1. Introduction
Breeding programmes are designed to produce a broiler with high potential for growth, yield, and feed efficiency. These traits may compromise health status under husbandry designed to maximise this genetic potential, since there is a trade-off between growth- and health-related traits with responses to husbandry programmes, age, sex, and genetic line (Siegel et al., 2001). In this respect, it is well known that chick viability is an important factor determining the profitability of the poultry industry. From fertilised ovum to the placement at the broiler farm, various factors, such as incubating egg quality, egg storage conditions, incubation conditions and conditions that exist between hatching and placement at the farm and their interactions are important determinants of chick quality (Decuipere et al., 2001). In contrast to mammals, chicken embryonic development takes place in a semi-closed system, the egg, where only exchange of gas and water takes place. It has been appreciated recently that the success of embryonic development depends on egg composition and conditions of egg incubation. Therefore, a great body of evidence indicates that avian maternal nutrition is the major determinant of the health and development of the progeny similar to humans and other mammals. The egg composition is designed in such a way that all nutrients necessary for the development of the future embryo are accumulated within the egg yolk and white (Speake et al., 1998; Surai et al., 1999d). Among different nutrients in the maternal diet which could significantly affect chick embryo development and their viability in the early posthatch life, natural antioxidants have been suggested to play a central role. In fact, the high levels of endogenous antioxidants within the egg and embryonic tissues can clearly serve as a major adaptive mechanism for the protection of the tissue during the oxidative stress experienced at hatching. Therefore, the aim of this review is a critical analysis of data related to antioxidant defences during chick embryonic development and nutritional means of their regulation.
2. Detrimental and regulatory roles of free radicals and antioxidants
Free radicals are atoms, molecules or any compounds containing one or more unpaired electrons. Most biologically-relevant free radicals are derived from oxygen and nitrogen. Both these elements are important for animal life, but in some circumstances they can be converted (deliberately or by chance) into free radical molecules. Free radicals are highly unstable, very reactive and are capable of damaging any biological molecules including DNA, proteins, lipids and carbohydrates. Damage to DNA is associated with mutations, translation errors, and disruption of protein synthesis. In some cases, damage to DNA leads to cancer. Damage to proteins causes modifications in ion transport and receptor functions, as well as altered enzymatic activities. Polyunsaturated fatty acid oxidation alters membrane composition, structure and properties (fluidity, permeability, etc.) and activity of membrane-bound enzymes (Surai, 2002a, 2006). The damage to biological molecules ultimately compromises growth, development, immunocompetence and reproduction. Cells are under constant attack by free radicals, many of which are formed as a natural consequence of normal metabolic activity and as part of the immune system's strategy for destroying invading microorganisms. For example, under normal physiological conditions about 3% to 5% of the oxygen taken up by the cell undergoes univalent reduction leading to the formation of free radicals (Singal et al., 1998). About 1012 O2 molecules processed by each rat cell daily and the leakage of partially reduced oxygen molecules is about 2%, yielding about 2 x 1010 molecules of reactive oxygen species (ROS) per cell per day (Chance et al., 1979). Furthermore, Helbock et al. (1998) have shown that the DNA in each rat cell is hit by about 100,000 free radicals a day; and each cell sustains as many as 10,000 potentially mutagenic (if not repaired) lesions per day arising from endogenous sources of DNA damage (Ames and Gold, 1997). Some oxidative lesions escape repair, and the steady state level of oxidative lesions increased with age. An old rat had accumulated about 66,000 oxidative DNA lesions per cell (Ames, 2003). An interesting calculation has been made by Halliwell (1994). He assumed that in mitochondria about 1% to 3% of oxygen consumed may leak from the electron transport chain to form superoxide radical and took into account that an adult at rest utilises approximately 3.5 mL O2/(kg·min) or 352.8 L/day (assuming 70 kg body mass) or 14.7 mol/day. Therefore, if 1% form superoxide this would be 0.147 mol/day or 53.66 mol/yr or about 1.72 kg/yr of superoxide radical. In stress conditions this amount would be substantially increased. In addition, to free radicals formed as a natural consequence of oxygen metabolism, immune cells produce free radicals as a means to destroy pathogens (Kettle and Winterbourn, 1997). These calculations show that free radical production in the body is substantial and underscore the fact that thousands of biological molecules can be easily damaged if not protected.
For the majority of organisms on earth, life without oxygen is impossible, animals, plants and many microorganisms relying on oxygen for the efficient production of energy. However, the high oxygen concentration in the atmosphere is potentially toxic for living organisms. During evolution living organisms have developed specific antioxidant protective mechanisms to deal with ROS and reactive nitrogen species (RNS). Therefore, it is only the presence of natural antioxidants in living organisms which enable them to survive in an oxygen-rich environment. These mechanisms are described by the general term “antioxidant system”. It is diverse and responsible for the protection of cells from the actions of free radicals. This system includes (Surai, 2002a, 2006):
- natural fat-soluble antioxidants (vitamins A, E, carotenoids, ubiquinones, etc.);
- water-soluble antioxidants (ascorbic acid, uric acid, taurine, etc.);
- antioxidant enzymes: glutathione peroxidase (GSH-Px), catalase (CAT) and superoxide dismutase (SOD);
- thiol redox system consisting of the glutathione system (glutathione/glutathione reductase/glutaredoxin/glutathione peroxidase and a thioredoxin system (thioredoxin/thioredoxin peroxidase/thioredoxin reductase).
The protective antioxidant compounds are located in organelles, subcellular compartments or the extracellular space enabling maximum cellular protection to occur. Thus antioxidant system of the living cell includes three major levels of defence. The first level of defence is responsible for prevention of free radical formation by removing precursors of free radicals or by inactivating catalysts and consists of three antioxidant enzymes namely SOD, GSH-Px and CAT plus metal-binding proteins. Since the superoxide radical is the main free radical produced in physiological conditions in the cell, SOD (EC 1.15.1.1) is considered to be the main element of the first level of antioxidant defence in the cell. This enzyme dismutates the superoxide radical in the following reaction: 2O¯2 + 2H+ + SOD→ H2O2 + O2. The hydrogen peroxide formed by SOD action can be detoxified by GSH-Px or CAT which reduces it to water. Transition metal ions also accelerate the decomposition of lipid hydroperoxides into cytotoxic products such as aldehydes, alkoxyl radicals and peroxyl radicals. Therefore, metal-binding proteins (transferrin, lactoferrin, haptoglobin, hemopexin, metallothionenin, ceruloplasmin, ferritin, albumin, myoglobin, etc.) also belong to the first level of defence. It is necessary to take into account that iron and copper are powerful promoters of free radical reactions and therefore their availability in “catalytic” forms is carefully regulated in vivo. Therefore, organisms have evolved to keep transition metal ions safely sequestered in storage or transport proteins. In this way, the metal-binding proteins prevent formation of hydroxyl radical by preventing them from participation in radical reactions. Unfortunately, this first level of antioxidant defence in the cell is not sufficient to completely prevent free radical formation and some radicals do escape through the preventive first level of antioxidant safety screen initiating lipid peroxidation and causing damage to DNA and proteins. Therefore, the second level of defence consists of chain-breaking antioxidants e vitamin E, coenzyme Q (CoQ), carotenoids, vitamin A, ascorbic acid, uric acid and some other antioxidants. Glutathione and thioredoxin systems also have a substantial role in the second level of antioxidant defence. Chain-breaking antioxidants inhibit peroxidation by keeping the chain length of the propagation reaction as small as possible. Therefore, they prevent the propagation step of lipid peroxidation by scavenging peroxyl radical intermediates in the chain reaction. However, even the second level of antioxidant defence in the cell is not able to prevent damaging effects of ROS and RNS on lipids, proteins and DNA. In this case, the third level of defence is based on systems that eliminate damaged molecules or repair them. This level of antioxidant defence includes lipolytic (lipases), proteolytic (peptidases or proteases) and other enzymes (DNA repair enzymes, ligases, nucleases, polymerases, proteinases, phospholipases and various transferases). All these antioxidants are operating in the body in association with each other forming an integrated antioxidant system. The co-operative interactions between antioxidants in the cell are vital for maximum protection from the deleterious effects of free radicals and toxic products of their metabolism. For example, it is well established that vitamin E is the major antioxidant in biological membranes, the “headquarters” of antioxidant network. However, it is usually present there in low molar ratios (one molecule per 2,000 to 3,000 phospholipids) but vitamin E deficiency is difficult to induce in adult animals. It is probably due to the fact that oxidized vitamin E can be converted back into the active reduced form by reacting with other antioxidants: ascorbic acid, glutathione, ubiquinols or carotenoids (Surai, 2006). Therefore, the antioxidant protection in the cell depends not only on vitamin E concentration and location, but also relies on the effective recycling. Indeed, if the recycling is effective then even low vitamin E concentrations are able to maintain high antioxidant protection in physiological conditions. Antioxidant recycling is the most important element in understanding mechanisms involved in antioxidant protection against oxidative stress. The rate of regeneration, or recycling, of the vitamin E radicals may affect both its antioxidant efficiency and its lifetime in biological systems.
