Butyrate: Feeding the Gut and Beyond for Animal Health
Published: January 31, 2014
By: James Pierce, MS, PhD; Nutriad Inc. Elgin, IL, USA; Maja Marien, DVM, PhD; Tim Goossens, PhD (Nutriad International NV Dendermonde, Belgium)
Introduction
In North America, there has been a tremendous body of research in recent years to find the ideal product or program to replace or reduce the use of antibiotics in livestock and poultry production. This is driven by the legislation in the EU and the pressure the health-care system has placed on the use of sub-therapeutic levels of antibiotics for growth promotion. There have been many arguments made regarding the validity of such claims, but ultimately, the consumer will decide what they are willing to accept and pay for accordingly.
With the projected increase in the human population as well as the rise in income over the next 20 years, especially in BRIC countries, there will be a concomitant increase in demand for animal products. The demands to increase the safety and efficiency of food production has become a major focus globally. In recent years, many molecules and additives have been evaluated for their ability to improve animal health, growth, efficiency, and reproduction. The products evaluated have included: prebiotics, probiotics, botanical products, acids (inorganic, organic, short and medium chain fatty acids), chelated minerals, and enzymes.
In order to evaluate these compounds, one should aim at understanding as much as possible about their safety, efficacy, and mode of action. If we are primarily looking to replace antibiotic growth promoters, we should first evaluate the effect on digestion and gut health. The intestine is the largest organ in the body and thus involves a complex system in which several factors influence the final outcome. In the intestine, three major components (the mucosal barrier, the composition of the microbiota and the local immune system) provide defensive measures against different pathogens through permanent contact and communication with each other. Feed additives may interact with host cells (intestinal cells, immune cells), with the host’s microbiota, or with pathogens impairing the normal intestinal function.
Butyrate – Proposed modes of action
Butyrate is somewhat unique among its biological functions when compared with other short chain fatty acids (SCFA). The (SCFA) constitute a group of molecules that contain from one to seven carbon atoms and which exist as straight or branched-chain compounds: predominant SCFAs are acetic, propionic and butyric acid. Because SCFA are weak acids with a pK of ≤4.8 and the pH of the gastrointestinal tract (GIT) is nearly neutral, 90-99% of the SCFA are present in the GIT as anions rather than free acids.
Among the SCFA, butyric acid has received particular attention. Butyric acid is available in the salt form of Na, K, Mg or Ca. The advantage of salts over free acids is that they are generally odorless and easier to handle in the feed manufacturing process owing to their solid and less volatile form. For the purpose of the present article, the term ‘butyrate’ is used interchangeably for the acid, the salt and the anion forms.
Fig 1. Absorption of n-butyrate in the large intestine and subsequent metabolism. Butyrate transport with monocarboxylate transporters (MCT) are saturable and coupled with H+ transport. Butyrate is recognized by G-protein receptors (GPR41, GPR43). (Adapted from Guiloteau et al., 2010).
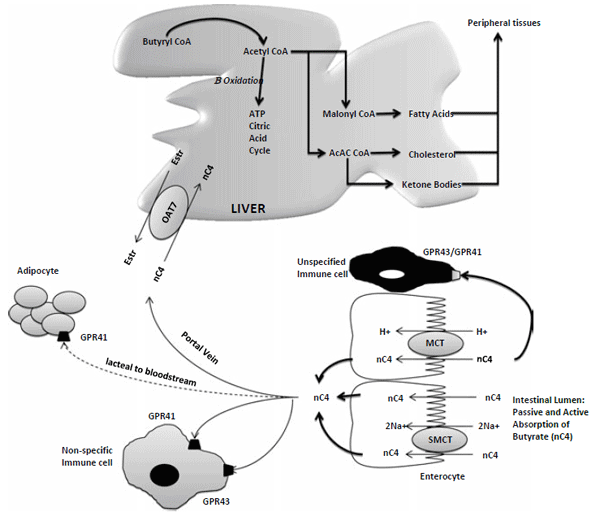
There are several potential fates of butyrate once consumed by the animal. Butyrate may function as a ligand for transmembrane receptors, as a modulator of gene activity, and as a direct energy source for cellular metabolism via oxidation. Several cell types, many of which associated with the digestive tract, have been described to be receptive to one or more of these biological effector functions, which explains the extensive range of biochemical effects that have been documented to be mediated by butyrate (Fig 1).
