INTRODUCTION
In current poultry production, physical separation between the hatchery and farms can cause newborn chickens to spend a variable period of time without food, and usually between 36 to 48 hrs. pass after birth before birds have access to feed, reducing the weight of the birds during this time (1). Delayed feed consumption can reduce the rate of growth and cause weight loss (2).
By contrast, immediate access to food and water after hatching may promote development of the digestive tract, increase body weight (3.4) and reduce mortality in the first 3 days of life (5).
With early feeding, nutrients necessary for infants are supplied during the first 48-72 hrs. It has been reported that this can reduce dehydration and promote rapid reabsorption of the yolk sac (6), the development of the liver, pancreas and intestine (7), increase the bird’s immediate development post-hatch, improve young chickens’ resistance against exposure to low temperatures (8) and promote further development of the immune system (9).
When studying the effect of feeding immediately after hatching on activation of the genetic program that regulates hepatic lipogenesis, it was found that during the first week after hatch regulating key lipogenic genes was significant, including ATP citrate lyase enzymes, malic enzyme, fatty acid synthase, acetyl-CoAcarboxilasa, alpha stearyl-CoA desaturase-1, etc. (10). Food restriction during the first 48 hrs. post-hatching caused a significant decrease in weight gain, plasma glucose and the regulation of lipogenic genes, which reverts to the beginning of feeding (10). There are no previous reports about how digestive enzyme activity can be affected, mainly those that are an integral part of the intestinal brush border, such as disaccharidases, phosphatases and phytase.
It is of practical interest to study nutritional requirements and effects of early feeding in birds at an early age, or in the period of time from hatching to their arrival at the farm, on the development of the gastrointestinal tract, as it can affect body development. Therefore, this research was conducted in order to evaluate the effect of providing two different supplements at an early age on the activity of pancreatic and duodenal digestive enzymes in broiler chickens.
MATERIALS AND METHODS
Incubation and hatching of fertile eggs.
A total of 300 fertile eggs from the Hubbard breeder line of hens, 33 weeks old, purchased from the Vipraca Farm, CA located in the state of Yaracuy, were incubated at 37.8°C in a Chick master hatchery with automatic dump from the Nutritional Biochemistry Center of the Faculty of Veterinary Science (FVS) at the Central University of Venezuela (CUV) located 67°36’36’’ east longitude, 10°16’20’’ north latitude and 443 meters above sea level, with an average temperature of 25.1°C and an average annual rainfall of 1063 mm.
At hatching, the chicks were removed from the hatcher, assigned to metal battery cages for their accommodation in the Department of Biochemistry’s animal facility at the FVS-CUV, maintained in suitable conditions of hydration, ventilation and density, and immediately subjected to experimental treatments (100 birds/treatment). The food and water were provided ad libitum from hatching to 72 hours old.
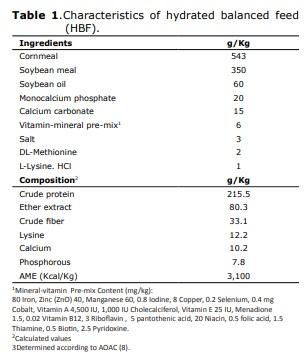
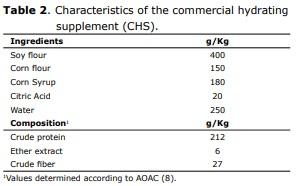
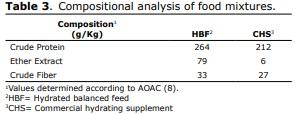
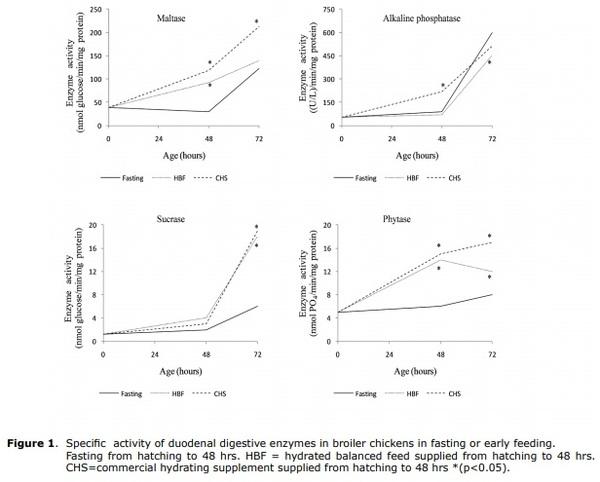
Management of birds and feeding. The hydrated balanced feed was formulated in accordance with international recommendations (11). In Tables 1, 2 and 3, the composition and compositional analysis are shown (12) for the food and food supplements provided. All birds received water ad libitum. In order to take their measurements, at birth and at 48 and 72 hrs. of age, they were sacrificed by cervical dislocation, 30 chicks/treatment/interval to extract, at temperatures between 0 and 4°C, the duodenal segments and pancreas from which the homogenates were prepared to determine intestinal enzyme activity for maltase, sucrase, alkaline phosphatase and phytase. In duodenal homogenate samples, pancreatic enzyme activity was determined for α-amylase, trypsin and the activity of lipase was determined in samples of pancreatic homogenate. International ethical standards for animal research were respected, avoiding the provocation of pain or suffering.
