Glucagon-like peptide (GLP) is one of the major intestinal hormones and has two subtypes, GLP-1 and GLP-2. GLP-1 is a 36-amino acid peptide released from intestinal L cells in response to ingested nutrients [1]. In the gut, this peptide hormone performs a variety of physiological and biological roles. Some of these functions include enhancing glucose-dependent insulin release, inhibiting glucagon secretion, increasing pancreatic β cell growth [10, 27], decreasing food intake [1], decelerating gastric emptying [27], inhibiting intestinal motility [36], and stimulating intestinal growth [21].
GLP-2 is a 33-amino acid intestinotrophic peptide [35] and has a range of effects on the gastrointestinal (GI) tract. This hormone promotes intestinal development by increasing epithelial proliferation and decreasing apoptosis [13]. GLP-2 also suppresses gastric acid secretion, reduces gastric motility, and regulates food intake [3, 6, 38].
GLP-1 and GLP-2 are cleaved from their common precursor protein, proglucagon (PG), in intestinal L cells by unique posttranslational proteolytic cleavage and other enzymatic changes [4, 9]. L cells are widely found in the small intestine and colon of mammals [23]. In chickens, L cells are located throughout the entire jejunum and ileum [18, 19] and L cell apical surface is covered with microvilli [28]. These microvilli contain several receptors and channels that receive chemical signals from the gut lumen, making them open-type enteroendocrine cells (EECs) [15].
The synthesis of GLPs positively correlates with the density of intestinal L cells [5]. Consequently, an increase in L cell density may enhance the physiological and biological activities of GLPs in the GI tract. In fact, our previous studies in chickens demonstrated that dietary protein and the supplement of amino acids increased the density of GLP-1-immunoreactive cells in the chicken ileum [24, 29]. However, relatively little data exist on the impact of dietary carbohydrate (CHO) levels on the density of L cells in the chicken ileum.
We recently revealed that dietary CHO positively affected the proliferation of epithelial cells in the chicken ileum [34]. In the present study, we aimed to evaluate the effect of dietary CHO on the density and proliferation of L cells in the chicken ileum using immunohistochemistry and in situ hybridization.
MATERIALS AND METHODS
The animal experiment protocol was reviewed by the Committee for Animal Experiments and approved by the president of Shinshu University (Approval Number 300090). Chickens were kept in the facility of the experiments for avian species of the Faculty of Agriculture, Shinshu University.
Animals and feeding management
Six-week-old, male White Leghorn chickens (n=15) were used in this study. They were assigned into the control, low-CHO, and CHO-free groups based on the average body weight. Each group consisted of five birds. Initially, all chickens received the control diet in separated cages for 3 days to acclimate them to the experimental circumstances. Subsequently, each experimental diet was supplied to the corresponding group for 7 days. Experimental diets for low-CHO and CHO-free groups contained 12.5% and 0% CHO of that of the control group which contains 491.4 g CHO/kg diet, respectively. Metabolizable energy (ME) of the two experimental diets was sustained at the same level as the control diet (ME=2,850 kcal/kg) by adding corn oil and cellulose. Therefore, the energy content of the diet satisfied the ME requirement specified by the Japanese Feeding Standard for Poultry [26]. Chickens could access feed and water freely during the experimental period. The daily feed intake and body weight of each chicken were measured at the same time during the experimental period.
Sample collection
All chickens were sacrificed through decapitation under anesthesia with sodium pentobarbital injection on the last day of the experimental period. Tissue samples approximately 2 cm long were taken from the proximal and distal parts of the ileum of each bird. After washing in 0.75% NaCl solution, tissue samples were fixed in Bouin’s solution for 24 hr at 4°C and then embedded in paraffin wax, according to the standard method. Paraffin sections cut at 5 μm thickness from each tissue sample were used in the following procedures.
Immunohistochemistry
The streptavidin-biotin method [14] was used to identify both GLP-1- and GLP-2-immunoreactive cells [16]. For the detection of GLP-1 immunoreactivity, sections treated with normal goat serum were incubated with rabbit antiserum against synthetic GLP-1 (1–19) (Affiniti Research Products, Devon, UK, No. GA1176, diluted to 1:2,000) for 24 hr at room temperature. For the detection of GLP-2 immunoreactivity, sections were first treated with a 0.5% antigen retrieval agent (Immunosaver®, Nisshin EM, Tokyo, Japan) at 98°C for 45 min [25]. They were then incubated with rabbit antiserum against human (Arg34)-GLP-2 (1:2,000, H-028-14, Phenix Pharmaceuticals, Burlingame, CA, USA) as a primary antibody for 24 hr following the treatment with 10% normal goat serum.