When butyrate is present in the blood stream or in the proximal parts of the intestinal tract, it induces the production of host defense peptides (Guilloteau et al., 2009). These peptides stimulate the development and repair of the intestinal tract through an increase in cell proliferation (Bartholome et al., 2004). Recently it has been shown that butyrate, when present in blood, stimulates a peptide that increases the absorption of glucose from the intestine. Indications that a similar mode of action can be expected in poultry, is shown by Hu and Guo (2007), who found an increased development of the villi when sodium-butyrate was added to the diet.
Butyrate also has been shown to stimulate several functions in the lower part of the intestinal tract. Studies have identified specific G-protein-coupled receptors, specifically GPR 41 and GPR 43, on gut epithelial cells in the epithelium of particularly the ileum, caeca and colon (Le Poul et al., 2003). When butyrate is attached to these receptors the production of several different peptides is stimulated (Cox et al., 2009, Tazoe et al., 2008). Some of these peptides have a positive effect on the development of the immune system and improve the functioning of the immune system in case of a health challenge (Cox et al., 2009). Other peptides have been shown to optimize gut motility, by reducing the rate of feed passage (Tazoe et al., 2008). In poultry, the emptying of the feed out of the gizzard into the small intestine is slowed down. Thus, it seems that butyrate is inducing a similar effect to passage rate as coarse particles such as oyster shell.
Indications that butyrate also stimulates the immune system in poultry were obtained by Leeson et al. (2005) in that, birds previously fed butyrate showed more ability to withstand against the stress of coccidial challenge at 21 d of age. Weber (2008) found when pigs where challenged with Escherichia coli lipopolysaccharide (LPS), sodium butyrate increased the magnitude of the cortisol response and increased skeletal muscle IL-6 mRNA expression, also indicating that dietary butyrate affects the response to inflammatory stimuli.
In summary, beneficial effects include, among others, stimulation of digestive enzyme production, enhanced development of intestinal villi, reduction of acute inflammatory responses, increased GIT retention time, inhibition of cancer cell growth and the secretion of host defense peptides (Guilloteau et al., 2012). Apart from effects in eukaryotic host cells, butyrate is also described to have an impact on the activity of prokaryotes residing in the animal’s GIT. For example, it has been shown to affect the colonization of Salmonella and Campylobacter and to influence the composition of the gut microbiota (see below).
Butyrate - Experimental results
Mucosal barrier
Dietary supplementation of butyrate has been shown to support enteric development and intestinal health of neonatal animals. At weaning, the small intestine of the piglet generally undergoes a decreased capacity of absorption that is associated with a marked reduction in villous height and crypt depth. These changes are accompanied with a decreased feed intake and poor growth (Piva et al., 2002; Pluske et al., 1996). Butyrate stimulates epithelial cell proliferation resulting in a larger absorptive surface, leading to improved feed utilization. Furthermore, butyrate in the weaner diet preserves villus length and thereby helps to maintain feed intake. Effect of butyrate on gut morphology is of great biological value to the weaning period when the weight of the small and large intestine increases three times faster than that of the whole body mass.
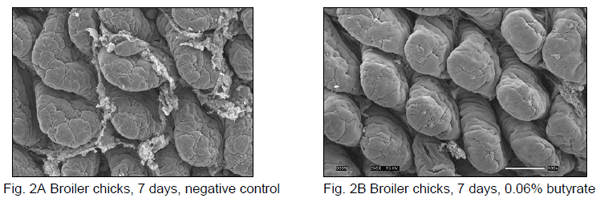
The following trial results (Ferket et al., 2010) were obtained in broilers, but do show the principle of the impact of butyrate supplementation on intestinal development and growth performance in young animals. Commercial broilers were randomly assigned to 32 floor pens containing 30 birds each and provided feed and water ad libitum until 49 days. Starter feed (pellet-crumbled) treatments consisting of 3 dietary supplementation levels of coated butyrate (0.0, 0.015, 0.03, and 0.06% butyrate) were subjected to 8 replicate pens per treatment from 1 – 14 days. Subsequently, all birds were fed common grower and finisher diets in pelleted form. Body weight (BW) and feed intake was determined at 7, 14, 21, 42, and 49 days and feed/gain (FCR) calculated. At 3, 8, and 14 days, 4 birds/treatment were sampled for gut histology evaluation. BW at 14 days increased linearly (p<0.01) as the level of butyrate increased (457 g vs. 470 g for 0 vs. 0.06% butyrate), but no effects on 1-14 days FCR were observed. Histomorphometic analysis correlated with early treatment effects on BW (see Fig 2A and 2B). The positive starter feed treatment effects was observed throughout the experiment, with 0.015% butyrate resulting in a 3% and 2% improvement in 42 days (p<0.02) and 49 days (p<0.10), respectively. A linear improvement in 1-42 days FCR by up to 3% was also observed as the level of butyrate increased in the starter feeds. Dietary supplementation of coated butyrate in starter feeds showed to have a lasting positive effect on broiler growth performance.
