Does selection for growth rate in broilers affect their resistance and tolerance to Eimeria maxima?
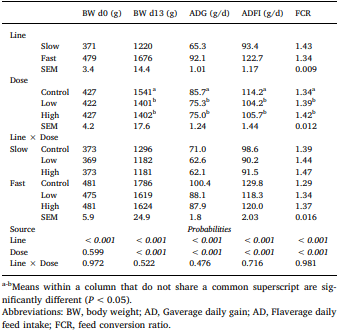
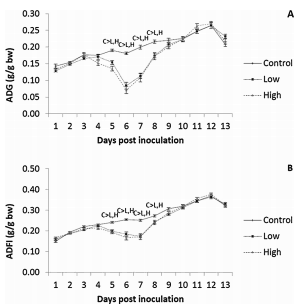
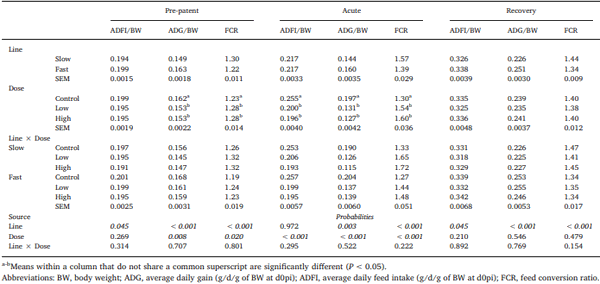
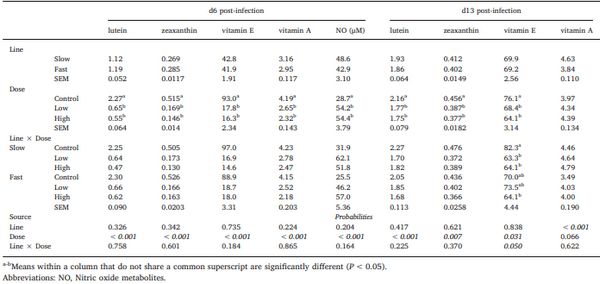
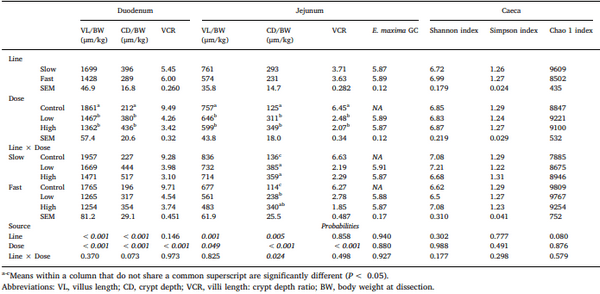
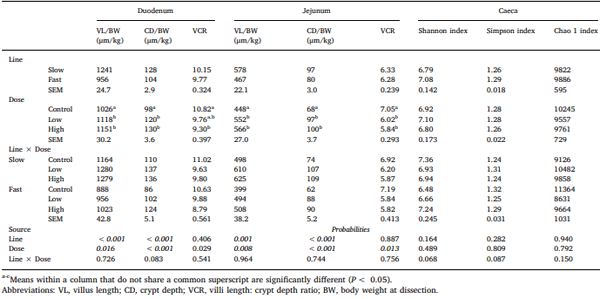
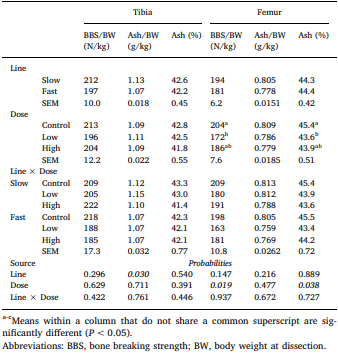
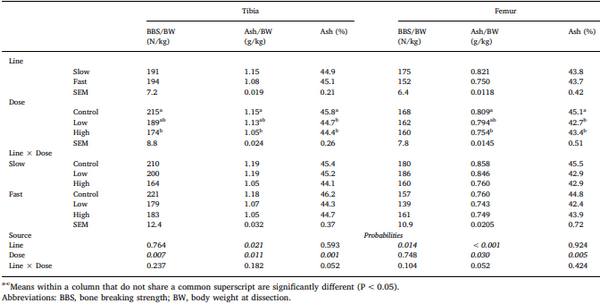
Chickens exhibit varied responses to infection with Eimeria parasites. We hypothesise that broilers selected for increased growth rate will show lower resistance and tolerance to a coccidian challenge. 288 chickens of fast (F) or slow (S) growing lines were inoculated with 0 (control), 2500 (low-dose), or 7000 (high-dose) sporulated E. maxima oocysts at 13 days of age in two consecutive rounds. Gain and Intake were measured daily and their values relative to BW at the point of infection were calculated over the pre-patent (days 1–4 post-infection), acute (d5–8pi), and recovery (d9–12pi) phases of infection to assess the impact of infection. Levels of plasma carotenoids, vitamins E and A, long bone mineralisation, caecal microbiota diversity indices, and histological measurements were assessed at the acute (d6pi) and recovery stage (d13pi). In addition, we measured the levels of nitric oxide metabolites and the number of parasite genome copies in the jejunum at d6pi. In absolute terms F birds grew 1.42 times faster than S birds when not infected. Infection significantly reduced relative daily gain and intake (P<0.001), with the effects being most pronounced during the acute phase (P< 0.001). Levels of all metabolites were significantly decreased, apart from NO which increased (P<0.001) in response to infection on d6pi, and were accompanied by changes in histomorphometric features and the presence of E.maxima genome copies in infected birds, which persisted to d13pi. Furthermore, infection reduced tibia and femur mineralisation, which also persisted to d13pi. Reductions in measured variables were mostly independent of dose size, as was the level of parasite replication. The impact of infection was similar for Sand F-line birds for all measured parameters, and there were no significant interactions between line x dose size on any of these parameters. In conclusion, our results suggest that line differences in productive performance do not influence host responses to coccidiosis when offered nutrient adequate diets.