- As can be seen from data presented above the antioxidant defence includes several options (Surai, 2015a,b,c,d,e; Surai and Fisimin, 2015a,b):
- decrease localised oxygen concentration;
- decrease activity of pro-oxidant enzymes and improve effi- ciency of electron chain in the mitochondria and decreasing electron leakage leading to superoxide production;
- prevention of chain initiation by scavenging initial radicals due to inducing various transcription factors (e.g., NF-E2-related factor 2 [Nrf2], nuclear factor-kB [NF-kB] and others) and ARErelated synthesis of AO enzymes (SOD, GSH-Px, CAT, glutathione reductase [GR], glutathione S-transferase [GST], etc.);
- binding metal ions (metal-binding proteins) and metal chelating;
- decomposition of peroxides by converting them to non-radical, nontoxic products (Se-GSH-Px);
- chain breaking by scavenging intermediate radicals such as peroxyl and alkoxyl radicals (vitamins E, C, GSH, uric acid, ubiquinol, bilirubin, etc.);
- repair and removal of damaged molecules (methionine sulfoxide reductase, DNA-repair enzymes, chaperons, etc.);
- redox-signalling and vitagene activation with synthesis and increased expression of protective molecules (GSH, thioredoxins, SOD, heat shock proteins [HSPs], sirtuins, etc.);
- antioxidant recycling mechanisms, including vitamin E recycling;
- apoptosis activation and removal terminally damaged cells and restriction of mutagenesis.
It is important to realise that all antioxidants in the body function in concert to provide antioxidant defence. The role of each member of the antioxidant ‘team’ is well defined. For example, selenium is part of several different selenoproteins that regulate antioxidant defence in different tissues and in different ways. Vitamin E, carotenoids and CoQ provide antioxidant defence in lipids, specifically in biological membranes. The function of each is important to the effectiveness of the other antioxidants. For example, vitamin E potency is very much dependent on the efficacy of its recycling. Indeed, vitamin C ‘recycles’ vitamin E from oxidised form to become active again, while glutathione does the same for vitamin C. Furthermore, vitamins B1 and B2 are also involved in vitamin E recycling. It is important to mentioned that ROS are no longer viewed as just toxic by-products of mitochondrial respiration, but are now appreciated for their role in regulating various cellular signalling pathways (Reczek and Chandel, 2015). Indeed, the adaptation to stressful conditions of our life is mediated via vitagene network in the body. If the diet is balanced and sufficiently provided with dietary antioxidants and antioxidant nutrients, then even low doses of such antioxidants as vitamin E are effective. On the other hand, under conditions of oxidative stress where free radical production dramatically increases, then without an external help it is difficult to prevent damages to major organs and systems. This external help takes the form of increased dietary supplementation of natural antioxidants, especially vitamin E, carotenoids and selenium. For the nutritionist or feed formulator the challenge is to understand when and how much external ‘help’ is needed and economically justified.
3. Antioxidant defences of the developing embryo
For a precocial species like the chicken, metabolic rate and oxygen consumption increase rapidly during and just after hatching to meet the demands of endothermy and locomotion, supported by the transition from chorioallantoic to pulmonary respiration (Hohtola, 2002). To cope with this sudden exposure to oxidative stress, it would clearly help if an effective antioxidant system was already in place. Indeed, chick embryo tissues contain a high proportion of highly polyunsaturated fatty acids in the lipid fraction (Speake et al., 1998; Surai et al., 1999d) and therefore need antioxidant defence (Surai et al., 1996). The antioxidant system of the newly hatched chick includes the antioxidant enzymes SOD, GSHPx, CAT (Surai, 1999a), water-soluble antioxidants ascorbic acid (Surai et al., 1996) and glutathione (Surai, 1999a), fat-soluble antioxidants vitamin E, carotenoids (Surai et al., 1996, 1999a, 2001d; Surai, 1999b; Surai et al., 2003; Surai, 2012a, 2014), CoQ (Karadas et al., 2011) as well as selenium (Surai, 2000, 2006).
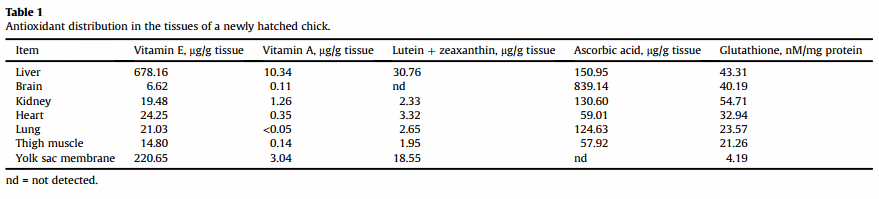
Ascorbic acid is shown to be absent from the initial yolk (Surai et al., 1996) but is synthesised in the yolk sac membrane (YSM) of the chick embryo (Yew, 1985). In our study, the tissue concentration of ascorbic acid in the YSM was the highest at the earliest developmental stage studied, i.e. day 8, and decreased continuously thereafter. The brain displayed a marked increase in ascorbic acid content between days 8 and 13 and was maintained at a high level throughout development (Surai et al., 1996). In fact the concentrations of ascorbic acid in the brain during the final week of embryonic development were far higher than in any other tissue studied; respectively 4.0-, 5.2-, 6.0-, 6.7- and 14.3-fold higher than in liver, kidney, lung, heart, and thigh muscle by 1 day after hatching. It is interesting to note that in the brain of the newly hatched chick, the highest ascorbic acid concentration was found in cerebrum and the lowest e in the cerebellum (Surai et al., 1999a). In accordance with ascorbic acid concentrations tissues of the newly hatched chicks can be placed in the following descending order: brain » liver = kidney = lung > heart = thigh muscle (Surai et al., 1999a). The concentration of GSH, a primary cell water-soluble antioxidant, gradually decreases throughout development. In fact, in the liver, brain and YSM, the greatest amounts of GSH were observed at day 10, with lower amounts after that time. Concentrations of GSH in kidney, lung, heart and skeletal muscle were relatively constant between day 15 and hatching (Surai, 1999a). In newly hatched chicks, based on GSH concentrations tissues can be placed in the following descending order: kidney = liver = brain > heart > lung = thigh muscle » YSM (Surai et al., 1999a). Indeed, at hatching time GSH concentration was 55.49 ± 1.86 nmol/mg protein which was similar to that in the brain (47.71 nmol/mg protein), heart (51.13 nmol/mg protein), kidney (60.65 nmol/mg protein), but much higher than that in the lung (24.43 nmol/mg protein) or muscles (29.49 nmol/mg protein) (Surai, 1999a). Glutathione is considered to be critical in protection against embryotoxicants (Tiboni et al., 1997). After hatching, hepatic GSH concentration in broiler chickens, quail and turkey increased with age (Enkvetchakul et al., 1995).
Coenzyme Q is the only fat-soluble antioxidant synthesised in body, originating from endogenous synthesis and from food. There are two major forms of CoQ in the human tissues namely CoQ10 comprising 95% to 97% in heart, kidney, liver and spleen and 92% in brain and 87% in lung (Dallner and Sindelar, 2000), the remainder, CoQ9 as an antioxidant component of the chicken antioxidant system has not been characterised. There are only limited data available on the CoQ10 distribution in the chicken tissues. In particular, in newly hatched chicks the liver and YSM are characterised by the highest CoQ10 concentrations (64.8 to 67.5 µg/g), while CoQ10 concentrations in the heart (36.2 µg/g), leg muscles (22.4 µg/g), brain (13.0 µg/g), breast muscles (11.9 µg/g) and lung (9.2 µg/g) were shown to be significantly lower (Karadas et al., 2011). It has been shown that CoQ10 concentration in the liver of newly hatched quail (80.1 µg/g; Karadas et al., 2006a) is similar to that reported for chicken (Karadas et al., 2011).
Vitamin E (Surai, 1999b, 2002a), carotenoids (Surai et al., 2001a; Surai, 2012b) and selenium (Surai, 2000, 2002a,b,c, 2006; Surai and Fisinin, 2014) are transferred from feed into egg and further to embryonic tissues and their levels in eggs and embryonic tissues can be regulated by dietary means. Glutathione and antioxidant enzymes GSH-Px, SOD and CAT are also expressed in the embryonic tissues at various stages of their development (Surai, 1999a; Surai et al., 1999a).