Modulation of microbiota and impact on pathogens
Studies done by Galfi and coworkers (Galfi et al., 1991) have shown that butyrate increases the number of intestinal lactic acid and lactobacilli in butyrate-fed pigs, while decreasing the number of coliforms and E. coli. In his PhD-thesis Pérez Gutiérrez (2010) investigated microbial composition after including different additives in the feed of (weaning) pigs. The qPCR-data revealed an increase in lactobacilli in the butyrate group, while weaning piglets that received butyrate also had a more homogeneous microbial profile, which was regarded as a positive effect.
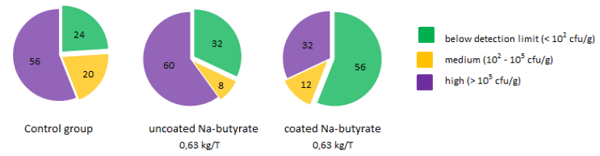
In studies of Van Immerseel and co-workers it was shown that butyrate, when present in the intestinal tract, was able to decrease Salmonella colonisation in broilers (Van Immerseel et al., 2005). The mode of action of this activity of butyrate seems at least partially mediated through modulation of gene expression. Butyrate specifically down-regulates Salmonella Pathogenicity Island 1 (SPI-1) gene expression, hereby preventing invasion of intestinal epithelial cells, one of the important steps of Salmonella pathogenesis in the bird (Gantois et al., 2006). The importance of an effective coating in order to get significant reduction in Salmonella colonisation in the ceca and internal organs in vivo was also shown in Fig 3. (Van Immerseel et al., 2005).
Fig. 3: Percentage of animals with a certain amount of Salmonella in caecum (n=25 per group).
A trial done at the University of Bologna (Bosi et al., 2010) was done to look at the possible impact of butyrate (coated and uncoated) on E. coli K88 (ETEC) infection in piglets. Fifty-four piglets (prone to ETEC intestinal adhesion) weaned at 21-28 days were used (three control groups of six animals, three challenged groups of 12 animals, balanced for litter and weight). Experimental diets were obtained with the addition of free or fat coated sodium butyrate. Full factorial design of two factors (four diets x challenge (E. coli K88) – yes/no), so groups were: 1) control diet, unchallenged; 2) 2 kg/T uncoated butyrate, unchallenged; 3) 2 kg/T coated butyrate, unchallenged; 4) control diet, challenged; 5) 2 kg/T uncoated butyrate, challenged; 6) 2 kg/T coated butyrate, challenged. The challenge with ETEC increased mortality in the control group (15% mortality) and had an impact on the growth rate of the piglets. The piglets receiving the uncoated sodium butyrate showed a lower mortality rate (5%) and were to a lesser extent influenced by growth than the challenged control group. In the group offered the coated sodium butyrate in the diet, none of the piglets died and the growth rate was only marginally influenced.
Butyrate - More than just an active compound
The effects on immune development, gut motility as well as the Salmonella effect via gene expression are only possible when the butyrate arrives in the lower parts of the intestinal tract. It is generally accepted that unprotected butyrate is quickly absorbed in the proximal part of the intestinal tract. Therefore, in order to get butyrate in significant levels available in the lower part of the intestinal tract the butyrate needs to be protected to achieve a target release.