Keywords: Coccidiosis Eimeria maxima Broiler Genetic selection Resistance Tolerance Growth rate Bone mineralisation.
Adams, C., Vahl, H.A., Veldman, A., 1996. Interaction between nutrition and Eimeria acervulina infection in broiler chickens: development of an experimental infection model. Br. J. Nutr. 75, 867–873.
Allen, P.C., 1992. Effect of coccidiosis on the distribution of dietary lutein in the chick. Poult. Sci. 71, 1457–1463.
Allen, P.C., 1997a. Nitric oxide production during Eimeria tenella infections in chickens. Poult. Sci. 76, 810–813.
Allen, P.C., 1997b. Production of free radical species during Eimeria maxima infections in chickens. Poult. Sci. 76, 814–821.
Allen, P.C., Fetterer, R.H., 2002a. Effects of dietary vitamin E on chickens infected with Eimeria maxima: observations over time of primary infection. Avian Dis. 46, 839–846.
Allen, P.C., Fetterer, R.H., 2002b. Interaction of dietary vitamin E with Eimeria maxima infections in chickens. Poult. Sci. 81, 41–48.
Allen, P.C., Lillehoj, H.S., 1998. Genetic influence on nitric oxide production during Eimeria tenella infections in chickens. Avian Dis. 42, 397–403.
Allen, P.C., Danforth, H.D., Vinyard, B.L., 2004. Development of a protective index to rank effectiveness of multiple treatments within an experiment: application to a cross-protection study of several strains of Eimeria maxima and a live vaccine. Avian Dis. 48, 370–375.
Allen, P.C., Jenkins, M.C., Miska, K.B., 2005. Cross protection studies with Eimeria maxima strains. Parasitol. Res. 97, 179–185.
Applegate, T.J., Lilburn, M.S., 2002. Growth of the femur and tibia of a commercial broiler line. Poult. Sci. 81, 1289–1294.
Aviagen, 2014a. Ross 308: Nutrition Specifications. Aviagen Ltd., Scotland, UK.
Aviagen, 2014b. Ross Broiler Management Handbook. Aviagen Ltd., Scotland, UK.
Blake, D.P., Hesketh, P., Archer, A., Shirley, M.W., Smith, A.L., 2006. Eimeria maxima: the influence of host genotype on parasite reproduction as revealed by quantitative realtime PCR. Int. J. Parasitol. 36, 97–105.
Blake, D.P., Qin, Z., Cai, J., Smith, A.L., 2008. Development and validation of real-time polymerase chain reaction assays specific to four species of Eimeria. Avian Pathol. 37, 89–94.