Our results indicate that there are tissue-specific features in antioxidant defence strategy during embryonic development of the chicken and SOD plays a crucial role as an integral part of the antioxidant network. Our data indicate that in the embryonic liver, SOD specific activity was maximal at day 11 but decreased sharply by day 15 and remained relatively constant thereafter. In great contrast, the specific activity of SOD in the brain from day 15 onwards was comparatively high being approximately 2 times higher than that in the liver (Surai, 1999a). The specific activities of SOD in kidney, lung, heart and skeletal muscle showed a gradual decrease between day 15 and hatching. In fact, the tissues of newly hatched chicks displayed a considerable variation in the Mn-SOD activity, with the heart having the highest value and lung the lowest (Surai et al., 1999a). By contrast, the lung was characterised by high Cu,ZnSOD activity; in the heart, activity of Cu,Zn-SOD was comparable to the other tissues. Based on the total SOD activity the tissues could be placed in the following descending order: heart > muscle > YSM > kidney > lung > liver (Surai et al., 1999a). Mn-SOD is shown to be the main enzymatic form in the liver and heart comprising 73.2% and 68% of the total SOD activity respectively. In great contrast, in the lung, YSM and thigh muscle, SOD is represented mainly by Cu,Zn-SOD comprising 98.5%, 98.3% and 84.7% of the total SOD activity, respectively. In various parts of the brain (cerebrum, cerebellum, brain stem and optic lobes) of the newly hatched chick the Cu,Zn-SOD is main form of the enzyme being almost 2- fold higher than that of Mn-SOD (Surai et al., 1999a). It should be mentioned that in the kidney both SOD forms are equally represented. Furthermore, the tissues differed markedly in the GSH-Px activities. In all the tissues, Se-dependent GSH-Px was the main enzymatic form, comprising from 65% (lung) up to 90% (heart) of the total enzyme activity. The liver and kidney displayed the highest total GSH-Px activity and the muscle the lowest. As in the case of GSH-Px, CAT activity was also maximal in the liver and kidney (Surai et al., 1999a). Comparison of GSH-Px activity in various tissues showed that the highest activity was found in the embryo liver at all stages of development (Surai, 1999a). At the early stages of embryo development (10 to 11 days) GSH-Px activity was 1.2 to 1.3 times higher compared with the yolk sac membrane and 2.5 to 2.6 times higher than that in the brain. At day 17 of development it was higher by 4.0, 2.8, 2.0, 1.7, 1.3 and 1.2 times compared to the brain, skeletal muscle, heart, YSM, kidney and lung, respectively. In general, changes in GSH-Px activity patterns during the development were very similar to those found for vitamin E, carotenoids (Surai et al., 1996) and vitamin A (Gaal et al., 1995). Probably GSH-Px plays a primary role in antioxidant defence of the liver by effectively removing hydrogen peroxide and lipid hydroperoxides from cells whereas CAT may be less important, decreasing throughout the development. Nevertheless, the concentration of GSH, a primary cell water-soluble antioxidant, gradually decreases throughout the development. Catalase had two peaks of activity at day 10 of embryonic development and in day old chicks. After hatching the liver displayed CAT activity similar to that in the kidney but much higher compared to other tissues (Surai, 1999a).
It is necessary to underline that maternal diet composition is a major determinant of antioxidant system development during embryogenesis and in early postnatal development. Vitamin E, carotenoids and minerals such as selenium are transferred from feed into egg yolk and further to embryonic tissues (Surai, 1999b). Our data indicate that increased supplementation of the maternal diet can substantially increase vitamin E, carotenoid and Se concentrations in developing chick tissues and significantly decrease susceptibility to lipid peroxidation (Surai, 2000; Surai et al., 1999b; Surai and Speake, 1998a). Postnatal development of the chick is associated with changes in the antioxidant defence strategy (Surai, 1999a,b, 2002a,b,c, 2006, 2012a,b; Surai et al., 2001b; Surai and Fisinin, 2014). The main protection from oxidation in newly hatched chicks is afforded through high concentrations of natural antioxidants, mainly vitamin E and in some cases (wild birds) carotenoids in tissues (Surai, 2002a, Table 1). However, during the first 10 days post-hatch, vitamin E and carotenoid concentrations in the chicken liver decreased 20-fold and the same is true for turkeys, ducks and geese (Surai et al., 1998). To compensate for this decrease, activity of GSH-Px in the liver significantly increases (Surai, 1999a). As a result, this Se-dependent enzyme becomes the major player in antioxidant defence during postnatal development of the chicken.
3.1. Vitamin E in egg yolk and embryo
Vitamin E is considered to be the main antioxidant in egg yolk, which is delivered from the feed and transferred to the developing embryo (Surai, 1999b). Vitamin E is main chain-breaking antioxidant located in biological membranes and lipid droplets. It exists in nature in 8 different forms including 4 tocopherols and 4 tocotrienols with alpha-tocopherol possessing the highest biological activity. Vitamin E is absorbed in the small intestine with efficiency depending on diet composition, level of supplementation, age, sex and other individual characteristics (Surai, 2002a). It is accumulated, to some extent, in the liver and adipose tissues, however, vitamin E reserves in the body are not sufficient to meet long-term requirements (Surai, 1999b). Crude plant oils are the richest source of vitamin E, however, oil refining decreases vitamin E concentration dramatically. In poultry industry vitamin E concentration in feed ingredients is not counted and supplemental vitamin E is the main source of the vitamin for poultry. The recommended level of vitamin E supplementation of breeders (Cobb and Ross) is 100 ppm. Increased (up to 250 ppm) vitamin E supplementation of laying hens in stress conditions is shown to decrease detrimental consequences of heat stress (Bollengier-Lee et al., 1998, 1999; Whitehead et al., 1998). Alpha-tocopherol is unstable and easily oxidised. Vitamin E commercial preparations usually contain esterified forms of the vitamin (e.g., tocopheryl acetate), which are comparatively stable during storage but do not possess antioxidant activity themselves. Only after digestion in the intestine is vitamin E converted to active alpha-tocopherol form (Surai, 2002a). Vitamin E is non-toxic and even very high doses are not associated with hypervitaminosis (Surai et al., 1995) but it is comparatively expensive.
Vitamin E is effectively transferred to the egg yolk and further to the developing embryo. With a single egg a laying hen releasing more vitamin E than all its reserves in the liver (Surai, 1999b). The vitamin E present in the initial yolk consists of a-tocopherol (90%), (b + γ)-tocopherol (8%), a-tocotrienol (0.3%) and (b + γ)-tocotrienol (1.3%) (Surai and Speake, 1998b) and the liver of the embryo may selectively excrete tocotrienols as components of bile, whilst retaining the tocopherols within the hepatocytes. There was discrimination between tocopherols and tocotrienols during their assimilation from the diet by the parent hen and during metabolism by the developing turkey embryo (Surai et al., 1999c). In accordance with vitamin E concentration the tissues of laying hen can be placed in the following descending order: internal fat » liver = heart > lung = pancreas > spleen > muscle > testes > kidney » cerebrum > cerebellum (Surai and Sparks, 2000). Eggs from wild birds are characterised by comparatively high levels of vitamin E concentration (Surai et al., 2001b; Surai, 2002a). For example, vitamin E concentration in the egg yolk of feral gees was shown to be 5-fold higher than that in housed commercial geese (Speake et al., 1999). Similarly, very high vitamin E concentrations were found in the egg yolk of wild gannets, pelicans, cormorants (Surai et al., 2001a), falcons (Barton et al., 2002), penguins, gold- finches, red bishops and house sparrows (Garamszegi et al., 2007). There are species-specific differences in vitamin E accumulation in avian eggs with the chicken egg to be richer in this vitamin in comparison to turkey, goose or duck eggs obtained from breeders supplemented with the same amount of vitamin E (Surai et al., 1998). It is interesting to note that wild gulls invest more vitamin E into the first laid egg in comparison to the third egg in the clutch (Royle et al., 1999). The highest vitamin E concentration is found at time of hatching and vitamin E accumulation in the embryonic liver is considered to be an adaptive mechanism providing antioxidant defence at critical time of hatching (Surai et al., 1996). For the first 2 weeks post-hatch there is a dramatic (10 to 20-fold) reduction in vitamin E concentration in the chicken liver (Surai et al., 1998). Vitamin E is effectively transferred from the egg yolk to the developing embryonic liver and its concentration in the egg determines vitamin E status of the chick at least for the first week post-hatch (Surai et al., 1997). Newly hatched chicks are not able to assimilate vitamin E from the diet effectively and are dependent on its reserved built during embryonic development (Surai, 2002a). Increased vitamin E dietary supplementation of the newly hatched chick has limited ability to increase their vitamin E status. It seems likely that providing vitamin E with water during first week posthatch could be an effective way to maintain their vitamin E status (Surai, 2002a). Indeed, when D-a-tocopherol was added in the drinking water, there was an increase of a-tocopherol in tissues and a decreased susceptibility of red blood cells to haemolysis (SotoSalanova, 1998). Moreover, day-old chickens were treated with 3.25 mg vitamin E/(bird·day) per os, via the drinking water, for 2 weeks. The vitamin E content of both the liver and the blood plasma was significantly higher in the treated chickens than in the untreated controls (Mezes, 1994). Indeed, provision of vitamin E (together with other nutrients, as a part of an antistress composition) with water at time of chicken placement was shown to be effective in commercial conditions of various countries (Shatskih et al., 2015) solving the problem of low vitamin E availability for newly hatched chicks.
Increased dietary vitamin E supplementation of the maternal diet was associated with increased vitamin E concentrations in the egg yolk, embryonic tissues and their increased resistance to oxidative stress (Surai et al., 1999b). There was no influence of dietary vitamin E levels on egg production, but mature breeders (47 weeks old) fed zinc-L-selenomethionine and 120 mg vitamin E/kg feed produced heavier eggs and albumen. Furthermore, hatchability of the eggs of breeders fed 120 mg vitamin E/kg feed was higher than breeders fed 30 mg vitamin at 29 weeks (Urso et al., 2015).
3.2. Carotenoids in egg yolk and embryo
Natural carotenoids have been included into the category of antioxidants (Surai, 2002a, 2012a,b). They could be characterised as follows. Carotenoids are pigments responsible for yellow, orange and sometimes red pigmentation in plants, insects, birds and marine animals. Chicken egg yolk pigmentation is due to presence of carotenoids which are working in the body as an integral part of the antioxidant system, providing recycling other antioxidants including vitamin E and affecting various transcription factors including Nrf2 and NF-kB. Carotenoids are easily accumulating in the egg yolk and transferring to the developing embryo (Surai et al., 2001b,c; Surai, 2012a,b) and the deposition of yolk carotenoids is linked to the process of yolk formation (Müller et al., 2012). It is interesting to note that high carotenoid concentrations were present in species with large body, brain and egg sizes, high absolute metabolic rate and a resident lifestyle (Galvan et al., 2012 ).
