Introduction
Serovars of Salmonellaenterica remain among the most important foodborne pathogens worldwide due to a significant number of human illnesses reported (Scallan et al., 2011). There are an extensive number of animals that serve as hosts for the members of this genus and are able to spread these agents to animal and human populations; however, salmonellosis in humans is most frequently associated with the consumption of contaminated fresh and processed poultry products (Lynch et al., 2006; Foley et al., 2011). According to Foley et al. (2011), Salmonella Typhimurium continues to be among the most common serovars isolated from poultry and a common cause of human salmonellosis. Furthermore, public concern associated with antibiotic-resistant strains is challenging the poultry industry to find alternative means of control and consequently, continuous studies on alternative methods to control foodborne pathogens are necessary (Boyle et al., 2007; McNulty et al., 2007). Chitosan is a biocompatible polymer derived by deacetylation of chitin from shellfish, and its use in industry, agriculture, and medicine is well described (Rabea et al., 2003; Senel and McClure, 2004; Friedman and Juneja, 2010). The interest in chitosan as a biological sanitizer arises from several studies reporting its antimicrobial and antioxidative effects in foods (No et al., 2002; Friedman and Juneja, 2010). The antimicrobial activities of chitosan against foodborne pathogens have been extensively investigated in the food industry (Singla and Chawla, 2001; No et al., 2002; Senel and McClure, 2004; Petrovich et al., 2008; El Hadrami et al., 2010; Kong et al., 2010; Vargas and Gonzalez-Martinez, 2010). However, to the best of our knowledge, nothing is yet known on the effect of chitosan against Salmonella in poultry. Therefore, this study was designed to evaluate the effect of dietary chitosan on ante mortem control of Salmonellaenterica serovar Typhimurium (ST) recovery via in vitro crop assays and infection and horizontal transmission in broiler chickens.
Materials and Methods
Chitosan
Deacetylated 95% food-grade chitosan was obtained commercially (Paragon Specialty Products, LLC, Rainsville, AL) and was used in all experiments. The chitosan molecular weight was 350 kDa with viscosity of 800 mPa, and particle size of 100 US mesh (sieve size 0.152 mm). For in vitro crop assay experiments, 0.2% (wt/vol) chitosan was prepared by dissolving it in a solution containing 0.2% (vol/vol) acetic acid. Further dilutions were made in sterile distilled water.
Animal source and diet
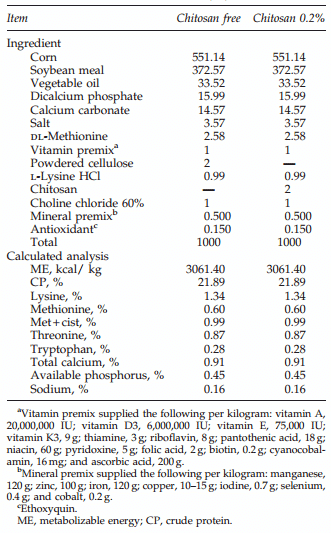
Day-of-hatch, off-sex broiler chickens were obtained from Cobb-Vantress (Siloam Springs, AR) for all the trials mentioned below. All animal-handling procedures were in compliance with Institutional Animal Care and Use Committee at the University of Arkansas. In all experiments, diets were fed in mash form, and were formulated to meet or exceed National Research Council (NRC, 1994) estimated nutrient requirements. The common starter diet was a typical corn soybean-meal diet (chemical analysis of nutrients is presented in Table 1). The diet with chitosan was similar to the common starter diet but was supplemented with 0.2% chitosan.
Table 1. Composition of the Starter Diet for Broiler Chickens (kg)
Table 2. Evaluation of Different Concentrations of Chitosan in Inhibiting Salmonella Typhimurium Growth in an In Vitro Crop Assay
Bacterial strain and culture conditions
A poultry strain of Salmonella Typhimurium, selected for resistance to nalidixic acid (NA, Catalog no. N-4382; Sigma, St. Louis, MO), was used for all experiments. The amplification and enumeration procedure for this strain has been described previously (Tellez et al., 1993). For these experiments, ST was grown in tryptic soy broth (Catalog no. 22092; Sigma, St. Louis, MO) for approximately 8 h. The cells were washed three times with 0.9% sterile saline by centrifugation (1864 X g), and the approximate concentration of the stock solution was determined spectrophotometrically at 625 nm. The stock solution was serially diluted and confirmed by colony counts of three replicate samples (0.1 mL/replicate) spread plated on brilliant green agar (BGA, Catalog No. 278820; Becton Dickinson, Sparks, MD) plates containing 25 mg/mL novobiocin (NO, Catalog No. N-1628, Sigma) and 20 mg/mL NA. For all experiments, ST recovery was completed on BGA plates containing NO and NA at these concentrations. The CFUs of ST for inoculation were determined by spread plating and are reported as concentration of CFU/mL for in vitro experiments and total CFU/bird for in vivo challenge experiments.