Brickett, K.E., Dahiya, J.P., Classen, H.L., Annett, C.B., Gomis, S., 2007. The impact of nutrient density, feed form, and photoperiod on the walking ability and skeletal quality of broiler chickens. Poult. Sci. 86, 2117–2125.
Bumstead, J.M., Bumstead, N., Rothwell, L., Tomley, F.M., 1995. Comparison of immune responses in inbred lines of chickens to Eimeria maxima and Eimeria tenella. Parasitology 111, 143–151.
Cheema, M.A., Qureshi, M.A., Havenstein, G.B., 2003. A comparison of the immune response of a 2001 commercial broiler with a 1957 randombred broiler strain when fed representative 1957 and 2001 broiler diets. Poult. Sci. 82, 1519–1529.
Coltherd, J.C., Bunger, L., Kyriazakis, I., Houdijk, J.G., 2009. Genetic growth potential interacts with nutrition on the ability of mice to cope with Heligmosomoides bakeri infection. Parasitology 136, 1043–1055.
Coltherd, J.C., Babayan, S.A., Bünger, L., Kyriazakis, I., Allen, J.E., Houdijk, J.G.M., 2011. Interactive effects of protein nutrition, genetic growth potential and Heligmosomoides bakeri infection pressure on resilience and resistance in mice. Parasitology 138, 1305–1315.
Conway, D.P., Sasai, K., Gaafar, S.M., Smothers, C.D., 1993. Effects of different levels of oocyst inocula of Eimeria acervulina, E. tenella, and E. maxima on plasma constituents, packed cell volume, lesion scores, and performance in chickens. Avian Dis. 37, 118–123.
Coop, R.L., Kyriazakis, I., 1999. Nutrition-parasite interaction. Vet. Parasitol. 84, 187–204.
Cornelissen, J.B.W.J., Swinkels, W.J.C., Boersma, W.A., Rebel, J.M.J., 2009. Host response to simultaneous infections with Eimeria acervulina, maxima and tenella: a cumulation of single responses. Vet. Parasitol. 162, 58–66.
Dawkins, M.S., Layton, R., 2012. Breeding for better welfare: genetic goals for broiler chickens and their parents. Anim. Welf. 21, 147–155.
Doeschl-Wilson, A.B., Kyriazakis, I., 2012. Should we aim for genetic improvement in host resistance or tolerance to infectious pathogens? Front. Genet. 3, 272.
Doeschl-Wilson, A.B., Brindle, W., Emmans, G., Kyriazakis, I., 2009. Unravelling the relationship between animal growth and immune response during micro-parasitic infections. PLoS ONE 4, e7508.
Fetterer, R.H., Miska, K.B., Mitchell, A.D., Jenkins, M.C., 2013. The use of dual-energy Xray absorptiometry to assess the impact of Eimeria infections in broiler chicks. Avian Dis. 57, 199–204.
Furlong, R.F., 2005. Insights into vertebrate evolution from the chicken genome sequence. Genome Biol. 6, 207.
Gabriel, I., Mallet, S., Leconte, M., Fort, G., Naciri, M., 2006. Effects of whole wheat feeding on the development of coccidial infection in broiler chickens until market age. Anim. Feed Sci. Technol. 129, 279–303.
Giraldo, C.A., Brown, D.R., Watkins, K.L., Southern, L.L., 1987. Responses to excess dietary magnesium as affected by experimental Eimeria acervulina infection or by dietary ammonium chloride ingestion in the chick. J. Nutr. 117, 1053–1059.
González-Cerón, F., Rekaya, R., Aggrey, S.E., 2015. Genetic analysis of bone quality traits and growth in a random mating broiler population. Poult. Sci. 94, 883–889.
Harrison, E.H., 2005. Mechanisms of digestion and absorption of dietary vitamin A. Annu. Rev. Nutr. 25, 87–103.
Havenstein, G.B., Ferket, P.R., Qureshi, M.A., 2003. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 82, 1500–1508.
Hein, H., 1968. The pathogenic effects of Eimeria acervulina in young chicks 1. Exp. Parasitol. 22, 1–11.