Dietary carotenoids determine carotenoid concentration in the egg yolk (Surai et al., 2000). In our study, eggs from carotenoidsupplemented hens contained up to 22 times more carotenoids than controls (Karadas et al., 2005a). The concentration of carotenoids in the livers of chicks hatched from the enriched eggs was initially 29 times greater than in the control chicks. Hepatic carotenoid concentrations in chicks from enriched eggs maintained on the control diet posthatch were sustained at higher values compared with chicks from control eggs that were fed the carotenoid-supplemented diet posthatch, for at least the first 7 days (Karadas et al., 2005a). Therefore, during embryonic development part of accumulated in egg yolk carotenoids are used for metabolic needs including antioxidant defences (Surai, 2002a). Many avian species in wild are characterised by comparatively high (5 to 10-times higher than commercial eggs) carotenoid concentrations (Speake et al., 1999; Surai et al., 2001a; Surai, 2002a; Ewen et al., 2006; Garamszegi et al., 2007). Depending on carotenoid concentration in the livers various species could be placed in the following descending order: free living pheasant > free-range guinea fowl > free-range hen > intensively housed hen > wild mallard > housed duck > free range duck (Karadas et al., 2005b). Eggs obtained from breeders fed a maize-based diet contained at least double the carotenoid concentration in comparison to eggs from laying hens fed a wheat-barley diet (Surai and Sparks, 2001). It was shown that lutein and zeaxanthin are major carotenoids in eggs and embryonic tissues of commercially-reared chickens. Furthermore, in comparison with the control group, there was an increase in the yolk concentrations of lutein, zeaxanthin, lycopene and b-carotene in eggs produced by female quail fed diets supplemented with natural carotenoids (Karadas et al., 2006b). Interestingly, lycopene was shown also to be transferred from the feed to the egg yolk and further to the liver of the developing embryo. Elevated carotenoid concentration in the egg yolk and correspondingly in the liver of newly hatched quail remains significant during the first week posthatch (Karadas et al., 2006a). It has been shown that retinyl oleate and retinyl palmitate concentrations in egg yolk and the liver of day old quail chick significantly increased as a result of carotenoid supplementation of the maternal diet (Karadas et al., 2005a).
Carotenoids have some health-promoting properties, including immune system modulation (Moller et al., 2000). For example, lutein exposure in the diet reduces parameters of inflammation in the liver and spleen of laying-type chicks (Meriwether et al., 2010). It was proven that carotenoid-dependent bright colouration of plumage in males in various avian species in wild is indicative of their health, in particular effective immunity (Blount et al., 2003). There are no established carotenoid requirements for animals. Many plant-derived foods are rich in carotenoids, but animalderived foods are poor carotenoid sources. Carotenoids reserves in the body are limited (Surai, 2002a). During embryonic development carotenoids are actively transferred from egg yolk to the developing embryo with maximal concentration in the chick liver at hatching time (Surai et al., 1996). Similarly to vitamin E, this accumulation is considered to be an adaptive mechanism to deal with hatch-related stress conditions and after hatching carotenoid concentration in the chick liver dramatically decreases (Surai et al., 1998). Chicks obtained from carotenoid-depleted eggs are not able to assimilate carotenoids from the diet to the same extent as those hatched from carotenoid-rich eggs (Koutsos et al., 2003). Increased dietary carotenoid supplementation of the maternal diet was associated with increased vitamin E concentrations in the egg yolk, embryonic tissues and their increased resistance to oxidative stress (Surai and Speake, 1998a; Surai et al., 2003). A combination of increased concentrations of lycopene and lutein in the egg yolk results in elevated concentrations of CoQ in the liver of the newly hatched quail (Karadas et al., 2006a). Increased lutein concentration in egg yolk was shown to have positive effect on offspring survival in the great tit (Marri and Richner, 2014). Furthermore, there is a direct positive relationship between enhanced maternal investment of carotenoids and the ability to cope with a specific and costly parasite in young birds (Ewen et al., 2009). In fact, maternal effects mediated by carotenoids may act as a proxy factor in development and phenotypic plasticity in traits associated with nestling fitness, such as immune response and ability to metabolise and use antioxidants, and ultimately participate in the evolution of phenotypic traits (Biard et al., 2007). It seems likely that carotenoid availability can modulate the trade-off between reproduction and resistance to oxidative stress in zebra finches (Bertrand et al., 2006). In fact, maternally derived carotenoid pigments affect offspring survival, sex ratio, and sexual attractiveness in a colourful songbird (McGraw et al., 2005). In the blue tit, nestlings from eggs laid by carotenoid supplemented females had longer tarsi, had faster development of the immune system and grew brighter yellow feathers than nestlings from control females (Biard et al., 2005). It was suggested that since carotenoid availability in wild birds is limiting during egg laying, females might have to balance the benefits of investing in egg quality against the potential costs of impairing their own future antioxidant protection. It seems likely that both sexual display and immune defences can be limited by carotenoid availability (Blount, 2004), providing support for the hypothesis that males allocating greater amounts of carotenoids to sexual colouration are advertising their superior health (Blount et al., 2003). Carotenoid availability has also been shown to limit egg-laying capacity in gulls (Blount, 2004; Blount et al., 2004). In fact, maternal yolk carotenoids can have a major effect in promoting a fundamental component of immunity that predicts offspring survival and suggests that adaptive early maternal effects can be mediated by transmission of antioxidants to egg (Saino et al., 2003). In red-legged partridges, plasma carotenoids at the end of laying were strongly correlated with the number of eggs laid (Bortolotti et al., 2003). Maternal carotenoid status modifies the incorporation of dietary carotenoids into immune tissues of growing chickens (Gallus gallus domesticus) (Koutsos et al., 2003). In a supplemental feeding study of lesser black-backed gulls, Larus fuscus, carotenoid-fed females were shown to have increased integument pigmentation, elevated plasma concentrations of carotenoids and enhanced antioxidant activity (Blount et al., 2002). It is concluded that the maternal diet plays an important role in antioxidant systems formation during chick embryonic development; the maizebased diet can increase the antioxidant potential of the egg yolk and embryonic tissues compared to the antioxidant potential provided by parent birds fed the wheat-based diet (Surai and Sparks, 2001).
Our results indicate that avian embryos discriminate between different carotenoids during their distribution from the yolk to the various tissues (Surai et al., 2001d). In contrast to data for eggs of the domestic chicken, b-carotene was a significant component in the yolks of three wild species (the common moorhen [Gallinula chloropus], American coot [Fulica americana] and lesser blackbacked gull [L. fuscus]), forming 25% to 29% by weight of the total carotenoids present. The concentration of total carotenoids in the livers of the newly-hatched chicks was 5 to 10 times higher than in the other tissues and b-carotene was again a major component, forming 37% to 58% of the hepatic carotenoids. In the newlyhatched gull, the proportions of both lutein and zeaxanthin were very low in the liver but high in the heart and muscle when compared with the yolk. By contrast, canthaxanthin (CX), echinenone and b-carotene were very minor constituents of heart and muscle when compared with their proportions in the yolk of the gull. The proportions of lutein and zeaxanthin in the liver of the newly-hatched coot and moorhen were also far lower than in the yolk whereas the liver was relatively enriched with b-cryptoxanthin, b-carotene and (in the moorhen) echinenone (Surai et al., 2001d).
It was proposed that maternal investment in yolk carotenoids promotes the evolution of carotenoid-based ornaments and proven that maternal effects mediated by yolk carotenoids play a role in the evolution of carotenoid-based signals as a response to sexual selection, likely based on organisational effects of carotenoids during embryo development (Biard et al., 2009). Across species, there were 6 P.F. Surai et al. / Animal Nutrition 2 (2016) 1e11 positive relationships between embryonic growth rate and total yolk levels of carotenoids or vitamin E. Moreover, there were consistent differences in antioxidant provision between pairs of species that share a common initial egg mass yet have differing rates of embryonic growth, such that the eggs of the fasterdeveloping species have higher levels of carotenoids and vitamin E (Deeming and Pike, 2013).
Among more than 750 known carotenoids, CX has a special place as a carotenoid with proven antioxidant and other biologically-relevant functions. A great body of evidence indicates that CX possesses high antioxidant activity that was shown in various in vitro model systems as well as in animal experiments in vivo. Recently effects of CX in the breeder's diet have been studied (Surai et al., 2003). The control group received the control diet containing <2 mg/kg total xanthophylls. For treatments 2 to 5 the control diet was supplemented with 3, 6, 12 and 24 mg/kg CX, respectively. The control diet did not contain CX. Inclusion of CX into the maternal diet caused a significant dose-dependent response in terms of its accumulation in the egg yolk. Egg yolk colour gradually changed from pale yellow in the control group to dark red in the laying hens fed CX at 24 mg/kg. The most important finding of this study was a positive effect of CX on vitamin E in the developing chicks. As a result of enrichment of the chick liver with CX and vitamin E due to the maternal dietary supplementation, tissue susceptibility to lipid peroxidation significantly decreased. This was the case in 1-day-old and 7-day-old chicks. In a later study it was shown that maternal diet is the main determinant of the progeny's carotenoid status for at least 7 days after hatching (Karadas et al., 2005a). The enhancement of the antioxidant system of the developing chick as a result of feed additives to maternal diet presents great opportunities for poultry producers. Postnatal development of the chicken is a crucial time for the maturation of major physiological systems, including the immune system, as well as a time of high risk of peroxidation (Surai, 2002a). Therefore, increased supplementation of the maternal diet with carotenoids, in particular CX, could help maintain antioxidant system efficiency and high chick viability.