In vitro crop assays
An assay previously described (Barnhart et al., 1999) was used with slight modifications. Briefly, 1.25 g of unmedicated chick starter feed was measured and placed into 13 X 100-mm borosilicate tubes and sterilized by autoclaving. The feed was suspended in 5 mL sterile saline as a control or 5 mL of a 0.05%, 0.1%, or 0.2% chitosan solution. Tubes were inoculated with a ST culture at a final concentration of approximately 106 CFU/mL. Each treatment had five replicates. After administering the treatment, the tubes were agitated via vortex stirring and incubated at 37°C for 6 h. The tubes were then agitated, and 20 lL of the content was serially diluted and plated in triplicate on BGA containing NO and NA. Typical ST colonies were counted after 24 h of incubation. A second experiment to measure the effects of 0.2% chitosan on recovery of ST was conducted following the same experimental procedures, but with sampling at 0.5 and 6 h of incubation at 37C. This assay was repeated for three replicate trials. Additionally, a trial to compare the effects of chitosan solution (chitosan 0.2% and acetic acid 0.2%) and acetic acid (0.2%) on Salmonella Typhimurium was conducted and followed the same procedures.
Table 3. Effect of 0.2% Chitosan on Levels of Salmonella Typhimurium Recovery in an In Vitro Crop Assay
in vivo experimental design
On the day of placement for each trial, 10 chicks were harvested for evaluation of wild-type Salmonella spp. infection. Chicks were humanely killed by CO2 inhalation; ceca and cecal tonsils, liver, and spleen were aseptically removed, enriched in tetrathionate broth (Catalog no. 210420, Becton Dickinson), and plated on BGA containing 25 lg/mL of NO.
In two direct-challenge trials, day-of-hatch chickens were randomly assigned to untreated control diet (n = 20) or dietary treatment with 0.2% chitosan (n = 20) for 7 days. Chicks were housed in brooder batteries with feed and water provided ad libitum. On day 4, all chicks were challenged with 2 · 105 CFU ST/bird. At 7 days, chicks were humanely killed by CO2 inhalation, and both ceca and cecal tonsils were aseptically collected and cultured for ST. A third experiment to evaluate effect of 0.2% dietary chitosan on ST horizontal transmission was done. Chickens were randomly assigned to untreated control diet (n = 50) or treatment with 0.2% chitosan (n = 50) for 10 days. Chicks were housed in floor pens with feed and water provided ad libitum. On day 3, 10 birds per group were challenged with 105 CFU ST/bird to act as seeders, while the other 40 birds per group were contacts challenged by horizontal transmission from the seeders. At 10 days of age, 10 seeders and 20 contact chicks were humanely killed by CO2 inhalation; ceca and cecal tonsils were aseptically harvested and cultured for ST recovery.
For all experiments, cecal tonsils were enriched in 10 mL of tetrathionate broth overnight at 37 C. Following enrichment, each sample was streaked for isolation on BGA. The plates were incubated at 37C for 24 h and examined for the presence or absence of colonies typical of antibiotic resistant ST. For direct plating to determine ST/g of cecal contents, ceca were weighed and homogenized in sterile sample bags (Catalog No. B00679WA; Nasco, Fort Atkinson, WI) using a rubber mallet. Sterile saline (4X weight:volume) was added to each sample bag containing homogenized cecal contents and hand stomached. Serial dilutions were spread plated on BGA; the plates were incubated at 37°C for 24 h and colonies typical of antibiotic-resistant ST were counted. All animal-handling procedures were in compliance with the Institutional Animal Care and Use Committee at the University of Arkansas.