Hernández-Velasco, X., Chapman, H.D., Owens, C.M., Kuttappan, V.A., Fuente-Martínez, B., Menconi, A., Latorre, J.D., Kallapura, G., Bielke, L.R., Rathinam, T., Hargis, B.M., Tellez, G., 2014. Absorption and deposition of xanthophylls in broilers challenged with three dosages of Eimeria acervulina oocysts. Brit. Poult. Sci. 55, 167–173.
Hocking, P.M., 2014. Unexpected consequences of genetic selection in broilers and turkeys: problems and solutions. Brit. Poult. Sci. 55, 1–12.
Hong, Y.H., Lillehoj, H.S., Lillehoj, E.P., Lee, S.H., 2006. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet. Immunol. Immunopathol. 114, 259–272.
Humphries, J.M., Khachik, F., 2003. Distribution of lutein, zeaxanthin, and related geometrical isomers in fruit, vegetables, wheat, and pasta products. J. Agric. Food Chem. 51, 1322–1327.
Idris, A.B., Bounous, D.I., Goodwin, M.A., Brown, J., Krushinskie, E.A., 1997. Quantitative pathology of small intestinal coccidiosis caused by Eimeria maxima in young broilers. Avian Pathol. 26, 731–747.
Julian, R.J., 1998. Rapid growth problems: ascites and skeletal deformities in broilers. Poult. Sci. 77, 1773–1780.
Kapell, D.N., Hill, W.G., Neeteson, A.M., McAdam, J., Koerhuis, A.N., Avendano, S., 2012. Genetic parameters of foot-pad dermatitis and body weight in purebred broiler lines in 2 contrasting environments. Poult. Sci. 91, 565–574.
Kipper, M., Andretta, I., Lehnen, C.R., Lovatto, P.A., Monteiro, S.G., 2013. Meta-analysis of the performance variation in broilers experimentally challenged by Eimeria spp. Vet. Parasitol. 196, 77–84.
Klasing, K., 2004. The costs of immunity. Acta Zool. Sin. 50, 961–969.
Kyriazakis, I., 2010. Is anorexia during infection in animals affected by food composition? Anim. Feed Sci. Technol. 156, 1–9.
Kyriazakis, I., 2014. Pathogen-induced anorexia: a herbivore strategy or an unavoidable consequence of infection? Anim. Prod. Sci. 54, 1190–1197.
Kyriazakis, I., Houdijk, J.G.M., 2007. Food intake and performance of pigs during health disease and recovery. In: J Wiseman, J., Varley, M.A., McOrist, S., Kemp, B. (Eds.), Paradigms in Pig Science. Nottingham University Press, Nottingham, pp. 493–513.
Lillehoj, H.S., Li, G.X., 2004. Nitric oxide production by macrophages stimulated with coccidia sporozoites, lipopolysaccharide, or interferon-gamma, and its dynamic changes in SC and TK strains of chickens infected with Eimeria tenella. Avian Dis. 48, 244–253.
Lillehoj, H.S., Ruff, M.D., 1987. Comparison of disease susceptibility and subclass-specific antibody response in Sc and Fp chickens experimentally inoculated with Eimeria Tenella, Eimeria Acervulina, or Eimeria Maxima. Avian Dis. 31, 112–119.
Lillehoj, H.S., Trout, J.M., 1996. Avian gut associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 9, 349–360.
Liu, Z., Lee, H.-J., Garofalo, F., Jenkins, D.J.A., El-Sohemy, A., 2011. Simultaneous measurement of three tocopherols, all-trans-retinol, and eight carotenoids in human plasma by isocratic liquid chromatography. J. Chromatogr. Sci. 49, 221–227.
Long, P.L., Millard, B.J., Joyner, L.P., Norton, C.C., 1976. A guide to laboratory techniques used in the study and diagnosis of avian coccidiosis. Folia Vet. Lat. 6, 201–217.
Lough, G., Kyriazakis, I., Bergmann, S., Lengeling, A., Doeschl-Wilson, A.B., 2015. Health trajectories reveal the dynamic contributions of host genetic resistance and tolerance to infection outcome. Proc. R. Soc. Lond. B. Biol. Sci. 282.
Macdonald, S.E., Nolan, M.J., Harman, K., Boulton, K., Hume, D.A., Tomley, F.M., Stabler, R.A., Blake, D.P., 2017. Effects of Eimeria tenella infection on chicken caecal microbiome diversity, exploring variation associated with severity of pathology. PLoS One 12, e0184890.