The idea of beneficial effect of dietary CX supplementation of the breeder diet was further developed by Zhang et al. (2011). In this study, the breeder hens were fed either a basal diet or the basal diet supplemented with 6 mg of CX per kg for 24 weeks. Canthaxanthin supplementation resulted in a significant increase in the yolk colourimetric score of DSM Yolk Colour Fan, con- firming an effective CX transfer from the diet to the egg yolk. Egg yolk enrichment with CX was associated with a significant improvement of the antioxidant status of the egg yolk. Indeed, malondialdehyde (MDA) content of the egg yolk significantly decreased, while total antioxidant capacity (TAC) of the egg yolk significantly increased. This could be an important element in an improvement of protection of laying birds from commerciallyrelevant stresses (Surai, 2002a). The antioxidant capacity of the newly hatched chicks was significantly increased due to maternal CX supplementation. There was a significant decrease in MDA in plasma of day-old chicks. It is of great importance to see an increased SOD activity in the chicken plasma. It is interesting to underline that maternal supplementation with CX has also positive effect on TAC of newly hatched chicken which increased by 33%. In a field trial, Robert et al. (2008) studied the effect of CX in ROSS breeders on the anti-oxidant status of their progeny. They observed that the antioxidant status of sera of 1-day-old chicks was significantly higher and the thiobarbituric acid reactive substance (TBARS) level significantly lower with 6 ppm CX in the breeder feed. The experimental chicks also showed a lower mortality (0 vs. 4%) during the first 21 days posthatching. These findings support the hypothesis that CX supplementation of the maternal diet enhances the protective capacity of tissues against oxidative stress in vivo, which might be beneficial for poultry producers. The next step in proving a beneficial effect of CX supplementation of the maternal diet was an experiment conducted in Brazil (Rosa et al., 2012). In terms of breeder performance, CX was observed to significantly improve egg laying rate in certain periods, but overall there was no difference between the two treatment groups. Similarly to previous observations the reduction of TBARS was observed in yolks from stored hatching eggs produced by breeders fed diets plus CX. This reduction was observed in eggs submitted to analysis on the same day they were produced and in eggs stored for 4, 8 and 12 days. The most important finding of this study was a positive effect of CX supplementation of the maternal diet on fertility, hatchability and embryonic mortality. In fact, CX in comparison to the control improved fertility and hatchability and reduced embryonic mortality. As a result, hatching rate was significantly improved. As to the different stages of embryo mortality, CX was most efficacious for its prevention in the first 48 h as well as between days 15 and 21 of incubation. In the study, CX supplementation improved breeder fertility perhaps by improving survival and storage of spermatozoa within the reproductive tract of the hen. It is interesting to mention that another carotenoid with a similar structure astaxanthin fed to breeders was shown to be transferred to the egg yolk and had a significant protective effect against decreasing hatchability due to egg storage at 21 °C (Saito and Kita, 2011). In the study the hatchability of eggs stored at 21 °C was significantly lower than that at 10 °C, and lowered hatchability was restored by maternal intake of astaxanthin. Elevated levels of dietary astaxanthin from 5 to 20 ppm gradually restrained the decrease in hatchability at 21 °C.
It is proven that, nonprovitamin A carotenoids (lutein, CX, astaxanthin and lycopene) are important regulators of cellmediated and humoural immune response in animals and humans (Moller et al., 2000; Chew and Park, 2004). Recent studies on the role of carotenoids in gene regulation and apoptosis have substantially advanced our knowledge on the possible mechanism by which carotenoids regulate immune functions. For example, a number of differentially expressed genes in chicken muscles due to carotenoid dietary supplementation at 100 mg/kg have been identified (Tarique et al., 2014). In particular, in CX and lutein supplemented chickens there were 54 (32 upregulated and 22 downregulated) and 23 (15 upregulated and 8 downregulated) altered known genes, respectively. Indeed, the differently expressed genes are shown to be involved in important biological processes, including pigmentation, growth, molecular mechanisms, fat metabolism, cell proliferation, immune response, lipid metabolism, and protein synthesis and degradation (Tarique et al., 2014).
As mentioned above, immunomodulating properties of carotenoids in wild birds have receive substantial attention. However, commercial poultry production is still waiting for some important answers related to the link between dietary carotenoids and immunocompetence in commercial birds. There is a possibility that a flock with certain types of pigments in their diet could be more resilient or use less energy towards their immune system compared with a flock without carotenoid supplementation. However, immunomodulating properties of carotenoids would depend on many different factors, including efficacy of carotenoid assimilation from the diet and their accumulation in target tissues; diet composition and concentrations of other antioxidants (e.g., vitamin E, Se, ascorbic acid) and prooxidants (e.g., polyunsaturated fatty acids) in the diet; various stresses, including specific immune challenges; age of birds and other factors (Surai, 2002a; Surai and Fisinin, 2012; Fisinin and Surai, 2013a,b; Shatskih et al., 2015). Indeed, more research is needed to make commercial P.F. Surai et al. / Animal Nutrition 2 (2016) 1e11 7 recommendations for the use of natural pigments for the purpose of immunity, but the aforementioned results are quite promising.
3.3. Selenium in egg yolk and embryo
Selenium is an essential part of a range of selenoproteins, including GSH-Px, thioredoxin reductase (TrxR), iodothyronine deiodinase (ID) and some others. In the animal and human body 25 selenoproteins have been identified to date (Surai, 2006). Food and feed ingredients contain variable concentrations of Se, but most of them are low in this element. Physiological requirement in Se is low, but if not met, antioxidant systems are compromised with detrimental consequences for animal/poultry health. In high doses Se is toxic. However, the doses of dietary Se which could be detrimental are at least 10-fold higher in comparison to those commercially used. There are two major sources of supplemental Se for poultry: a natural source in the form of various seleno-amino acids including selenomethionine (Se-Met) or inorganic selenium in the form of selenite or selenate (SS, Surai and Fisinin, 2014). It is proven that organic selenium supplementation has physiological and biochemical benefits in animals, including poultry (Surai, 2006; Surai and Fisinin, 2014, 2015b,c). Selenium concentration in the egg yolk and egg white depends on its concentration and form in the diet (Surai, 2000). The main form of selenium in the egg is SeMet, which cannot be synthesised by laying hens (Lipiec et al., 2010). Therefore, inclusion of organic selenium in the chicken diet is associated with a substantial increase Se concentration in the embryonic tissues (Surai, 2000; Yuan et al., 2011). Increased Se concentrations in the egg yolk and albumin are related to increase Se levels in the embryonic liver and increased antioxidant defences of the developing chicks (Surai, 2000). The order of Se distribution was liver > kidney > spleen > cardiac muscle > egg > blood > breast muscle, irrespective of the addition level or source (Pan et al., 2007). Dietary organic Se supplementation was shown to increase antioxidant defences in the tissues of the developing embryos (Surai, 2000). It was shown that the Se retention efficiency of SeMet is higher than that of SY or SS for broiler breeders and their offspring (Yuan et al., 2011). In fact, the embryonic liver and breast muscle from Se-Met treatments had higher Se concentrations than those of sodium selenite treatments (Yuan et al., 2011). The Se concentrations in liver, kidney, and breast muscle of 1-day-old chicks were greater in Se-yeast or Se-Met treatments compared with SS treatments. Furthermore, Se-Met was shown to be more effective than sodium selenite in maintaining antioxidant protection of the developing chick embryo (Xiao et al., 2015). Indeed, within the organic Se sources, L-selenomethionine showed higher Se transfer to eggs than Se-enriched yeast (Delezie et al., 2014; Jing et al., 2015). However, there is some concern about SeMet stability in the commercial premixes (Surai and Fisinin, 2014, 2015b, 2016) and it seems likely that a new stable organic Se source in the form of 2-hydroxy-4-methylselenobutanoic acid (HMSeBA) is going to be a major Se source in poultry and pig production. Indeed, hens fed the diet with HMSeBA at 0.2 ppm accumulated more Se in their eggs (+28.78%; P < 0.01) and muscles (+28%; P < 0.01) than those fed the diet supplemented with Se-yeast at the same level of supplementation (Jlali et al., 2013). In an experiment with the growing chickens it was shown that after 21 days, organic Se sources maintained (Se-yeast) or increased (HMSeBA) (P < 0.05) breast muscle Se concentration compared to hatch value whereas inorganic source (SS) or non-supplemented group (NC) showed a significant decrease in tissue Se concentration (P < 0.05) (Couloigner et al., 2015). Maternal Se has a long-term consequences for the developing embryo and newly hatched chicks (Pappas et al., 2005). Maternal organic selenium and polyunsaturated fatty acids were shown to affect progeny performance and levels of selenium and docosahexaenoic acid in the chick tissues (Pappas et al., 2006a). It is interesting to note that fish oil inclusion in the breeder diet increased embryonic mortality at week 3 of incubation and reduced both hatchability and 1-day-old chick weight in hens of both ages (Pappas et al., 2006b). The addition of Se to the fish oil-containing diets ameliorated some of these adverse effects, because chicks hatched from eggs laid by 23-week-old breeders of the fish oil + Se treatment were heavier than those receiving the fish oil treatment (Pappas et al., 2006b). It seems likely that Se accumulated in the egg can affect gene expression of the developing embryo and this could result in better antioxidant defences and reduced Se use during development or alternatively specific enzymatic systems involved in the Se metabolism could be affected and Se would be more effectively assimilated from the diet. The dietary inclusion of organic selenium in the form of zinc-L-selenomethionine promoted heavier hatchling weight until egg production peak (33 weeks), but did not influence hatchling quality or hatching window (Urso et al., 2015).