After several washes with phosphate buffered saline (PBS), all sections were incubated with biotin-labeled goat antiserum against rabbit immunoglobulin G (IgG, AP132B, Millipore, Temecula, CA, USA, 1:300) and streptavidin-polyHRP20 (SP20C, Stereospecific Detection Technologies, Baesweiler, Germany, 1:300) as the secondary antibody and label for immunocomplex, respectively. Immunocomplex was visualized through incubation with a 3,3′-diaminobenzidine-hydrogen peroxide solution. Preparations were counterstained with Mayer’s hematoxylin and observed under a light microscope. All incubations were performed in a moisture chamber at room temperature.
Morphometry
The occurrence of GLP-1- and GLP-2-immunoreactive cells was assessed in the following manner [19]. First, immunoreactive cells for GLP-1 and GLP-2 antisera with a clearly visible nucleus were counted on a microphotograph taken at 5× magnification. Second, the mucosal area was measured and the frequency of immunoreactive cells (cells/mm2 ± SD) was calculated. A computerized image analysis system (KS-400, ZEISS, Göttingen, Germany) was used for this quantification. Twenty areas were randomly selected from each ileal part of each chicken. In total, 100 areas of each ileal part were assessed from the five chickens in each group.
In situ hybridization and double immunofluorescence method
Several sets of mirror sections cut at 5 μm thickness were utilized in the following procedures. An oligonucleotide probe labeled with digoxigenin (DIG) was commercially synthesized (BEX, Tokyo, Japan) according to the chicken PG sequence shown by Richards and McMurtry [32]. Sequences of PG antisense and sense probes were 5′-GCTGTAGTCACTGGTGAATGTGCCTTGTGAATGACGCTTTA-3′ and 5′-TAAAGCGTCATTCACAAGGCACATTCACCAGTGACTACAGC-3′, respectively. The signal of PG mRNA was detected using a commercial in situ hybridization kit (IsHyb In Situ Hybridization Kit; Biochain Institute, Newark, CA, USA) on a pair of mirror section [37]. The sections were then treated with anti-DIG serum diluted with alkaline phosphatase solution (1:500), and the conjugated probe was visualized using the mixture solution of nitro blue tetrazolium and 5-bromo-4-chloro-3-indolyl-phosphate. The sense probe was utilized as a negative control for PG mRNA.
Double immunofluorescence for GLP-1 and GLP-2 was conducted on complementary sections of mirror sections stained for in situ hybridization mentioned above. Sections treated with antigen retrieval agent (Immunosaver®) were incubated with 10% normal goat serum. They were treated with a mouse monoclonal antibody against synthetic human GLP-1 (aa 7-36 amide) (A6104.1, Immunodiagnostik, Bensheim, Germany, 1:2,000) for 24 hr. Samples were subsequently treated with a rabbit antiserum against human (Arg34)-GLP-2 serum (1:500, H-028-14, Phenix Pharmaceuticals) for 24 hr. After washing with PBS, sections were incubated with a cocktail of DyLight 488-conjugated goat anti-mouse IgG (611-741-127; Rockland Immunochemicals, Gilbertsville, PA, USA, diluted to 1:300) and DyLight 549-conjugated goat anti-rabbit IgG (611-700-127; Rockland Immunochemicals, diluted to 1:300) for 3 hr at room temperature. Finally, sections were covered with coverslips in an aqueous mounting medium and photographed using a fluorescence microscope (AxioImagerA1, Zeiss).