Table 4. Effect of 0.2% Acetic Acid and 0.2% Chitosan Solution on Levels of Salmonella Typhimurium Recovery in an In Vitro Crop Assay
Statistical analysis
In vitro crop assay and cecal CFU data were converted to log10 and compared using the GLM procedure of SAS (SAS Institute, 2002) with significance reported at p < 0.05. The incidence of ST recovery within experiments was compared using the chi-square test of independence (Zar, 1984) to determine significant ( p < 0.05) differences between control and treated groups.
Results and Discussion
All chitosan concentrations significantly reduced the total recovered CFU of ST from crop assays (Table 2). However, the concentration of 0.2% showed a marked reduction of more than 2.5 log10, and it was selected for further in vitro and in vivo evaluations. Chitosan at a concentration of 0.2% significantly reduced the total recovered CFU of ST at both 0.5 and 6 h postinoculation when compared with control in all three additional crop assay trials (Table 3). When comparing the chitosan solution containing acetic acid with the acetic acid solution, the results showed that at 0.5 h only the chitosan/acetic acid solution was able to significantly reduce ST levels. In the 6 h of incubation, acetic acid solution reduced ST levels in 2.29 log10 and chitosan/acetic acid solution reduced ST by 3.24 log10, which shows a synergistic effect of the acetic acid with chitosan (Table 4). However, it has been reported that acetic acid at the concentrations of 0.01, 0.1, and 1% are ineffective in reducing Salmonella Enteritidis levels in an in vitro crop assay (Barnhart et al., 1999).
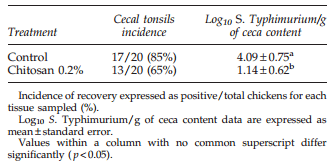
For all in vivo trials, chicks were negative for wild-type Salmonella spp. at placement. The effect of dietary chitosan on ST intestinal colonization at 7 days of age in broiler chickens is described in Table 5. Dietary 0.2% chitosan significantly reduced CFU/g of ST recovered from the ceca in both experiments. However, no significant reduction in the incidence of ST from cecal tonsils was observed. In the ST horizontal transmission experiment, seeder chicks were harvested and 100% ST infection was confirmed for both control and treated chickens (data not shown). For contact chicks, dietary 0.2% chitosan significantly reduced the CFU/g of ST recovered from the ceca (Table 6), showing a reduction in the horizontal transmission of ST in birds treated with dietary chitosan.
Table 5. Effect of 0.2% Dietary Chitosan on Incidence and Levels (Colony-Forming Units/g) of Salmonella Typhimurium Recovery at 7 Days of Age in Broiler Chickens
Chitosan is a molecule that has antimicrobial activity against many Gram-negative and Gram-positive bacteria (Rabea et al., 2003; Zheng and Zhu, 2003; Ganan et al., 2009; Friedman and Juneja, 2010; Batista et al., 2011; Islam et al., 2011). For example, Lee et al. (2009) demonstrated both in vitro and in vivo (mice) that chitosan oligosaccharides have antibacterial effect on the Gram-negative bacterium Vibrio vulnificus, which causes sepsis and gastrointestinal illness in humans. However, most of the studies are related to the in vitro effect of chitosan in reducing bacteria, not considering its effects in the presence of organic matter and more importantly under in vivo conditions. Moreover, to our knowledge, there are no previous reports in the literature about the effect of chitosan on Salmonella in poultry.
The mechanism of antimicrobial activity of chitosan has not yet been fully elucidated, and different hypotheses have been proposed. A common hypothesis is alteration of cell permeability due to interactions between the positive charge of chitosan molecules (amino group at C-2) and the negative charge of bacterial cell membranes (Helander et al., 2001; No et al., 2007; Friedman and Juneja, 2010). Additionally, chelation of metals and essential nutrients by chitosan molecules has been hypothesized to inhibit bacterial growth (Rabea et al., 2003). Zheng and Zhu (2003) also suggested that high-molecular-weight chitosan could be able to form a polymer membrane around the bacterial cell, preventing it from receiving nutrients. They also proposed that low-molecularweight chitosan could move into cells through pervasion, and disrupt the physiological activities of the bacterial cell (Zheng and Zhu, 2003).
