Effect of SQM Iron in Broilers Under Microbial Exposure
Published: September 11, 2019
By: Dr. Jack Garrett / Ph.D., Director of Technical Services & Research at QualiTech.
Iron plays an important role in poultry nutrition. From the active site in the heme group in blood to functional groups in enzymes, iron is critical for metabolic balance. These roles for iron aren’t unique to animals, as it is also important in the growth of bacteria, including pathogens. Also, iron is a significant antagonist against minerals, vitamins and other nutrients in diets. The right balance of active and available iron is critical to optimize broiler growth. Several trials have been conducted recently with SQM® Iron which showed overall significant influence on broiler performance*. In an attempt to determine a potential SQM Iron mode of action with regards to microbial populations, a microbial exposure study was conducted.
A total of 2,496 mixed sex broiler chicks were obtained from a commercial hatchery. Broilers were randomly assigned to one of 12 pens per treatment containing 52 healthy/viable broiler mixed-sexes per pen for a total of 624 broilers per treatment group. Broilers were fed ad libitum their respective treatment from Trial Day 0 to 42 days of age.
Description of Microbial Exposure: The following system was employed throughout the entire 0 to 42-day live performance study.
- Coccidial protection: Cocci-Vac was administered to the birds at the hatchery on Day 0. No other protection was provided.
- The sub clinical exposure (built-up litter and moderate stress conditions via bacterial and coccidial exposure), relating to age of litter (a minimum of three previous flocks), was obtained with built-up litter, containing a mixture of coccidia E. acervulina (>50,000 oocysts/bird), Clostridium perfringens (>104 per bird) and E. coli (106 per bird) bacteria were administered into the litter on Day 7.
Each experimental test unit of broiler mixed-sex chickens were housed in separated pens, located in a room containing forced air heaters with a cross-house precision-controlled ventilation system. Broilers were placed in a 5 ft x 10 ft pen floor area with a minimum of 0.85 ft2 per bird. Water was provided by at least two nipple drinkers per pen (via well water). Continuous (24 hr) use of incandescent lights were used during the entire study. Full lighting of >3 fc was used the first week, and then dimmed to 1 fc for the next two weeks of age. Three diets (corn-soy based) were formulated per iron source (Prestarter 0-7 day, Grower 8-35 day and Finisher 36-42 day). Nutrient composition was ME (kcal/kg) 3417, 3086, and 3197; CP % 20, 18, and 17; lysine % 1.45, 1.20, 1.02; methionine % min 0.6, 0.5 and 0.4. The level of iron fortification as 20 ppm for all diets using either iron sulfate or SQM Iron. Live performance body weights and feed intakes were collected on Day 0, 14, 21, 35 and 42 during the growing period. Weight gain, feed intake, feed:gain ratio (feed efficiency) were calculated for 0-21, 0-35, 0-42, 15-21, 22- 35 and 36-42 days of age. Lesion scores were collected at 21 and 42 days of age. Data was analyzed as a 2x2 factorial arrangement of treatments (iron source x microbial exposure). The results of this study are shown in Table 1.
There were no interactions between the iron source and the microbial exposure for growth performance parameters. The only significant interaction was in mortality, starting around day 21 and continuing to the end of the study. Those birds receiving SQM Iron had reduced mortality (p<0.05) compared to the sulfate birds in the exposure treatment. For all of the performance parameters, there were either significant differences for improved performance or strong trends for the SQM Iron birds compared to the birds receiving iron sulfate. The performance of the exposed birds receiving SQM Iron closely approached that of the unexposed iron sulfate birds. This indicates that SQM Iron was reducing the impact that pathogens had on performance within the exposure groups, as well as under a subclinical situation in the unexposed birds. This protection effect can be observed in the microbial counts in Figures 1 and 2. While there was no difference in the E. coli levels, those birds receiving SQM Iron had reduced counts of C. perfringens and Eimeria in both exposure treatments relative to iron sulfate.