Immunohistochemistry for Ki-67 to determine L cells proliferation
Double immunofluorescence using an antiserum against Ki-67 was conducted to evaluate the influence of dietary CHO on L cell proliferation. Ki-67 is expressed as a nonhistone nucleoprotein during all stages of the cell cycle [22], indicating that it is valuable as a marker of cell proliferation. Paraffin sections were treated with 10% HistoVT one (Nacalai tesque, Kyoto, Japan) as an antigen retrieval agent for 20 min at 90°C. After several PBS washes, sections were treated with 2.5% normal donkey serum, and incubated with mouse monoclonal antibody against Ki-67 (MAB4190, Sigma-Aldrich, St. Louis, MO, USA, 1:500) for 24 hr. Following PBS washing, sections were treated with 10% normal goat serum and incubated with rabbit antiserum against synthetic GLP-1 (1–19) conjugated to bovine serum. Immunocomplexes were revealed using a cocktail of DyLight 549-conjugated donkey anti-mouse IgG serum and FITC-conjugated goat anti-rabbit IgG serum for 3 hr. All incubations took place in a moisture chamber at room temperature.
Statistical analysis
Frequencies of GLP-1- and GLP-2-immunoreactive cells were given as mean ± standard error of the mean. One-way analysis of variance was used. Additionally, Tukey’s test was used to assess any significant differences in the mean among groups. The significance level for the data was set at P< 0.05. All statistical analyses were conducted using statistical analysis software (SAS, Inst. Inc., Cary, NC, USA).
RESULTS
Effects of dietary CHO on GLP-1- and GLP-2-immunoreactive cells
Many endocrine cells immunoreactive for GLP-1 antiserum were found throughout the ileum. GLP-1-immunoreactive cells were scattered in the epithelium of middle to bottom parts of villi and crypts (Fig. 1Aa, 1Ad, arrows) in the proximal and distal ileum. There were no discernible variations in the distribution of GLP-1-immunoreactive cells among the control and the two experimental groups. In the control group, GLP-1-immunoreactive cells had a spindle-like or pyramidal shape with a long cytoplasmic process reaching the intestinal lumen in the villus epithelium (Fig. 1Ab, arrows). These cells demonstrated a comma-like shape in crypts (Fig. 1Ac, arrow). GLP-1-immunoreactive cells in the two experimental groups were found in an oval or round shape in both the villus epithelium (Fig. 1Ae, arrows) and crypts (Fig. 1Af). Figure 1B and 1C show the frequencies of GLP-1-immunoreactive cells in the proximal and distal ileum, respectively. The frequency of GLP-1-immunoreactive cells was significantly reduced with the decrease of dietary CHO level in both ileal parts, with the lowest measured in the CHO-free group.
Many GLP-2-immunoreactive cells were also observed in the whole ileum. GLP-2-immunoreactive cells were detected in the epithelium of middle to bottom parts of the villi (Fig. 2Aa, arrows) and crypts (Fig. 2Ad, arrows). No apparent differences were observed in the distribution pattern of GLP-2-immunoreactive cells between the control and the experimental groups. Most of the cells in the control group exhibited a spindle-like shape with a long cytoplasmic process reaching the intestinal lumen in the villus epithelium (Fig. 2Ab, arrow) and a comma-like shape in crypts (Fig. 2Ac, arrows). In the two experimental groups, especially in the CHO-free group, GLP-2-immunoreactive cells in the villus epithelium (Fig. 2Ae, arrows) and crypts (Fig. 2Af, arrows) were more oval or round than those in the control group. Figure 2B and 2C show the frequencies of GLP-2-immunoreactive cells in the proximal and distal ileum, respectively. There were significant differences in the frequency of GLP-2-immunoreactive cells between the control and the experimental groups in both the proximal and distal ileum. GLP-2-immunoreactive cells were significantly reduced with the decrease of dietary CHO level in both ileal parts, and were lowest in the CHO-free group.
Effects of dietary CHO on PG mRNA signals in the ileum
Cells expressing PG mRNA signal were identified in the proximal and distal ileum in the control and both experimental groups (Fig. 3, top panels). These were mainly located in the epithelium of the bottom part of villi and crypts. There were no distinguishable differences in the distribution pattern of cells expressing PG mRNA signal between the control and the experimental groups for either part of the ileum. Sense probe control did not produce any discernible PG mRNA signal (Fig. 3, bottom panels).
However, apparent differences were observed in the relative frequency of cells expressing PG mRNA signal among the three groups (Table 1). The CHO-free group showed a lower frequency of cells expressing PG mRNA signal in the proximal ileum compared with that in the other two groups. There were no discernible differences between the control and the low-CHO groups. The relative frequency of cells expressing PG mRNA signal was reduced with the decrease of dietary CHO level in both ileal parts, with the lowest in the CHO-free group.