3.4. Other antioxidants
Since ascorbic acid is synthesised in the chicken body and not found in the egg its direct role in internal egg quality is questionable. The results of Creel et al. (2001) did not provide evidence of a beneficial reproductive response to the inclusion of ascorbic acid in commercial broiler breeder diets. However, in stress conditions ascorbic acid supplementation of the breeder's diet could have some positive effects (Shit et al., 2012; Wang et al., 2011; Cheng et al., 1990). It is interesting to mention that, L-carnitine, possessing antioxidant properties (Surai, 2015a,d,e), in the diet of hens affected carcase traits of their progeny. In particular, dietary hen Lcarnitine decreased carcase fat and increased breast meat in progeny fed high nutrient density diets (Kidd et al., 2005). Furthermore, dietary L-carnitine, as compared with the control diet, increased egg yolk L-carnitine concentration, decreased hatchling yolk sac weights, decreased yolk sac lipid content at hatch, and culminated in compositional changes of yolk fatty acids (Zhai et al., 2008).
There were several attempts to improve hatching egg quality by inclusion into the breeder's diet various plant extracts, essential oils and some other compounds, however, it is too early to make any conclusion about their efficacy. Indeed, low polyphenol absorption and tissue levels (Surai, 2014) indicate that the mechanisms of their biological activities are far beyond antioxidant activities. Among various plant extracts widely used in human medicine silymarin has a special place being a natural hepatoprotector in various stress conditions (Madrigal-Santillan et al., 2014; Casas-Grajales and Muriel, 2015). However, research data on the role of silymarin in poultry nutrition are quite limited (Schiavone et al., 2007; Khatoon et al., 2013). It seems likely that silymarin will receive substantial attention in future due to its unique properties and antioxidant action (Surai, 2015c). Furthermore, taking into account antioxidant properties of taurine (Shimada et al., 2015) and its effects on transcription factors (NF-kB and Nrf2, Agca et al., 2014) one can expect its positive effect of egg production and quality, including antioxidant defences of the developing chick embryo. In addition, roles of vitamin D in antioxidant defence system regulation (Bhat and Ismail, 2015) and cell signalling (Berridge, 2015) warrant reconsideration of its role in chick embryo development and antioxidant defence.
4. Conclusions
Natural antioxidants play important roles in poultry production. In the body all antioxidants work together to create the antioxidant 8 P.F. Surai et al. / Animal Nutrition 2 (2016) 1e11 system. There is a delicate balance between the amount of free radicals generated in the body and the antioxidants to protect against them and the disruption of such a balance causes an oxidative stress. In breeder nutrition, vitamin E, carotenoids and selenium received most of attention as a way of improving stress resistance of poultry. Furthermore, adaptive effects of other natural antioxidants, such as carnitine, taurine and silymarin are a subject for future research.
This article was originally published in Animal Nutrition, Volume 2, Issue 1, March 2016, Pages 1-11. https://doi.org/10.1016/j.aninu.2016.01.001. This is an Open Access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
References
Agca CA, Tuzcu M, Hayirli A, Sahin K. Taurine ameliorates neuropathy via regulating NF-kB and Nrf2/HO-1 signaling cascades in diabetic rats. Food Chem Toxicol 2014;71:116e21.
Ames BN. An enthusiasm for metabolism. J Biol Chem 2003;278:4369e80.
Ames BN, Gold LS. The causes and prevention of cancer: gaining perspective. Environ Health Perspect 1997;105(Suppl. 4):865e73.
Barton NWH, Fox NC, Surai PF, Speake BK. Vitamins E and A, carotenoids and fatty acids of the egg yolk of raptors. J Raptor Res 2002;36:33e8.
Berridge MJ. Vitamin D cell signalling in health and disease. Biochem Biophys Res Commun 2015;460:53e71.
Bertrand S, Alonso-Alvarez C, Devevey G, Faivre B, Prost J, Sorci G. Carotenoids modulate the trade-off between egg production and resistance to oxidative stress in zebra finches. Oecologia 2006;147:576e84.
Bhat M, Ismail A. Vitamin D treatment protects against and reverses oxidative stress induced muscle proteolysis. J Steroid Biochem Mol Biol 2015;152:171e9.
Biard C, Surai PF, Møller AP. Effects of carotenoid availability during laying on reproduction in the blue tit. Oecologia 2005;144:32e44.
Biard C, Surai PF, Møller AP. An analysis of pre- and post-hatching maternal effects mediated by carotenoids in the blue tit. J Evol Biol 2007;20:326e39.
Biard C, Gil D, Karadas¸ F, Saino N, Spottiswoode CN, Surai PF, et al. Maternal effects mediated by antioxidants and the evolution of carotenoid-based signals in birds. Am Nat 2009;174:696e708.
Blount JD. Carotenoids and life-history evolution in animals. Arch Biochem Biophys 2004;430:10e5.
Blount JD, Surai PF, Nager RG, Houston DC, Møller AP, Trewby ML, et al. Carotenoids and egg quality in the lesser blackbacked gull Larus fuscus: a supplemental feeding study of maternal effects. Proc Biol Sci 2002;269:29e36.
Blount JD, Houston DC, Surai PF, Møller AP. Egg-laying capacity is limited by carotenoid pigment availability in wild gulls Larus fuscus. Proc Biol Sci 2004;271(Suppl. 3):S79e81.
Blount JD, Metcalfe NB, Birkhead TR, Surai PF. Carotenoid modulation of immune function and sexual attractiveness in zebra finches. Science 2003;300:125e7.
Bollengier-Lee S, Williams PE, Whitehead CC. Optimal dietary concentration of vitamin E for alleviating the effect of heat stress on egg production in laying hens. Br Poult Sci 1999;40:102e7.
Bollengier-Lee S, Mitchell MA, Utomo DB, Williams PE, Whitehead CC. Influence of high dietary vitamin E supplementation on egg production and plasma characteristics in hens subjected to heat stress. Br Poult Sci 1998;39:106e12.
Bortolotti GR, Negro JJ, Surai PF, Prieto P. Carotenoids in eggs and plasma of redlegged partridges: effects of diet and reproductive output. Physiol Biochem Zool 2003;76:367e74.
Casas-Grajales S, Muriel P. Antioxidants in liver health. World J Gastrointest Pharmacol Ther 2015;6:59e72.
Chew BP, Park JS. Carotenoid action on the immune response. J Nutr 2004;134: 257Se61S.
Chance B, Sies H, Boveries A. Hydroperoxide metabolism in mammalian organs. Physiol Rev 1979;59:527e605.
Cheng TK, Coon CN, Hamre ML. Effect of environmental stress on the ascorbic acid requirement of laying hens. Poult Sci 1990;69:774e80.
Couloigner F, Jlali M, Briens M, Rouffineau F, Geraert PA, Mercier Y. Selenium deposition kinetics of different selenium sources in muscle and feathers of broilers. Poult Sci 2015;94:2708e14.
Creel LH, Maurice DV, Lightsey SF, Grimes LW. Stability of dietary ascorbic acid and the effect of supplementation on reproductive performance of broiler breeder chickens. Br Poult Sci 2001;42:96e101.
Dallner G, Sindelar PJ. Regulation of ubiquinone metabolism. Free Radic Biol Med 2000;29:285e94.
Decuipere E, Tona K, Bruggeman V, Bamelis F. The day-old chick: a crucial hinge between breeders and broilers. World's Poult Sci J 2001;57:127e38.
Deeming DC, Pike TW. Embryonic growth and antioxidant provision in avian eggs. Biol Lett 2013;9(6). 20130757.
Delezie E, Rovers M, Van der Aa A, Ruttens A, Wittocx S, Segers L. Comparing responses to different selenium sources and dosages in laying hens. Poult Sci 2014;93:3083e90.
Enkvetchakul B, Anthony NB, Bottje WG. Liver and blood glutathione in male broiler chickens, turkeys, and quail. Poult Sci 1995;74:885e9.
Ewen JG, Thorogood R, Karadas F, Pappas AC, Surai PF. Influences of carotenoid supplementation on the integrated antioxidant system of a free living endangered passerine, the hihi (Notiomystis cincta). Comp Biochem Physiol A Mol Integr Physiol 2006;143:149e54.
Ewen JG, Thorogood R, Brekke P, Cassey P, Karadas F, Armstrong DP. Maternally invested carotenoids compensate costly ectoparasitism in the hihi. Proc Natl Acad Sci U S A 2009;106:12798e802.
Fisinin VI, Surai PF. Immunity in modern animal and poultry production: from theory to practical aspects of immunomodulation. Russ Poult Sci Ptitsevodstvo, Russ 2013a;5:4e10.