Experimental design and treatment. Birds were assigned in a completely randomized design
to the following treatments:
T0: Fasting from hatching to 48 hrs.
T1:Hydrated balanced food (HBF) from hatching to 48 hrs. of life.
T3: Commercial Hydrating Supplement (CHS) from hatching to 48 hrs. of life.
Compositional analysis of food. The compositional analyzes were performed in a commercial laboratory, and included the determination of crude protein, crude fat and crude fiber, according to standardized methods (12).
Preparation of intestinal homogenates. Duodenal segments were extracted at temperatures between 0 and 4°C, cut longitudinally, laid on a glass plate and gently scraped with a glass slide to obtain the mucous that was homogenized at maximum speed for 1 min in buffer with 2 mM HEPES pH 7.1 and stored in aliquots at -20°C until analysis, within a period not exceeding 15 days.
Pancreatic homogenate preparation. The entire procedure was conducted between 0 and 4°C. The pancreas were homogenized at maximum speed for 3 min at 4 g/ml of physiological saline, filtered through a sterile cotton cloth and the extract was stored in aliquots at -20°C until they were required for enzymatic determinations, within a period no longer than 15 days.
Enzymatic activities. Enzyme analyzes were performed at the Enzymology and Toxicology Laboratory attached to the Department of Biochemistry, FVS-CUV. Homogenized composite replicas were analyzed in all cases, with a pool of 3 chicks for a total of 10 determinations/treatment in each time interval.
Maltase (EC 3 .2 .1 .20 ) and Sucrase (EC 3.2.1.48). The specific activity of the disaccharidases maltase and sucrase was determined using 56 mM maltose or sucrose as substrate in 100 mM sodium maleate, pH 6.0 and dissolved in 15% NaOH (w / v) (13). The reaction mixture was incubated at 41°C, for 15 and 30 min for maltase and sucrase respectively, after which stopped by placing the tubes in boiling water for 20 sec. The amount of glucose released in the reaction was quantified using a GOD-PAP kit 900® Bioscience. The absorbance was measured in a spectrophotometer (Perkin Elmer, Lambda 11, California, USA) at a wavelength of 510 nm. The specific activity was expressed as nmoles of glucose released/min/mg protein (13).
Alkaline phosphatase (EC 3.1.3.1). The activity of this enzyme was determined using LABTEST® Kits. The alkaline phosphatase present in the sample hydrolyzes the substrate thymolphthalein monophosphate, releasing thymolphthalein and inorganic phosphate. 50 μl of substrate, 500 μl of buffer supplied by the manufacturer and 50 μl of the sample were incubated for 10 min at 41°C. The reaction was stopped with a solution containing 150 mM sodium carbonate and 100 mM sodium hydroxide. Absorbance was measured in a spectrophotometer at a wavelength of 590 nm and compared against a known concentration standard. The activity was expressed in (Units/L)/ min/mg protein (13).
Phytase (EC 3.1.3.8). Determined at 41°C by incubating 40 μl of duodenal homogenate with 200 μl of 1 mM inositolhexaphosphoric sodium salt and 200 μl of 25 mM MgCl2 and 50 mM MES buffer pH 6.0. The reaction was stopped by adding 3600 ml of a solution containing 0.28% (w/v) ammonium molybdate, 1.1% Sodium dodecyl sulfate (w/v), and 1.1% ascorbic acid (w/v). The concentration of phosphate released was determined in a spectrophotometer at a wavelength of 820 nm. Absorbance values were compared to a standard curve prepared from a 0.5 mM KH2PO4 solution and expressed as nmoles of PO4 hydrolyzed/min/mg protein (13,14).
α-Amylase (EC 3.2.1.1). The homogenate (50 μl) was incubated for 15 min at 41°C in a reaction mixture containing 100 μl of 160 mM Tris-HCl pH 7.5 and 200 μl of 0.15% starch in saturated sorbic acid. The reaction was stopped by adding 100 μL of 0.5N HCl after which 2 μl of distilled water was added along with 100 μl of 0.01N iodine solution. The amount of starch degraded by amylase was determined spectrophotometrically by measuring the decrease of blue color of the starch-iodine complexion at a wavelength of 620 nm (15). The values were compared to a standard curve prepared from a 0.15% starch solution (w/v). The activity was expressed as μg of starch degraded/min/ mg protein (15).