Most cells expressing PG mRNA signal exhibited immunoreactivity for both GLP-1 and GLP-2 in the proximal and distal ileum (Fig. 4a–c, arrows). However, there was no clear difference in the ratio of cells expressing PG mRNA signal and GLP immunoreactivity among the three groups. Cells expressing only PG mRNA signal were rarely observed in the control and two experimental groups (Fig. 4d–f, arrows).
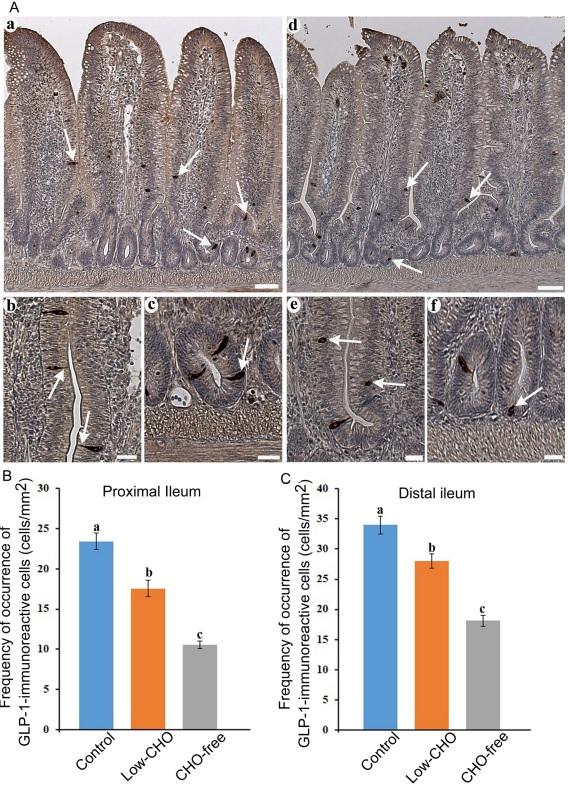
Fig. 1. Glucagon-like peptide (GLP)-1-immunoreactive cells in the chicken ileum. A: Photomicrographs showing the distribution of GLP-1-immunoreactive cells (arrows) in the distal ileum from the control (a) and carbohydrate (CHO)-free (d) groups. Bars: 50 μm. High magnification views of GLP-1-immunoreactive cells in the villus epithelium (b, e) and crypts (c, f). A long cytoplasmic process of GLP-1-immunoreactive cells reaches the intestinal lumen. In the CHO-free group, GLP-1-immunoreactive cells in a round or oval shape are frequently observed. Bars: 20 μm. B: Frequency of occurrence of GLP-1-immunoreactive cells in the proximal ileum from the control, low-CHO, and CHO-free groups. There is a significant difference between different alphabets. P< 0.05. Error bars: Standard error. a> b> c. C: Frequency of occurrence of GLP1-immunoreactive cells in the distal ileum from the control, low-CHO, and CHO-free groups. There is a significant difference between different alphabets. P< 0.05. Error bars: Standard error. a> b> c.
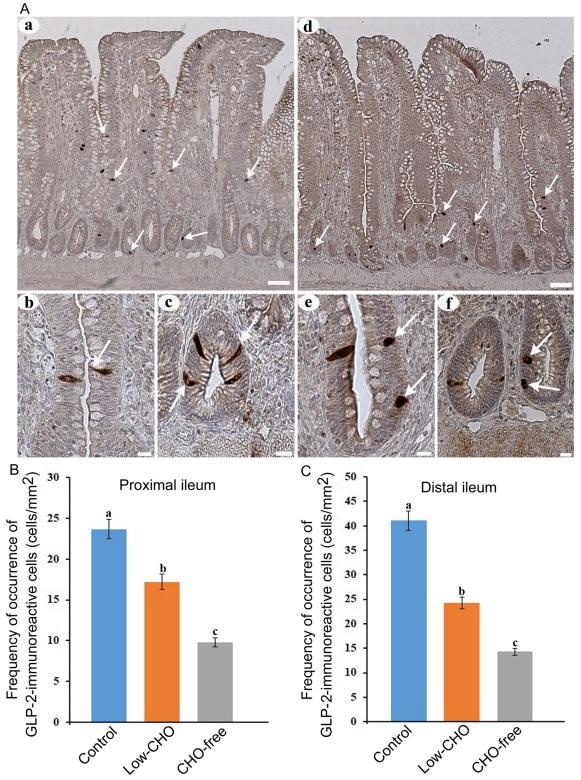
Fig. 2. Glucagon-like peptide (GLP)-2-immunoreactive cells in the chicken ileum. A: Photomicrographs showing the distribution of GLP-2-immunoreactive cells (arrows) in the distal ileum from the control (a) and carbohydrate (CHO)-free (e) groups. Bars: 50 μm. High magnification views of GLP-2-immunoreactive cells in villus epithelium (b, e) and crypts (c, f). A long cytoplasmic process of GLP-2-immunoreactive cells reaches the intestinal lumen (arrow in b). In the CHO-free group, GLP-2-immunoreactive cells in a round or oval shape are frequently observed (arrows in f). Bars: 20 μm. B: Frequency of occurrence of GLP-2-immunoreactive cells in the proximal ileum from the control, low-CHO, and CHO-free groups. There is a significant difference between different alphabets. P< 0.05. Error bars: Standard error. a> b> c. C: Frequency of occurrence of GLP-2-immunoreactive cells in the distal ileum from the control, low-CHO, and CHO-free groups. There is a significant difference between different alphabets. P< 0.05. Error bars: Standard error. a> b> c.