Fisinin VI, Surai PF. Gut immunity in birds: facts and thinking. Agric Biol Selskokhozaistvennaya Biol Russ 2013b;4:1e25.
Gaal T, Mezes M, Noble RC, Dixon J, Speake BK. Development of antioxidant capacity in tissues of the chick embryo. Comp Biochem Physiol B 1995;112:711e6.
Galvan I, Erritzøe J, Karadas ¸ F, Møller AP. High levels of liver antioxidants are associated with life-history strategies characteristic of slow growth and high survival rates in birds. J Comp Physiol B 2012;182:947e59.
Garamszegi LZ, Biard C, Eens M, Møller AP, Saino N, Surai P. Maternal effects and the evolution of brain size in birds: overlooked developmental constraints. Neurosci Biobehav Rev 2007;31:498e515.
Halliwell B. Free radicals and antioxidants: a personal view. Nutr Rev 1994;52: 253e65.
Helbock HJ, Beckman KB, Shigenaga MK, Walter PB, Woodall AA, Yeo HC, et al. DNA oxidation matters: the HPLC-electrochemical detection assay of 8-oxo-deoxyguanosine and 8-oxo-guanine. Proc Natl Acad Sci U S A 1998;95:288e93.
Hohtola E. Facultative and obligatory thermogenesis in young birds: a cautionary note. Comp Biochem Physiol A Mol Integr Physiol 2002;131:733e9.
Jing CL, Dong XF, Wang ZM, Liu S, Tong JM. Comparative study of DLselenomethionine vs sodium selenite and seleno-yeast on antioxidant activity and selenium status in laying hens. Poult Sci 2015;94:965e75.
Jlali M, Briens M, Rouffineau F, Mercerand F, Geraert PA, Mercier Y. Effect of 2- hydroxy-4-methylselenobutanoic acid as a dietary selenium supplement to improve the selenium concentration of table eggs. J Anim Sci 2013;91:1745e52.
Karadas F, Pappas AC, Surai PF, Speake BK. Embryonic development within carotenoid-enriched eggs influences the post-hatch carotenoid status of the chicken. Comp Biochem Physiol B Biochem Mol Biol 2005a;141:244e51.
Karadas F, Wood NA, Surai PF, Sparks NH. Tissue-specific distribution of carotenoids and vitamin E in tissues of newly hatched chicks from various avian species. Comp Biochem Physiol A Mol Integr Physiol 2005b;140:506e11.
Karadas F, Surai P, Grammenidis E, Sparks NH, Acamovic T. Supplementation of the maternal diet with tomato powder and marigold extract: effects on the antioxidant system of the developing quail. Br Poult Sci 2006a;47:200e8.
Karadas F, Grammenidis E, Surai PF, Acamovic T, Sparks NHC. Effects of carotenoids from lucerne, marigold and tomato on egg yolk pigmentation and carotenoid composition. Br Poult Sci 2006b;47:561e6.
Karadas F, Surai PF, Sparks NH. Changes in broiler chick tissue concentrations of lipid-soluble antioxidants immediately post-hatch. Comp Biochem Physiol A Mol Integr Physiol 2011;160:68e71.
Kettle AJ, Winterbourn CC. Myeloperoxidase: a key regulator of neutrophil oxidant production. Redox Rep 1997;3:3e15.
Khatoon A, Zargham Khan M, Khan A, Saleemi MK, Javed I. Amelioration of ochratoxin A-induced immunotoxic effects by silymarin and vitamin E in white leghorn cockerels. J Immunotoxicol 2013;10:25e31.
Kidd MT, McDaniel CD, Peebles ED, Barber SJ, Corzo A, Branton SL, et al. Breeder hen dietary L-carnitine affects progeny carcase traits. Br Poult Sci 2005;46:97e103.
Koutsos EA, Clifford AJ, Calvert CC, Klasing KC. Maternal carotenoid status modifies the incorporation of dietary carotenoids into immune tissues of growing chickens (Gallus gallus domesticus). J Nutr 2003;133:1132e8.
Lipiec E, Siara G, Bierla K, Ouerdane L, Szpunar J. Determination of selenomethionine, selenocysteine, and inorganic selenium in eggs by HPLC-inductively coupled plasma mass spectrometry. Anal Bioanal Chem 2010;397:731e41.
Madrigal-Santillan E, Madrigal-Bujaidar E, Alvarez-Gonz alez I, Sumaya- Martínez MT, Gutierrez-Salinas J, Bautista M, Morales-Gonz alez A, García-Luna, Gonz alez-Rubio M, Aguilar-Faisal JL, Morales-Gonz alez JA. Review of natural products with hepatoprotective effects. World J Gastroenterol 2014;20: 14787e804.
Marri V, Richner H. Yolk carotenoids increase fledging success in great tit nestlings. Oecologia 2014;176:371e7.
Meriwether LS, Humphrey BD, Peterson DG, Klasing KC, Koutsos EA. Lutein exposure, in ovo or in the diet, reduces parameters of inflammation in the liver and spleen laying-type chicks (Gallus gallus domesticus). J Anim Physiol Anim Nutr Berl 2010;94:e115e22.
McGraw KJ, Adkins-Regan E, Parker RS. Maternally derived carotenoid pigments affect offspring survival, sex ratio, and sexual attractiveness in a colourful songbird. Naturwissenschaften 2005;92:375e80.
Mezes M. Effect of vitamin E treatment on early postnatal changes of vitamin E status of chicken. Acta Vet Hung 1994;42:477e80.
Moller AP, Biard C, Blount JD, Houston DC, Ninni P, Saino N, et al. Carotenoiddependent signals: indicators of foraging efficiency, immunocompetence or detoxification ability? Poult Avian Biol Rev 2000;11:137e59.
Müller W, Vergauwen J, Eens M, Blount JD. Environmental effects shape the maternal transfer of carotenoids and vitamin E to the yolk. Front Zool 2012;9(1):17.
Pan C, Huang K, Zhao Y, Qin S, Fu Chen, Hu Q. Effect of selenium source and level in hen's diet on tissue selenium deposition and egg selenium concentrations. J Agric Food Chem 2007;55:1027e32.
Pappas AC, Karadas F, Surai PF, Speake BK. The selenium intake of the female chicken influences the selenium status of her progeny. Comp Biochem Physiol B Biochem Mol Biol 2005;142:465e74. P.F.
Pappas AC, Acamovic T, Surai PF, McDevitt RM. Maternal organo-selenium compounds and polyunsaturated fatty acids affect progeny performance and levels of selenium and docosahexaenoic acid in the chick tissues. Poult Sci 2006a;85: 1610e20.
Pappas AC, Acamovic T, Sparks NH, Surai PF, McDevitt RM. Effects of supplementing broiler breeder diets with organoselenium compounds and polyunsaturated fatty acids on hatchability. Poult Sci 2006b;85:1584e93.
Reczek CR, Chandel NS. ROS-dependent signal transduction. Curr Opin Cell Biol 2015;33:8e13.
Robert F, Panheleux-LeBastard M, Hamelin C, Boulard C. Effect of canthaxanthin supplementation in the ROSS breeder diet on oxidative stress in chick. In: Proc. 16th European symposium on poultry nutrition; 2008. p. 731e4. France.
Rosa AP, Scher A, Sorbara JO, Boemo LS, Forgiarini J, Londero A. Effects of canthaxanthin on the productive and reproductive performance of broiler breeders. Poult Sci 2012;91:660e6.
Royle NJ, Surai PF, McCartney RJ, Speake BK. Parental investment and egg yolk lipid composition in gulls. Funct Ecol 1999;13:298e306.
Saino N, Ferrari R, Romano M, Martinelli R, Møller AP. Experimental manipulation of egg carotenoids affects immunity of barn swallow nestlings. Proc Biol Sci 2003;270:2485e9.
Saito F, Kita K. Maternal intake of astaxanthin improved hatchability of fertilized eggs stored at high temperature. J Poult Sci 2011;1:33e9. Shatskih E, Latipova E, Fisinin V, Denev S, Surai PF. Molecular mechanisms and new strategies to fight stresses in egg producing birds. Agric Sci Technol 2015;7: 3e10.
Schiavone A, Righi F, Quarantelli A, Bruni R, Serventi P, Fusari A. Use of Silybum marianum fruit extract in broiler chicken nutrition: influence on performance and meat quality. J Anim Physiol Anim Nutr Berl 2007;91: 256e62.
Shimada K, Jong CJ, Takahashi K, Schaffer SW. Role of ROS production and turnover in the antioxidant activity of taurine. Adv Exp Med Biol 2015;803: 581e96.
Shit N, Singh RP, Sastry KV, Agarwal R, Singh R, Pandey NK, et al. Effect of dietary Lascorbic acid (L-AA) on production performance, egg quality traits and fertility in Japanese quail (Coturnix japonica) at low ambient temperature. Asian-Australas J Anim Sci 2012;25:1009e14.
Siegel PB, Price SE, Meldrum B, Picard M, Gefraert PA. Performance of pureline broiler breeders fed two levels of vitamin E. Poult Sci 2001;80:1258e62.
Singal PK, Khaper N, Palace V, Kumar D. The role of oxidative stress in the genesis of heart disease. Cardiovasc Res 1998;40:426e32.
Soto-Salanova MF. Vitamin E in young turkeys: a reassessment of the requirement [Retrospective theses and dissertations]. Digital Repository @ Iowa State University; 1998.