Trypsin (EC 3.4.21.4). Pancreatic homogenate was incubated for 15 min at 41°C with 200 μl of 1% (w/v) casein as substrate in 0.1 M sodium phosphate pH 7.4. The reaction was stopped by adding 1 μl of 5% (w/v) trichloroacetic acid and the mixture was filtered on 42 Whatman paper. 250 μl of the filtrate reacted with 1250 μl of 0.2 M Na2CO3 and 250 μl Folin-Ciocalteu reagent diluted 1:5 in distilled water (16). After incubation for 20 min at 41°C absorbance was measured at a wavelength of 625 nm. The activity was expressed as Units of Proteolytic Activity (UPA) defined as the amount (mEq) of amino acid tyrosine released as a result of proteolysis of the substrate in the experiment conditions (15).
Lipase (EC 3.1.1.3). The activity of this enzyme was determined in pancreatic homogenate, determining the volume of 0.05M NaOH required to neutralize the fatty acids released during 3 hrs. of incubation, with gentle stirring, at 37°C with 3 ml of olive oil used as substrate. The activity was expressed as ml of NaOH/mg protein (16).
Protein Analysis. Protein content was determined by reacting the samples with a filtered solution on 40 Whatman paper containing 1% Coomassie Brilliant Blue G250 in ethanol, diluted 1:2 in 8.9% orthophosphoric acid (13) using the corresponding buffer as a blank and Bovine Serum Albumin (BSA) as a standard.
Statistical analysis. The results were expressed as the mean ± standard deviation and statistically analyzed through a Kruskal-Wallis analysis and the average range tests at a significant level of p≤0.05 (17).
RESULTS
Maltase (EC 3.2.1.20). Immediately after hatching, the specific activity of maltase enzyme determined in homogenate of duodenal segments was 39 ± 1 nmoles of glucose/min/mg protein. At both 48 and 72 hours old, the activity was higher (p≤0.05) in animals receiving CHS from birth until 48 hrs. post-hatching (T2) in comparison with the control, which was kept in fasting for 48 hrs. (Figure 1). Birds receiving HBF showed higher activity of this enzyme at 48 hrs. old (p≤0.05).
Sucrase (EC 3.2.1.48). Upon hatching, the specific activity of sucrase, determined in the homogenate of duodenal segments was 1.2±0.1 nmoles glucose/min/mg protein, about 32 times lower than the maltase activity after hatching. In the first 48 hrs. after hatching there were no significant differences between treatments. However, at 72 hrs. there were significant differences in favor of T1 and T2 (Figure 1). Therefore, supplying HBF and CHS in the first 48 hrs. of life improved sucrase activity in birds that were 72 hrs. old (p≤0.05), leading to increased activity about 3-fold compared to the control.
Alkaline phosphatase (EC 3.1.3.1). Immediately after hatching, the specific activity of the alkaline phosphatase enzyme, determined in homogenate of duodenal segments was 53 ± 12 (U/L)/min/mg protein. At 48 hrs. of life, animals that consumed CHS (T2) had 1.2 times higher activity, with significant differences (p<0.05) compared to the control group (T0) (Figure 1). However, in birds that fasted for 48 hrs. after hatching (T0) activity significantly increased (p≤0.05) on the third day, 24 hrs. after receiving food.
Phytase (EC 3.1.3.8). Immediately after hatching, the specific activity of the phytase enzyme, determined in homogenate of duodenal segments was 5.1 ± 0.2 nmoles of PO4/min/ mg protein. The trend, as in the case of alkaline phosphatase, is increasing in the first 72 hours post-hatching. Early feeding of both CHS and HBF promoted greater phytase activity in the first 72 hours of life (p≤0.05), with increases of up to 3.2 fold compared to the control (Figure 1).
Overall, in animals in the control group that fasted for 48 hrs. there was an effect of providing food after the fasting period, which caused an increase in the activity of integral enzymes in the intestinal brush border. In this case, in the duodenal segment, maltase, sucrase, alkaline phosphatase and phytase, at 72 hrs. after hatching. Similarly, in birds given some food or supplement (HBF or CHS) during the first 48 hrs. of life, a marked increasing tendency in the activity of these digestive enzymes was observed.