Martynova-Van Kley, M.A., Oviedo-Rondon, E.O., Dowd, S.E., Hume, M., Nalian, A., 2012. Effect of Eimeria infection on cecal microbiome of broilers fed essential oils. Int. J. Poult. Sci. 11, 747–755.
McDevitt, R.M., McEntee†, G.M., Rance†, K.A., 2006. Bone breaking strength and apparent metabolisability of calcium and phosphorus in selected and unselected broiler chicken genotypes. Br. Poult. Sci. 47, 613–621.
Nolan, M.J., Tomley, F.M., Kaiser, P., Blake, D.P., 2015. Quantitative real-time PCR (qPCR) for eimeria tenella replication—implications for experimental refinement and animal welfare. Parasitol. Int. 64, 464–470.
Persia, M.E., Young, E.L., Utterback, P.L., Parsons, C.M., 2006. Effects of dietary ingredients and eimeria acervulina infection on chick performance, apparent metabolizable energy, and amino acid digestibility. Poult. Sci. 85, 48–55.
Pinard-Van Der Laan, M.H., Monvoisin, J.L., Pery, P., Hamet, N., Thomas, M., 1998. Comparison of outbred lines of chickens for resistance to experimental infection with coccidiosis (Eimeria tenella). Poult. Sci. 77, 185–191.
Polansky, O., Sekelova, Z., Faldynova, M., Sebkova, A., Sisak, F., Rychlik, I., 2016. Characterisation of the most important metabolic pathways and biological processes expressed in chicken caecal microbiota. Appl. Environ. Microbiol. 82, 1569–1576.
Preston-Mafham, R.A., Sykes, A.H., 1970. Changes in body weight and intestinal absorption during infections with Eimeria acervulina in the chicken. Parasitology 61, 417–424.
Qadir, O.K., Teh, J., Siervo, M., Seal, C.J., Brandt, K., et al., 2013. Method using gas chromatography mass spectrometry (GC-MS) for analysis of nitrate and nitrite in vegetables. In: D’Haene, K., Vandecasteele, B., De Vis, R., Crappé, S., Callens, D., Mechant, E. (Eds.), NUTRIHORT: Nutrient Management, Innovative Techniques and Nutrient Legislation in Intensive Horticulture for an Improved Water Quality. Institute for Agricultural and Fisheries Research, Ghent, Belgium.
Rauw, W.M., 2012. Immune response from a resource allocation perspective. Front. Genet. 3, 267.
Rauw, W.M., Kanis, E., Noordhuizen-Stassen, E.N., Grommers, F.J., 1998. Undesirable side effects of selection for high production efficiency in farm animals: a review. Livest. Prod. Sci. 56, 15–33.
Reid, A.J., Blake, D.P., Ansari, H.R., Billington, K., Browne, H.P., Bryant, J., Dunn, M., Hung, S.S., Kawahara, F., Miranda-Saavedra, D., Malas, T.B., Mourier, T., Naghra, H., Nair, M., Otto, T.D., Rawlings, N.D., Rivailler, P., Sanchez-Flores, A., Sanders, M., Subramaniam, C., Tay, Y.L., Woo, Y., Wu, X., Barrell, B., Dear, P.H., Doerig, C., Gruber, A., Ivens, A.C., Parkinson, J., Rajandream, M.A., Shirley, M.W., Wan, K.L., Berriman, M., Tomley, F.M., Pain, A., 2014. Genomic analysis of the causative agents of coccidiosis in domestic chickens. Genome Res. 24, 1676–1685.
Sandberg, F.B., Emmans, G.C., Kyriazakis, I., 2007. The effects of pathogen challenges on the performance of naive and immune animals: the problem of prediction. Animal 1, 67–86.
Schneider, C.A., Rasband, W.S., Eliceiri, K.W., 2012. NIH image to imageJ: 25 years of image analysis. Nat. Methods 9, 671–675.
Schokker, D., Veninga, G., Vastenhouw, S.A., Bossers, A., de Bree, F.M., Kaal-Lansbergen, L.M.T.E., Rebel, J.M.J., Smits, M.A., 2015. Early life microbial colonization of the gut and intestinal development differ between genetically divergent broiler lines. BMC Genom. 16, 418.