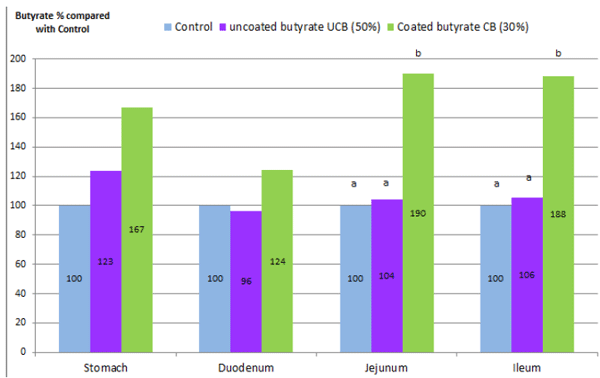
In recent research done at the University of Illinois (Stein et al, 2012) the importance of a good coating was again demonstrated. The researchers investigated the disappearance kinetics of different sources of butyrate in diets fed to weanling pigs. Weanling pigs (n = 24; 8.0 ± 0.5 kg BW) were randomly allotted to 3 dietary treatments (6 replicate pigs per dietary treatment): 1) a control diet, 2) the control diet + 4kg/T of an uncoated butyrate (50% product, UCB), 3) the control diet + 4kg/T of a coated butyrate (30% product, CB). The dietary treatments were provided to pigs daily for 7 d as 3 times the estimated energy requirement for maintenance. On the last day of the experiment, all pigs were euthanized to collect samples of contents in the stomach, duodenum, jejunum, ileum, cecum, and proximal and distal colon. Concentrations of dry matter (DM) and butyrate were analyzed in all samples. There was a similar pattern of the concentration of butyrate in the digestive tract, indicating that the concentration of butyrate was greater in the stomach than in the duodenum and jejunum, and gradually increased in the ileum. Weanling pigs fed the CB diet had greater (P < 0.05) concentrations of butyrate in the jejunum (190 % μg butyrate/g digesta DM) and ileum (188% μg butyrate/g digesta DM) than weanling pigs fed the control diet (μg butyrate/g digesta DM put as 100%). In the cecum and colon, endogenous production was more variable among piglets and therefore no significant differences could be demonstrated. In conclusion, the coated butyrate in spite of lower percentage of butyrate (30%), increased significantly the concentrations of butyrate in the lower intestinal contents of pigs. Supplementation of the more concentrated (50%) but less protected butyrate did not result in significant differences in the amount of butyrate in the intestinal tract compared with the control group.
Fig. 4: Amount of butyrate in % compared to the unsupplemented control group.
Butyrate is a molecule of great interest to both human and veterinary medicine. Owing partly to the interest from human medicine, more and more profound knowledge on the mode of action has been obtained which allows for more sound advices to producers. In the current paper an attempt is made to summarize some of the newest developments on the research as well as to show experimental results in production animals; because, in the end the benefits to the efficiency of animal production are what matter. As shown above (Fig 4), butyrate has an effect on several levels (mucosal barrier, feed passage, microbiota, immune system, pathogens and others) and this combination of effects contribute to its general acceptance as helping for improved health as well as for improved performance.
References
Bartholome, A.L.,D.M. Albin, DH Baker, J.J. Holst, and K.A. Tappenden. 2004. Supplementation of total parental nutrition with butyrate increases structural aspects of intestinal adaptation after an 80% jejunoileal resection in neonatal piglets. JPEN. 28(4): 210-222.
Cox, M.A., J. Jackson, M. Stanton, A. Rojas-Trian, L. Bober, M. Laverty, X. Yang, F. Zhu, J. Liu, S. Wang, F. Monsma, G. Vassileva, M. Maguire, E. Gustafson, M. Bayne, C. Chou, D. Lundell, and C. Jenh. 2009 Short-chain fatty acids act as anti-inflammatory mediators by regulating prostaglandin E2 and cytokines. China Natl. J. New Gastroenterol. 15(44): 5549-5557.
Galfi, P., S. Neogradi. and T. Sakata. 1991. Effects of volatile fatty acids on the epithelial cell proliferation of digestive tract and its hormonal mediation. In: Tsuda, T., Y. Sasaki, and R. Kawashima editors, Physiological aspects of digestion and metabolism in ruminants, Academic Press, Orlando, Florida, 49-59.
Gantois, I., R. Ducatelle, F. Pasmans, F. Haesebrouck, I. Hautefort, A. Thompson, J.C. Hinton, and F. Van Immerseel. 2006. Butyrate specifically down-regulates Salmonella pathogenicity island 1 gene expression. Appl Environ Microbiol. 72 (1): 946-949.