Chitin and chitin derivatives (chitosan and chitosan oligosaccharides, for example) have also been shown to have an effect on innate and adaptive immune responses such as activation of innate immune cells and induction of cytokine and chemokine production (Lee et al., 2008; Lee et al., 2009). According to Bueter et al. (2013), the chitosan particles are recognized via specific receptor(s) and phagocytosed, inducing a response, which is not all defined, and up-regulating the innate immune system in mammals. It has been suggested that dietary oligochitosan can act as a prebiotic in chickens (Huang et al., 2005, 2007). Huang et al. (2007) suggested that the prebiotic effect of chitosan could be related to a chitosan attachment to the bacteria, leading to an immune response to this antigen, or by direct stimulation of the immune system. Chitosan could be also improving nutrients utilization by the host and/or facilitating beneficial bacteria growth (Huang et al., 2007). Wang et al. (2003) described a reduction in Escherichia coli recovered from the ceca of chicks treated with 0.1% of oligochitosan in the feed, and also an improvement of small intestine microvilli density and growth performance. Dietary oligochitosan has been related to an increase of ileal digestibility of nutrients and performance improvement in broiler chickens (Huang et al., 2005). According to Huang et al. (2007), dietary supplementation of oligochitosan improved serum levels of immunoglobulin A (IgA), IgG, and IgM, suggesting that this increase may be related to cytokine production stimulation. An increase of the relative weight of spleen, bursa, and thymus in broilers after oligochitosan supplementation has also been reported, suggesting an improvement in the immune response (Huang et al., 2007; Deng et al., 2008).
Table 6. Effect of Chitosan on Horizontal Transmission of Salmonella Typhimurium in Broiler Chickens
Conclusions
The in vitro studies were designed to mimic conditions in the crop of birds because colonization of Salmonella in this organ is a persistent problem associated with processing-age birds (Byrd et al., 1998a, 1998b). In vivo reduction in cecal Salmonella Typhimurium may decrease the overall pathogen load in birds, making them less likely to spread the infection further. Overall, the addition of 0.2% chitosan in the diet was able to reduce colonization of ST in broiler chicks. In the present study, the bactericidal activity of dietary 0.2% chitosan was able to significantly reduce ST both in vitro and in vivo, thus proving to be an alternative tool to reduce crop, ceca, and consequently carcass ST contamination as well as decreasing the amount of ST shed to the environment. Further studies are in progress to evaluate the additive or synergistic effects of chitosan with other natural agents and compounds such as direct-fed microbials.
Disclosure Statement
No competing financial interests exist
References
Batista ACL, Dantas GC, Santos J, Amorim RVS. Antimicrobial effects of native chitosan against opportunistic Gram-negative bacteria. Microbiol J 2011;1:105–112.
Barnhart ET, Sarlin LL, Caldwell DJ, Byrd JA, Corrier DE, Hargis BM. Evaluation of potential disinfectants for preslaughter broiler crop decontamination. Poult Sci 1999;78:32–37.
Boyle EC, Bishop JL, Grassl GA, Finlay BB. Salmonella: From pathogenesis to therapeutics. J Bacteriol 2007;189:1489–1495.
Bueter CL, Specht CA, Levitz SM. Innate sensing of chitin and chitosan. PLoS Pathog 2013;9:e1003080.
Byrd JA, Corrier DE, Hume ME, Bailey RH, Stanker LH, Hargis BM. Effect of feed withdrawal on Campylobacter in the crops of market-age broiler chickens. Avian Dis 1998a;42:802–806.
Byrd JA, Corrier DE, Hume ME, Bailey RH, Stanker LH, Hargis BM. Incidence of Campylobacter in crops of preharvest marketage broiler chickens. Poult Sci 1998b;77:1303–1305.
Deng X, Li X, Liu P, Yuan S, Zang J, Li S, Piao X. Effect of chitooligosaccharide supplementation on immunity in broiler chickens. J Anim Sci 2008;21:1651–1658.
El Hadrami A, Adam LR, El Hadrami I, Daayf F. Chitosan in plant protection. Mar Drugs 2010;8:968–987.
Foley SL, Nayak R, Hanning IB, Johnson TJ, Han J, Ricke SC. Population dynamics of Salmonella enterica serotypes in commercial egg and poultry production. Appl Environ Microbiol 2011;77:4273–4279.
Friedman M, Juneja VK. Review of antimicrobial and antioxidative activities of chitosans in food. J Food Prot 2010;73: 1737–1761.
Ganan M, Carrascosa AV, Martinez-Rodriguez AJ. Antimicrobial activity of chitosan against Campylobacter spp. and other microorganisms and its mechanism of action. J Food Prot 2009;72:1735–1738.