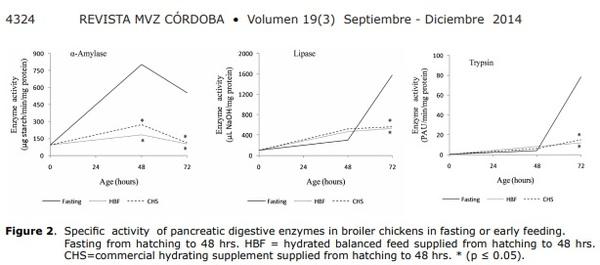
α-Amylase (EC 3.2.1.1). Immediately after hatching, the specific activity of the α-amylase enzyme, determined in pancreatic homogenate, was 94 ± 7 μg of degraded starch min/mg protein. The trend was to present a greater value at 48 hrs. and to diminish at 72 hrs.
At 48 hrs. old, chickens receiving all treatments showed significantly lower activity of this enzyme (p≤0.05) in comparison with control animals, with greater activity in T0 very close to 300% with respect to immediately lesser treatment [T2] (Figure 2).
Trypsin (EC 3.4.21.4). Immediately after hatching, the specific activity of the trypsin enzyme, determined in pancreatic homogenate, was 0.3 ± 0.06 PAU/min/mg protein and showed an increasing trend in the control animals, in the first 72 hrs. of life.
Supplying CHS or HBF did not increase the activity of this enzyme (Figure 2). In the first 48 hrs. of life, activity was similar in both animals that fasted and those that received any food or supplements. After 48 hours of age, a marked increase in trypsin activity was observed only in animals that had been fasting (T0) and a significant reduction (p≤0.05) in activity is observed in groups that received HBF and CHS (T1 and T2).
Lipase (EC 3.1.1.3). Immediately after hatching, the specific activity of the lipase enzyme, determined in pancreatic homogenate, was 106 ± 17 μl of NaOH/mg protein. The general trend was increasing in the first 72 hrs.
On the second day of life, all treatments showed a tendency to increase this enzyme’s activity in relation to birth, even in birds that were under total fasting conditions (T0). At 72 hrs. accelerated activity of this enzyme was observed in the group of animals that fasted for 48 hrs. (Figure 2).
DISCUSSION
The supply of both HBF and CHS provided from birth until 48 hrs. post-hatching caused an increase in the activity of the maltase enzyme. This effect could have a favorable impact, in terms of better digestive activity, increased glucose uptake from the diet, and at the same time would improve the energy metabolism of the bird. The specific activity of maltase showed a marked increase with age, in agreement with trends reported by other authors (13).
Sucrase activity was almost 32 times lower than maltase activity after hatching. In older chickens, it has been reported that sucrase activity is usually about 13 to 16% lower than that of maltase (13). Providing HBF and CHS in the first 48 hrs. of life increased sucrase activity at 72 hrs. (p≤0.05). Like maltase, specific sucrase activity increased with the age of the animals over the time span of the study period and increased after food restriction; similar to findings reported by Pinheiro et al (18). It has been reported that digestive enzyme activity may be affected by characteristics of diet and availability of substrate (19), among other factors. The activity of the enzymes maltase, sucrase, alkaline phosphatase and phytase, all integral catalytic proteins of the intestinal brush border, determined in the duodenal segment, increased in the first 72 hrs. of life in groups that received some type of food or supplement immediately after hatching.
In assessing the effect of feed restriction between 7 and 14 days of life in broiler chickens, an increase was reported in maltase and sucrase activity immediately after the restriction period (18). This could be applied to partially explain the effect of the food supply at 48 hrs. on birds in the control group, by increasing the activity of duodenal enzymes.
As for pancreatic enzymes, some authors have reported that specific activity of digestive enzymes increases with age, peaking in the pancreas on the 8th day for amylase and lipase, on the 11th day for trypsin and on the 17th day for amylase (9). During the first 48 hrs. of life, trypsin activity was similar in all groups, including animals that were fasting, indicating that activity is not very dependent on the presence of substrate, in accordance with findings reported by other authors (20) who observed that trypsin activity was not affected after evaluating two different energy levels in the diet of broiler chickens during the first week of life. The results obtained in the present study may be related to previous studies which have shown that when you start feeding chickens 36 hrs. after hatching there is a significant decrease in body weight in the first three days of life (11) as well as growth retardation and reduced meat production (6), which may result from decreased digestive enzyme activity in the intestinal brush border.
In conclusion, providing HBF or CHS to chickens during the first 48 hours of life increased specific activity of duodenal enzymes along the intestinal brush border: maltase, sucrase and phytase, and decreased pancreatic enzyme activity for α-amylase, trypsin and lipase in the first 3 days of life.
This article was originally published in Revista MVZ Córdoba 19(3):4316-4327, 2014. ISSN: 0122-0268.







.jpg&w=3840&q=75)