Speake BK, Murray AMB, Noble RC. Transport and transformation of yolk lipids during development of the avian embryo. Prog Lipid Res 1998;37:1e32.
Speake BK, Surai PF, Noble RC, Beer JV, Wood N. Differences in egg lipid and antioxidant composition between wild and captive pheasants and geese. Comp Biochem Physiol B 1999;124(1):101e7.
Surai PF. Tissue-specific changes in the activities of antioxidant enzymes during the development of the chicken embryo. Br Poult Sci 1999a;40: 397e405.
Surai PF. Vitamin E in avian reproduction. Poult Avian Biol Rev 1999b;10:1e60. Surai PF. Effect of the selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. Br Poult Sci 2000;41: 235e43.
Surai PF. Natural antioxidants in avian nutrition and reproduction. Nottingham, UK: Nottingham University Press; 2002a.
Surai PF. Selenium in poultry nutrition: a new look at an old element. 1. Antioxidant properties, deficiency and toxicity. World's Poult Sci J 2002b;58:333e47.
Surai PF. Selenium in poultry nutrition: a new look at an old element. 2. Reproduction, egg and meat quality and practical applications. World's Poult Sci J 2002c;58:431e50.
Surai PF. Selenium in nutrition and health. Nottingham, UK: Nottingham University Press; 2006.
Surai PF. The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 1. World's Poult Sci J 2012a;68:465e75.
Surai PF. The antioxidant properties of canthaxanthin and its potential effects in the poultry eggs and on embryonic development of the chick. Part 2. World's Poult Sci J 2012b;68:717e26.
Surai PF. Polyphenol compounds in the chicken/animal diet: from the past to the future. J Anim Physiol Anim Nutr 2014;98:19e31.
Surai PF. Antioxidant action of carnitine: molecular mechanisms and practical applications. EC Vet Sci 2015a;2:66e84.
Surai PF. Antioxidant systems in poultry biology: heat shock proteins. J Sci 2015b;5: 1188e222. Surai PF. Silymarin as a natural antioxidant: an overview of the current evidence and perspectives. Antioxidants 2015c;4:204e47.
Surai PF. Carnitine enigma: from antioxidant action to vitagene regulation. Part 1. Absorption, metabolism and antioxidant activities. J Vet Sci Med 2015d;3(2): 14.
Surai PF. Carnitine enigma: from antioxidant action to vitagene regulation. Part 2. Transcription factors and practical applications. J Vet Sci Med 2015e;3(2): 17.
Surai PF, Bortolotti GR, Fidgett A, Blount J, Speake BK. Effects of piscivory on the fatty acid profiles and antioxidants of avian yolk: studies on eggs of the gannet, skua, pelican and cormorant. J Zool Lond 2001a;255:305e12.
Surai PF, Fisinin VI. Selenium in poultry breeder nutrition. An update. Anim Feed Sci Technol 2014;191:1e15.
Surai PF, Fisinin VI. Antioxidanteprooxidant balance in the intestine: applications in chick placement and pig weaning. J Vet Sci Med 2015a;3:1e16.
Surai PF, Fisinin VI. Selenium in sow nutrition. Anim Feed Sci Technol 2016;211: 18e30.
Surai PF, Fisinin VI. Selenium in pig nutrition and reproduction: boars and semen quality e a review. Asian-Australas J Anim Sci 2015b;28:730e46.
Surai PF, Fisinin VI. Modern methods of fighting stresses in poultry production: from antioxidants to vitagenes. Agric Biol Selskokhozaistvennaya Biol Russ 2012;4:3e13.
Surai P, Gaal T, Noble R, Speake B. The relationship between a-tocopherol content of the yolk and its accumulation in the tissues of the newly hatched chick. J Sci Food Agric 1997;75:212e6.
Surai P, Ionov I, Kuchmistova E, Noble RC, Speake BK. The relationship between the levels of a-tocopherol and carotenoids in the maternal feed, yolk and neonatal tissues: comparison between the chicken, turkey, duck and goose. J Sci Food Agric 1998;76:593e8.
Surai P, Ionov I, Buzhin A, Buzhina N. Vitamin E and egg quality. In: Proc. VI European symposium on the quality of egg and egg products, Zaragoza, Spain; 1995. p. 387e94.
Surai PF, MacPherson A, Speake BK, Sparks NH. Designer egg evaluation in a controlled trial. Eur J Clin Nutr 2000;54:298e305.
Surai PF, Noble RC, Speake BK. Relationship between vitamin E content and susceptibility to lipid peroxidation in tissues of the newly hatched chick. Br Poult Sci 1999b;40:406e10.
Surai PF, Noble RC, Speake BK. Tissue-specific differences in antioxidant distribution and susceptibility to lipid peroxidation during development of the chick embryo. Biochim Biophys Acta 1996;1304:1e10.
Surai PF, Sparks NH. Tissue-specific fatty acid and alpha-tocopherol profiles in male chickens depending on dietary tuna oil and vitamin E provision. Poult Sci 2000;79:1132e42.
Surai PF, Sparks NH. Comparative evaluation of the effect of two maternal diets on fatty acids, vitamin E and carotenoids in the chick embryo. Br Poult Sci 2001;42: 252e9.
Surai PF, Sparks NH, Noble RC. Antioxidant systems of the avian embryo: tissuespecific accumulation and distribution of vitamin E in the turkey embryo during development. Br Poult Sci 1999c;40:458e66.
Surai PF, Speake BK. Distribution of carotenoids from the yolk to the tissues of the chick embryo. J Nutr Biochem 1998a;9:645e51.
Surai PF, Speake BK. Selective excretion of yolk-derived tocotrienols into the bile of the chick embryo. Comp Biochem Physiol B Biochem Mol Biol 1998b;121: 393e6.
Surai P, Speake B, Noble R, Mezes M. Species-specific differences in the fatty acid profiles of the lipids of the yolk and of the liver of the chick. J Sci Food Agric 1999d;79:733e6.
Surai PF, Speake BK, Noble RC, Sparks NHC. Tissue-specific antioxidant profiles and susceptibility to lipid peroxidation of the newly hatched chick. Biol Trace Elem Res 1999a;68:63e78.
Surai PF, Speake BK, Sparks NHC. Carotenoids in avian nutrition and embryonic development. 2. Antioxidant properties and discrimination in embryonic tissues. J Poult Sci 2001b;38:117e45.
Surai PF, Speake BK, Sparks NHC. Carotenoids in avian nutrition and embryonic development. 1. Absorption, availability and levels in plasma and egg yolk. J Poult Sci 2001c;38:1e27.
Surai PF, Speake BK, Wood NAR, Blount JD, Bortolotti GR, Sparks NHC. Carotenoid discrimination by the avian embryo: a lesson from wild birds. Comp Biochem Physiol B 2001d;128:743e50.
Surai AP, Surai PF, Steinberg W, Wakeman WG, Speake BK, Sparks NHC. Effect of canthaxanthin content of the maternal diet on the antioxidant system of the developing chick. Br Poult Sci 2003;44:612e9.
Tarique TM, Yang S, Mohsina Z, Qiu J, Yan Z, Chen G, et al. Identification of genes involved in regulatory mechanism of pigments in broiler chickens. Genet Mol Res 2014;13:7201e16.
Tiboni GM, Bucciarelli T, Amicarelli F, Angelucci S, Iammarrone E, Belati U, et al. Spatial distribution of glutathione, glutathione related and antioxidant enzymes in cultured mouse embryos. Arch Toxicol 1997;1997(72): 38e44.
Urso UR, Dahlke F, Maiorka A, Bueno IJ, Schneider AF, Surek D, et al. Vitamin E and selenium in broiler breeder diets: effect on live performance, hatching process, and chick quality. Poult Sci 2015;94:976e83.
Wang JP, Lee JH, Jang HD, Yan L, Cho JH, Kim IH. Effects of d-aminolevulinic acid and vitamin C supplementation on iron status, production performance, blood characteristics and egg quality of laying hens. J Anim Physiol Anim Nutr Berl 2011;95:417e23.
Whitehead CC, Bollengier-Lee S, Mitchell MA, Williams PE. Vitamin E can alleviate the depressed egg production of heat-stressed laying hens. Br Poult Sci 1998;39:S44e6.
Xiao X, Yuan D, Wang YX, Zhan XA. The protective effects of different sources of maternal selenium on oxidative stressed chick embryo liver. Biol Trace Elem Res 2015 [in press]. 10 P.F. Surai et al. / Animal Nutrition 2 (2016) 1e11 Yew MS. Biosynthesis of ascorbic acid in chick embryos. Experientia 1985;41: 943e4.
Yuan D, Zhan X, Wang Y. Effects of selenium sources and levels on reproductive performance and selenium retention in broiler breeder, egg, developing embryo, and 1-day-old chick. Biol Trace Elem Res 2011;144: 705e14.
Zhang W, Zhang KY, Ding XM, Bai SP, Hernandez JM, Yao B, et al. Influence of canthaxanthin on broiler breeder reproduction, chick quality, and performance. Poult Sci 2011;90:1516e22.
Zhai W, Neuman SL, Latour MA, Hester PY. The effect of male and female supplementation of L-carnitine on reproductive traits of white leghorns. Poult Sci 2008;87:1171e81.










.jpg&w=3840&q=75)