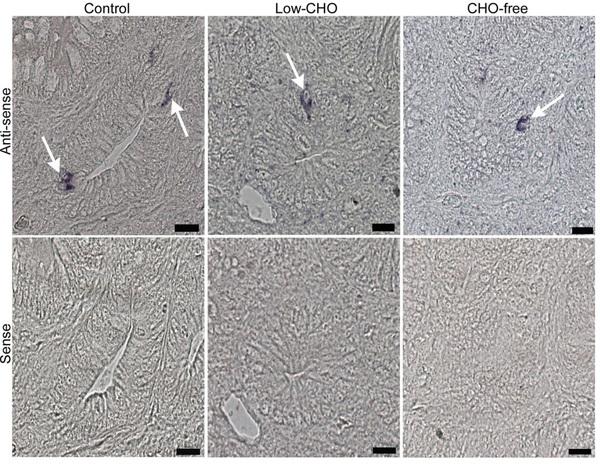
Fig. 3. Photomicrographs indicating cells expressing proglucagon (PG) mRNA signal in the chicken distal ileum from the control, low-carbohydrate (CHO), and CHO-free groups. Antisense probe shows cells expressing PG mRNA signal (arrows in top panels) mainly in crypts of all groups. However, the sense probe shows no cells showing PG mRNA signal (bottom panels). Bars: 20 μm.
Effects of dietary CHO on ileal L cell proliferation
Double immunofluorescence revealed the coexpression of Ki-67 and GLP-1 immunoreactivity in the same cell (Fig. 5A, arrows in top panels). The ratio of GLP-1-immunoreactive cells also showing Ki-67 immunoreactivity was similar in the proximal ileum among the three groups (Fig. 5B). However, this was reduced with the decrease of dietary CHO level in the distal ileum. A significant difference was revealed between the control and the CHO-free groups (Fig. 5C). GLP-1-immunoreactive cells without Ki-67-immunoreactivity were also observed in all groups (Fig. 5A, arrowheads in bottom panels).
DISCUSSION
The present study demonstrates that dietary CHO level influences the density of GLP-immunoreactive cells, or L cells, in the chicken ileum. Our previous study using transmission electron microscope images indicated that L cells in the chicken ileum were characterized by a long cytoplasmic process and microvilli on the apical surface. This study also revealed that GLP-1 and GLP-2 are costored in the same secretory granule [28]. These types of EECs, called open-type, act as chemosensors to detect intraluminal nutrients [12]. Furthermore, the microvilli of open-type EECs contain various receptors that respond to chemical signals from contents in the intestinal lumen [2]. Therefore, EECs, immunoreactive for GLP-1 and GLP-2, would function as chemical signal receptors for contents in the lumen.