Sharma, V.D., Fernando, M.A., 1975. Effect of Eimeria acervulina infection on nutrient retention with special reference to fat malabsorption in chickens. Can. J. Comp. Med. 39, 146–154.
Shaw, A.L., van Ginkel, F.W., Macklin, K.S., Blake, J.P., 2011. Effects of phytase supplementation in broiler diets on a natural Eimeria challenge in naive and vaccinated birds. Poult. Sci. 90, 781–790.
Shim, M.Y., Karnuah, A.B., Anthony, N.B., Pesti, G.M., Aggrey, S.E., 2012. The effects of broiler chicken growth rate on valgus, varus, and tibial dyschondroplasia. Poult. Sci. 91, 62–65.
Siegel, P.B., 2014. Evolution of the modern broiler and feed efficiency. Annu. Rev. Anim. Biosci. 2, 375–385.
Singh, S.P., Donovan, G.A., 1973. A relationship between coccidiosis and dietary vitamin a levels in chickens. Poult. Sci. 52, 1295–1301.
Su, S., Miska, K.B., Fetterer, R.H., Jenkins, M.C., Wong, E.A., 2014. Expression of digestive enzymes and nutrient transporters in Eimeria acervulina challenged layers and broilers. Poult. Sci. 93, 1217–1226.
Swaggerty, C.L., Pevzner, I.Y., He, H., Genovese, K.J., Nisbet, D.J., Kaiser, P., Kogut, M.H., 2009. Selection of broilers with improved innate immune responsiveness to reduce on-farm infection by foodborne pathogens. Foodborne Pathog. Dis. 6, 777–783
Swaggerty, C.L., Pevzner, I.Y., Kogut, M.H., 2015. Selection for pro-inflammatory mediators produces chickens more resistant to Eimeria tenella. Poult. Sci. 94, 37–42.
Swinkels, W.J.C., Post, J., Cornelissen, J.B., Engel, B., Boersma, W.J.A., Rebel, J.M.J., 2007. Immune responses to an Eimeria acervulina infection in different broilers lines. Vet. Immunol. Immunopathol. 117, 26–34.
Takhar, B.S., Farrell, D.J., 1979. Energy and nitrogen metabolism of chickens subjected to infection and reinfection with Eimeria acervulina. Brit. Poult. Sci. 20, 213–224.
Tallentire, C.W., Leinonen, I., Kyriazakis, I., 2016. Breeding for efficiency in the broiler chicken: a review. Agron. Sustain. Dev. 36, 66.
Tixier-Boichard, M., Leenstra, F., Flock, D.K., Hocking, P.M., Weigend, S., 2012. A century of poultry genetics. World’s Poult. Sci. J. 68, 307–321.
Tsikas, D., 2000. Simultaneous derivatization and quantification of the nitric oxide metabolites nitrite and nitrate in biological fluids by gas chromatography/mass spectrometry. Phy/mass spectrometry. Anal. Chem. 72, 4064–4072. Turk, D.E., 1973. Calcium absorption during coccidial infections in chicks. Poult. Sci. 52, 854–857.
van der Aar, P., Doppenberg, J., Kwakernaak, C., Burton, E., Gatcliffe, J., O’Neill, H., Scholey, D., 2016. In: Burton, E., Gatcliffe, J., O’Neill, H.M., Scholey, D. (Eds.), Sustainable Poultry Production in Europe. CABI, Oxon, pp. 103–111.
van der Most, P.J., de Jong, B., Parmentier, H.K., Verhulst, S., 2011. Trade-off between growth and immune function: a meta-analysis of selection experiments. Funct. Ecol. 25, 74–80.
Varmuzova, K., Kubasova, T., Davidova-Gerzova, L., Sisak, F., Havlickova, H., Sebkova, A., Faldynova, M., Rychlik, I., 2016. Composition of gut microbiota influences resistance of newly hatched chickens to Salmonella enteritidis infection. Front. Microbiol. 7, 957.
Ward, T.L., Watkins, K.L., Southern, L.L., 1990. Interactive effects of sodium zeolite a (ethacal) and monensin in uninfected and Eimeria acervulina infected chicks. Poult. Sci. 69, 276–280.