Helander IM, Nurmiaho-Lassila EL, Ahvenainen R, Rhoades J, Roller S. Chitosan disrupts the barrier properties of the outer membrane of Gram-negative bacteria. Int J Food Microbiol 2001;71:235–244.
Huang RL, Yin YL, Wu GY, Zhang YG, Li TJ, Li LL, Li MX, Tang ZR, Zhang J, Wang B, He JH, Nie XZ. Effect of dietary oligochitosan supplementation on ileal digestibility of nutrients and performance in broilers. Poult Sci 2005;84:1383–1388.
Huang RL, Deng ZY, Yang C, Yin YL, Xie MY, Wu GY, Li TJ, Li LL, Tang ZR, Kang P, Hou ZP, Deng D, Xiang H, Kong XF, Guo YM. Dietary oligochitosan supplementation enhances immune status of broilers. J Sci Food Agric 2007;87:153–159.
Islam MDM, Masum SMD, Mahbub KR. In vitro antibacterial activity of shrimp chitosan against Salmonella paratyphi and Staphylococcus aureus. J Bangladesh Chem Soc 2011;24:185–190.
Kong M, Chen XG, Xing K, Park HJ. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int J Food Microbiol 2010;144:51–63.
Lee CG, Silva CA, Lee JY, Hartl D, Elias JA. Chitin regulation of immune responses: An old molecule with new roles. Curr Opin Immunol 2008;20:684–689.
Lee BC, Kim MS, Choi SH, Kim KY, Kim TS. In vitro and in vivo antimicrobial activity of water-soluble chitosan oligosaccharides against Vibrio vulnificus. Int J Mol Med 2009;24:327–333.
Lynch M, Painter J, Woodruff R, Braden C. Centers for Disease Control and Prevention. Surveillance for foodborne-disease outbreaks United States, 1998–2002.
MMWR Surveill Summ 2006;55:1–42. McNulty CA, Boyle P, Nichols T, Clappison P, Davey P. The public’s attitudes to and compliance with antibiotics. J Antimicrob Chemother 2007;60(Suppl 1):i63–i68.
[NRC] National Research Council. Nutrient Requirements of Poultry, 9th rev. ed. Washington, DC: National Academy Press, 1994.
No HK, Park NY, Lee SH, Meyers SP. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int J Food Microbiol 2002;74:65–72.
No HK, Meyers SP, Prinyawiwatkul W, Xu Z. Applications of chitosan for improvement of quality and shelf life of foods: A review. J Food Sci 2007;72:R87–R100.
Petrovich I, Grigor’iants LA, Gurin AN, Gurin NA. Chitosan: Structure, properties, use in medicine and stomatology. Stomatologiia (Mosk) 2008;87:72–77.
Rabea EI, Badawy MET, Stevens CV, Smagghe G, Steurbaut W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003;4:1457–1465.
SAS Institute. SAS User Guide. Version 9.1. Cary, NC: SAS Institute Inc., 2002. Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson M, Roy SL, Jones JL, Griffin PM. Foodborne illness acquired in the United States—Major pathogens. Emerg Infect Dis 2011;17:7–15.
Senel S, McClure SJ. Potential applications of chitosan in veterinary medicine. Adv Drug Deliv Rev 2004;56:1467–1480. Singla AK, Chawla M. Chitosan: Some pharmaceutical and biological aspects—An update. J Pharm Pharmacol 2001;53: 1047–1067.
Tellez G, Dean CE, Corrier DE, Deloach JR, Jaeger L, Hargis BM. Effect of dietary lactose on cecal morphology, pH, organic acids, and Salmonella enteritidis organ invasion in Leghorn chicks. Poult Sci 1993;72:636–642.
Vargas M, Gonzalez-Martinez C. Recent patents on food applications of chitosan. Recent Pat Food Nutr Agric 2010;2:121–128.
Wang XW, Du YG, Bai XF, Li SG. The effect of oligochitosan on broiler gut flora, microvilli density, immune function and growth performance. Acta Zoonutrim Sin 2003;15:32–45.
Zar JH. Biostatistical Analysis, 2nd ed. Upper Saddle River, NJ: Prentice-Hall, 1984. Zheng LY, Zhu JF. Study on antimicrobial activity of chitosan with different molecular weights. Carbohydr Polym 2003;54: 527–530.













.jpg&w=3840&q=75)