Several reports have shown that ingested nutrients, such as CHOs, proteins, and fatty acids, have a variety of effects on L cells in the mammalian gut [5, 20, 40]. In addition, L cells release both GLP-1 and GLP-2 in response to glucose in the small intestine of rats [33]. Moreover, changes in dietary components impede GI hormone production, as do changes in EEC prevalence [11]. We have demonstrated that dietary protein and supplementation of amino acids influence the density of L cells in the chicken small intestine [24, 29]. Moreover, our recent study indicated that dietary CHOs positively affected the proliferation of intestinal epithelial cells including enterocytes and goblet cells in the chicken ileum [34]. These findings suggest the close relationship of dietary nutrients with activities of epithelium in the chicken small intestine.
In the present study, the occurrence of GLP-1- and GLP-2-immunoreactive cells was significantly decreased in the two experimental groups, especially in the CHO-free group. Furthermore, GLP-1- and GLP-2-immunoreactive cells exhibited an oval or round shape were observed in the CHO-free group. Such cells were mostly seen in the fasting chicken ileum, where they form vacuoles with tiny lobule nuclei in the perikaryon (our unpublished data). An ultrastructural study demonstrated fasting-induced degeneration in epithelial cell components of the small intestine of White Leghorn chicken [39]. As a consequence of these findings, the phenomenon implies that a lack of luminal CHO leads to L cell degeneration. This might explain why the frequency of GLP-immunoreactive cells decreased in the lower CHO groups in the present study. Our previous studies [17, 24] demonstrated that the effect of dietary protein on GLP-2-immunoreactivee cells was opposite to that on GLP-1-immunoreactive cells. However, there was no difference in the effect of dietary carbohydrate on these cells in this study. This is a reasonable result because GLP-1 and GLP-2 are co-stored in the same secretory granule of the chicken intestinal L cell [28].
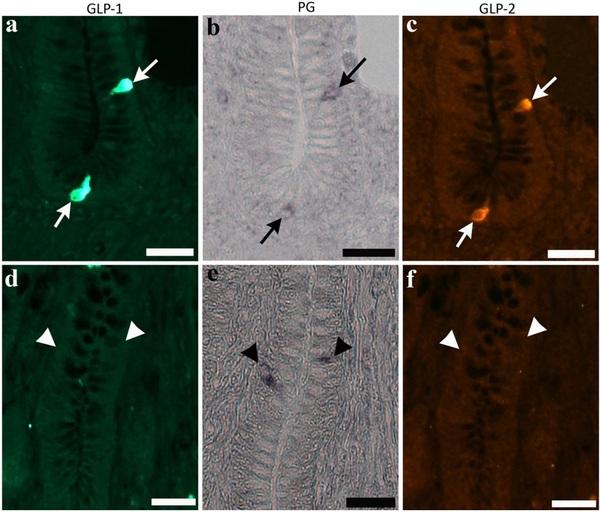
Fig. 4. Photomicrographs indicate cells expressing immunoreactivity for glucagon-like peptide (GLP)-1 (a) and GLP-2 (c), and signal of proglucagon (PG) mRNA (b) in the bottom part of villi of the chicken distal ileum from the control group (arrows in top panels). Same cell types are also observed in the two experimental groups. Cells expressing only PG mRNA signal (arrowheads in bottom panels, e), but not immunoreactivity for GLP-1 (d) and GLP-2 (f), are rarely observed in crypts. Bars: 20 μm.
Prohormone convertase (PC) enzymes mediate tissue-specific post-translational processing, which liberates peptide hormones in a tissue-specific way. PC1/3 enzymatic modifications in intestinal L cells generate GLP-1 and GLP-2 peptide hormones from PG [4, 9, 10]. However, studies conducted on GLUTag cell culture, or GLP-1 secreting cells, have revealed that glucose is the most efficient stimulator for regulating PG transcription [7, 30]. Additionally, various nondigestible CHOs and their derivatives may upregulate the PG transcription level in the distal mammalian gut [5, 41]. Taken together, CHO levels significantly impact the expression of PG mRNA in a variety of animal species. This study demonstrated that the prevalence of cells expressing PG mRNA signal decreased in the CHO-free group compared with that in the control group. Therefore, the present findings suggest that CHO may control PG transcription in the ileum.