Guilloteau, P., R. Zabielski, J.C. David, J.W. Blum, J.A. Morisset, M. Biernat, J. Wolinski, D. Laubitz, and Y. Hamon. 2009. Sodium-butyrate as a growth promoter in milk replacer formula for young calves. J. Dairy Sci. 92: 1038-1049.
Guilloteau, P., L. Martin, V. Eeckhaut, R. Ducatelle, R. Zabielski., and F. Van Immerseel. 2012. From the gut to the peripheral tissues: the multiple effects of butyrate. Nutr Res Rev. 23(2):366-84.
Hu, Z. and Y. Guo. (2007). Effects of dietary sodium butyrate supplementation on the intestinal morphological structure, absorptive function and gut flora in chickens. Animal Feed Science and Technology, 132 (3): 240-249.
Leeson, S., Namkung, H., Antongiovanni, M. and Lee, E.H. (2005). Effect of butyric acid on the performance and carcass yield of broiler chickens. Poult Sci. 84: 1418-1422.
Le Poul, E., C. Loison,S. Struyf, J.Y. Springael., V. Lannoy, M. Decobecq, S. Brezillon, V. Dupriez, G. Vassart, J. Van Damme, M. Parmentier, and M. Detheux. 2003. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J Biol Chem. 278(28): 25481-25489.
Pérez Gutiérrez, O. 2010. Unraveling piglet gut microbiota dynamics in response to feed additives. Thesis submitted in fulfillment of the requirements for the degree of doctor at Wageningen University, The Netherlands.
Piva, A., M. Morlcchini, G. Casadie, P.P. Gatta, G. Biagi, and A. Prandini. 2002. Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Ital. J. Anim. Sci. 1: 35-41.
Pluske, J.R., M.J. Thompson, C.S. Atwood, P.H. Bird, I.H. Williams, and P.E. Hartmann. 1996. Maintenance of villus height and crypt depth, and enhancement of disaccharide digestion and monosaccharide absorption, in piglets fed on cows' whole milk after weaning. Br J Nutr. 76 (3):409-22.
Scheppach W. and F. Weiler. 2004. The butyrate story: old wine in new bottles? Curr Opin Clin Nutr Metab Care. 7: 563–567.
Sossai, P. 2012. Butyric acid: what is the future for this old substance? Swiss Med Wkly. June 6;142:w13596. doi: 10.4414/smw.2012.13596.
Tazoe, H., Y. Otomo, L. Kaji, R. Tanaka, S.I. Karaki, and A. Kuwahara. 2008. Roles of short-chain fatty acids receptors, GPR41 and GPR43, on colonic functions. Acta Physiol Pol. 59 : 251-262.
Van Immerseel, F., F. Boyen, I. Gantois,L. Timbermont, L. Bohez, F. Pasmans, F. Haesebrouck, and R. Ducatelle 2005. Supplementation of coated butyric acid in the feed reduces colonization and shedding of Salmonella in poultry. Poult Sci. 84(12): 1851-1856.
Weber, T.E., and B.J. Kerr. 2008. Effect of sodium butyrate on growth performance and response to lipopolysaccharide in weanling pigs. J. Anim Sci. 86: 442-450.
This paper was presented at the XVI AMENA Congress, Puerto Vallarte, Mexico, October 2013
Related topics
Authors:

Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post
14 de marzo de 2014
Dr.Mane, yes, we recommend Na-Butyrate
Rafa, I understand your question, but do not have a definite answer. The coated Na-butyrate we use is released along the small intestine where it can provide direct energy to the enterocytes. What you are suggesting with fibers would probably work best in older pigs, especially reproducing sows.
Thank you both for your interest and let me know if I can answer any further questions.
James

IFF - International Flavors & Fragrances
17 de febrero de 2014
Hello Dr. Pierce. My question correlates very much with pigs more than poultry. How effective could be feeding some fibres which could eventually scape small intestinal digestion and being digested/fermented in the caeca-hind gut, producing effective butyrate?. If this was the case, what sort of fibre would you recommend to attain this objective?. Many thanks from Madrid, Rafa.

14 de febrero de 2014
What should be the composition of BUTYRATE in diet and the schedule of feeding?
Can we prefer sodium n-butyrate for the trial?







You may be interested in
.jpg&w=3840&q=75)