Watkins, K.L., Vagnoni, D.B., Southern, L.L., 1989. Effect of dietary sodium zeolite A and excess calcium on growth and tibia calcium and phosphorus concentration in uninfected and Eimeria acervulina-infected chicks. Poult. Sci. 68, 1236–1240.
Watson, B.C., Matthews, J.O., Southern, L.L., Shelton, J.L., 2005. The interactive effects of Eimeria acervulina infection and phytase for broiler chicks. Poult. Sci. 84, 910–913.
Williams, R.B., 2001. Quantification of the crowding effect during infections with the seven Eimeria species of the domesticated fowl: its importance for experimental designs and the production of oocyst stocks. Int. J. Parasitol. 31, 1056–1069.
Williams, R.B., 2005. Intercurrent coccidiosis and necrotic enteritis of chickens: rational, integrated disease management by maintenance of gut integrity. Avian Pathol. 34, 159–180.
Williams, B., Solomon, S., Waddington, D., Thorp, B., Farquharson, C., 2000. Skeletal development in the meat-type chicken. Brit. Poult. Sci. 41, 141–149.
Williams, B., Waddington, D., Murray, D.H., Farquharson, C., 2004. Bone strength during growth: influence of growth rate on cortical porosity and mineralization. Calcif. Tissue Int. 74, 236–245.
Willis, G.M., Baker, D.H., 1981. Phosphorus utilization during Eimeria acervulina infection in the chick. Poult. Sci. 60, 1960–1962.
Yunis, R., Ben-David, A., Heller, E.D., Cahaner, A., 2000. Immunocompetence and viability under commercial conditions of broiler groups differing in growth rate and in antibody response to Escherichia coli vaccine. Poult. Sci. 79, 810–816.
Zaralis, K., Tolkamp, B.J., Houdijk, J.G.M., Wylie, A.R.G., Kyriazakis, I., 2008. Changes in food intake and circulating leptin due to gastrointestinal parasitism in lambs of two breeds1. J. Anim. Sci. 86, 1891–1903.
Zekarias, B., Ter Huurne, A.A.H.M., Landman, W.J.M., Rebel, J.M.J., Pol, J.M.A., Gruys, E., 2002. Immunological basis of differences in disease resistance in the chicken. Vet. Res. 33, 109–125.
Zhou, Z., Nie, K., Huang, Q., Li, K., Sun, Y., Zhou, R., Wang, Z., Hu, S., 2017. Changes of cecal microflora in chickens following Eimeria tenella challenge and regulating effect of coated sodium butyrate. Exp. Parasitol. 177, 73–81.
Zhu, J.J., Lillehoj, H.S., Allen, P.C., Yun, C.H., Pollock, D., Sadjadi, M., Emara, M.G., 2000. Analysis of disease resistance-associated parameters in broiler chickens challenged with Eimeria maxima. Poult. Sci. 79, 619–625. Zuidhof, M.J., Schneider, B.L., Carney, V.L., Korver, D.R., Robinson, F.E., 2014. Growth, efficiency, and yield of commercial broilers from 1957, 1978, and 20051. Poult. Sci. 93, 2970–2982.
P. Sakkas et al. Veterinary Parasitology 258 (2018) 88–98 9





L.S.,
In your introduction you mentioned:
Such an emphasis on productive traits may have compromised the ability of modern broilers to cope with metabolic and skeletal disorders (Dawkins and Layton, 2012; Julian, 1998) and infectious pathogens (Cheema et al., 2003; Yunis et al., 2000). This raises concerns amongst the general public and have led, for example, the Dutch Organisation of Retailers to take the strategic decision that they will only sell chicken meat from slow-growing animals.
The reasons, however, why Dutch retailers chose for slow growing birds are very diverse. It was not so much the general public but more the pressure of action groups with different agendas that forced the retailers to take actions. These actions were partly based on false accusations.
Different parameters of disease and welfare in regular broilers were and are very good in the Netherlands. The average mortality rates and the amount of antibiotics used are very low in both the regular and the slower growing birds.
To our surprise, however, we are seeing more severe outbreaks of Coccidiosis (mainly E. tenella) in slower growing birds than in the regular broilers.
I would be curious to hear about your ideas about this observation.
Regards.








.jpg&w=3840&q=75)