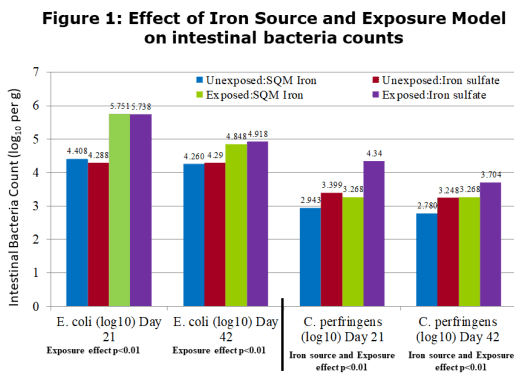
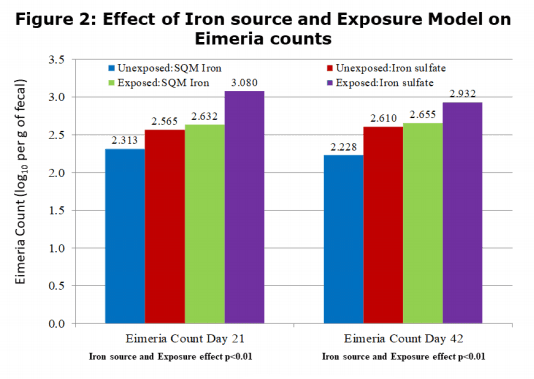
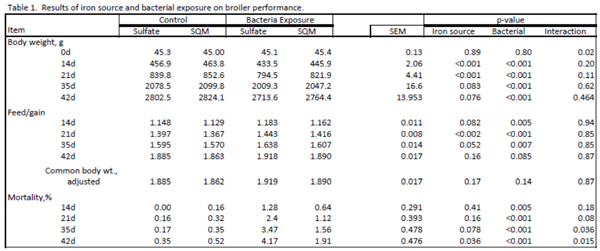
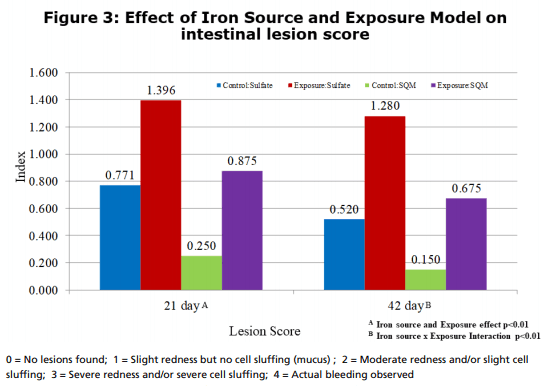
Figure 3 shows the effect on lesion scores observed in the study at 21 and 42 days. There was a significant (p<0.01) reduction in the lesion scoring at 21 days due to the feeding of SQM iron vs iron sulfate and due to the microbial exposure. At 42 days, there was a significant (p<0.01) interaction where the feeding of SQM iron not only reduced the lesion score but also reduced the severity of the lesions under a exposure situation.
Conclusion
The results of this study further confirmed the benefit of feeding SQM Iron over iron sulfate that had been observed in previous research*. The influence of iron on pathogenic growth varied across microbial species evaluated in this study. E. coli counts were not influenced by iron source, and had similar loads within unexposed and exposure treatments. However, Clostridium perfringens counts, as well as Eimeria, counts, were significantly reduced by SQM Iron relative to iron sulfate in both exposed and unexposed groups. Notably, the level of infection during the exposure was reduced by SQM Iron to the level observed in the unexposed sulfate treatment. This reduction could be a good explanation for the overall significant improvement in performance observed with SQM Iron birds. Other research in the literature has indicated that the form of iron (either ferric or ferrous) can have an influence on the proliferation of pathogenic bacteria, which require iron for growth. With an organic iron source, such as SQM Iron, it appears that the improved iron availability to the host animal, and consequent reduction in the amount of iron available to the microbes, would be just as effective in decreasing overall pathogen numbers. This combined action can explain the benefits of SQM Iron in helping improve broiler performance.
Related topics
Authors:

Influencers who recommended :
Maqsood JafferyJoin to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post
21 de septiembre de 2019
what does it mean SQM IRON







You may be interested in
.jpg&w=3840&q=75)