Ki-67 is a nonhistone nuclear protein expressed in the G1, S, G2 and M phases of the cell cycle [22], generating its usefulness as a proliferation marker. This study evaluated the coexpression of Ki-67 and GLP-1 in cells from the proximal and distal ileum. The ratio of coexpressive cells was not impacted by dietary CHO level in the proximal ileum. However, the distal ileum showed a decreasing trend in the ratio of coexpressive cells with the reduction of dietary CHO level. A significant difference was measured between the control and CHO-free groups. These findings coincide with the results of PG mRNA expression mentioned above. Our previous findings indicated the upregulating effect of dietary CHO on the proliferation of epithelial cells in the chicken distal ileum [34]. Results of this study, therefore, suggest that dietary CHO also effectively activates L cell proliferation in the chicken distal ileum.
The number of L cells gradually increased from proximal to distal parts of the chicken small intestine, with the highest frequency found in the distal ileum [18, 19]. GLP-2 is a trophic hormone released from L cell and has been shown to stimulate epithelial cell proliferation [8]. Reimann et al. [31] demonstrated that the facilitative glucose transporter2 (GLUT2) was involved in the regulation of the hormone release from rodent L cells. Though there has been no data showing the expression of GLUT2 in chicken intestinal L cells, it is hypothesized that the high number of L cells may secrete higher amount GLP-2 which in turn actively stimulates L cells proliferation in the distal ileum in the present study. This issue will need more systematic investigation in order to be addressed.
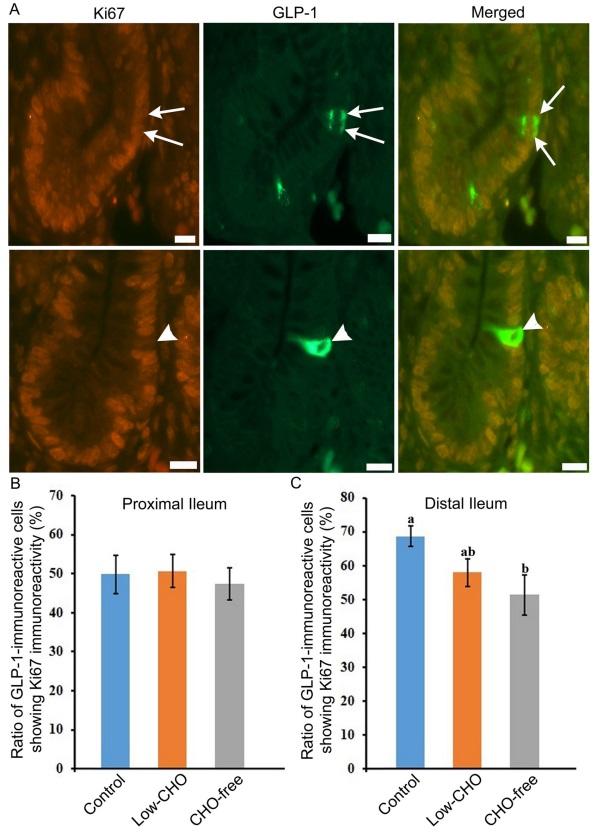
Fig. 5. Glucagon-like peptide (GLP)-1-immunoreactive cells showing immunoreactivity for Ki-67 in the chicken distal ileum from the control group. A: Arrows in top panels indicate GLP-1-immunoreactive cells which also show immunoreactivity for Ki-67. Arrowheads in the bottom panels indicate immunoreactivity for GLP-1 but not for Ki-67. B: The ratio of GLP-1-immunoreactive cells showing Ki-67 in the proximal ileum from three groups. No significant difference in the ratio is observed among the three groups. C: The ratio of GLP-1-immunoreactive cells showing Ki-67 in the distal ileum from the control, low-carbohydrate (CHO), and CHO-free groups. There are significant differences between different alphabets. P< 0.05. Error bars: Standard error. a> b.
Corn oil and cellulose were included at a higher level in all groups’ diets to maintain isoenergic conditions. Obviously, increasing the concentration of these substances had no significant effect on any of the parameters investigated in this study; if they had, they would be seen in the CHO-free group.
In conclusion, dietary CHO could play a key role in stimulating activities and L cell proliferation in the chicken ileum. Indeed, the results presented here provide evidence that dietary CHO level influences the density of L cells in the chicken small intestine.
This article was originally published in Journal of Veterinary Medical Science 84(2): 265–274, 2022. doi: 10.1292/jvms.21-0572. This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial No Derivatives (by-nc-nd) License. (CC-BY-NC-ND 4.0: https://creativecommons.org/licenses/by-nc-nd/4.0/). 








.jpg&w=3840&q=75)