pad weights were determined. The experimental data, as a 3 × 3 factorial array, were subject to analyses of variance using the IBM® SPSS® Statistics 24 program (IBM Corporation. Somers, NY). Experimental units were cage means (7 replicate cages of 6 birds per dietary treatment) and probability levels of less than 5% were considered statistically significant by Student’s t-test.
Table 1- Outline of dietary treatments
III. RESULTS AND DISCUSSION
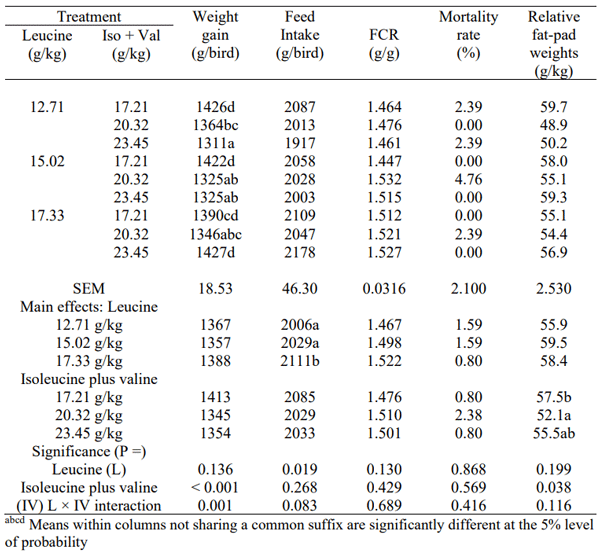
The effects of dietary treatments on growth performance and relative abdominal fat-pad weights are shown in Table 2. A treatment interaction (P = 0.001) between leucine and isoleucine plus valine was observed for weight gain. Weight gains of birds offered diets with digestible leucine contents of 12.71 and 15.02 g/kg declined with increasing isoleucine plus valine concentrations by 8.06% (1311 versus 1426 g/kg; P < 0.001) and 6.82% (1325 versus 1422 g/kg; P = 0.001), respectively. In contrast, at 17.33 g/kg leucine, 23.45 g/kg additional isoleucine plus valine numerically increased weight gains by 2.66% (1427 versus 1390 g/bird P < 0.20), where probability levels are based on pair-wise comparisons. Increasing leucine concentrations significantly (P < 0.02) influenced feed intakes with an increase of 5.23% (2111 versus 2006) following the transition from 12.71 to 17.33 g/kg digestible leucine. These values reflect findings from earlier studies that suggested that inclusions of valine increased efficacy and reduced antagonisms of leucine when included at equally high or higher levels with both leucine and isoleucine as opposed to sub-optimal inclusions. Lower levels of all three BCAA simultaneously have shown no impact, but comparatively fare better than suboptimal inclusions of valine where antagonisms would be more pronounced (D’Mello and Lewis, 1970; Farran and Thomas,1990).
Table 2 - Effects of dietary treatments on weight gains, feed intakes, feed conversion ratios (FCR), mortality rates and relative abdominal fat-pad weights from 7 to 28 days post hatch
FCR was not influenced (P > 0.130) by dietary treatments. The low overall mortality rate of 1.32% was not influenced by treatment (P > 0.40). Isoleucine plus valine concentrations significantly (P < 0.05) influenced relative abdominal fat-pad weights. Indeed, a quadratic relationship (r = 0.310; P = 0.048) between isoleucine plus valine concentrations and relative fat-pad weights was found, where:
y(g/kg) = 240.836 – 18.217 × (Ile + Val)(g/kg) + 0.4398 × (Ile + Val)(g/kg)2 .
It may be deduced from this regression equation that the minimal relative fat-pad weight of 52.2 g/kg would be generated by an isoleucine plus valine concentration of 20.71 g/kg. Reductions in dietary CP often increase bird abdominal fat due to higher starch levels in the diet; however, in a similar study conducted on birds between 21-42 days post hatch, abdominal fat decreased linearly (P< 0.05) with increased valine and leucine (Ospina-Rojas et al., 2017), with leucine decreasing blood triglycerides levels and valine decreasing fatty acid synthesis without stimulating lipid degradation (Ospina-Rojas et al., 2017). The findings of the present study did not meet expected outcomes, with higher inclusions of leucine not altering growth performance in a reduced-CP, wheat-based diet. Further investigation is thus warranted, with adjustments in the inclusion levels of leucine in concomitant with valine and isoleucine to further understand their interactions in the context of reduced-CP diets to improve on broiler performance outcomes.
ACKNOWLEDGEMENTS: The authors would like to acknowledge the financial support and guidance of The AgriFutures Australia Chicken Meat Program and the technicians that have contributed to the data used to produce this paper.
Presented at the 32th Annual Australian Poultry Science Symposium 2021. For information on the next edition, click here. 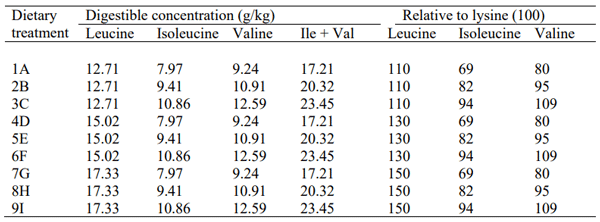











.jpg&w=3840&q=75)