Protective Efficacy of Different Live Attenuated Infectious Bronchitis Virus Vaccination Regimes Against Challenge With IBV Variant-2 Circulating in the Middle East
Author details:
1 Birds and Rabbit Diseases Department, Faculty of Veterinary Medicine, Sadat City University, Sadat City, Egypt; 2 Poultry Diseases Department, Faculty of Veterinary Medicine, Beni-Suef University, Beni-Suef, Egypt; 3 Poultry Diseases Department, Faculty of Veterinary Medicine, Suez Canal University, Ismailia, Egypt; 4 Virology Department, Faculty of Veterinary Medicine, Sadat City University, Sadat City, Egypt; 5 Reference Laboratory for Veterinary Quality Control on Poultry Production, Animal Health Research Institute, Giza, Egypt.
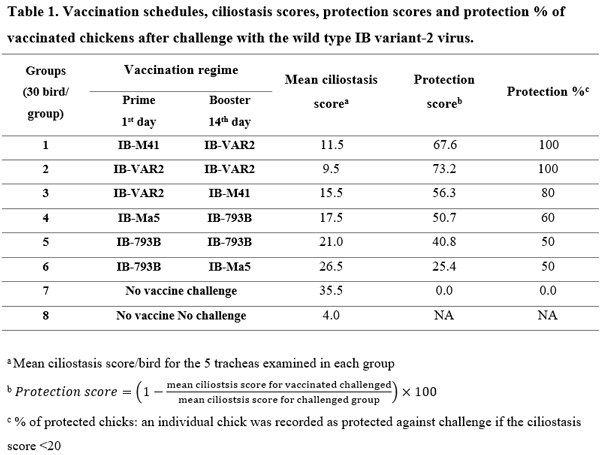
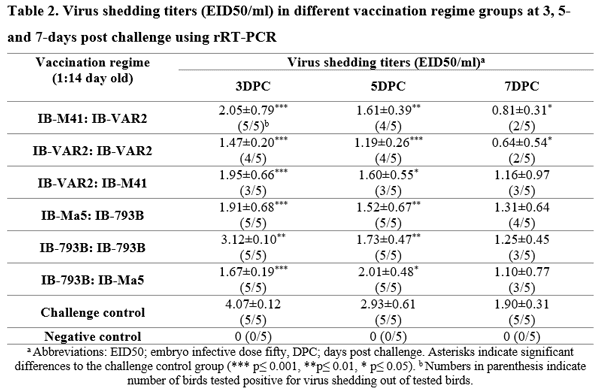
Six vaccination regimes using classical (Mass-type) and variant (IB-VAR2 and IB-793B) live vaccines were evaluated against Middle Eastern variant-2 infectious bronchitis virus challenge. Six groups of SPF chicks (30 birds/group) were vaccinated using prime-boost regimes at day-1 and day-14 using; IB-M41:IB-VAR2, IB-VAR2:IB-VAR2, IB-VAR2:IB-M41, IB-Ma5:IB-793B, IB-793B:IB-793B, and IB-793B:IB-Ma5, respectively. Ciliostasis and lesion scores were evaluated at day-5 after each vaccination. Birds were challenged intranasally at 14-day post 2nd vaccination using 105EID50/0.1ml/bird of wild-type IBV (Eg/1212B/2012). At 3, 5, and 7-day post challenge (DPC) virus shedding was monitored by real-time RT-PCR. Five chicks/group were euthanized at 7DPC for ciliostasis and lesion scoring &, and histopathology was conducted on 3 chicks/group. Seroconversion was evaluated at 14 DPC. All groups primed with the 793B vaccine showed relatively higher ciliostasis scores compared to other groups. The IB-VAR2 vaccinated groups showed the highest protection rates (80-100%) and high protection score (67.6-73.2%) compared to the 793B vaccine groups (50-60%). The virus shedding was significantly reduced at 3 and 5DPC in groups received the IBV-VAR2 (prime or booster) compared to those received the 793B vaccine. In conclusion, the homologous IBV-VAR2 vaccine showed superior results compared to 793B or Mass-type vaccines confirming the importance of IBV vaccine seed homology to the circulating IBV strains.
Keywords: IBV, live attenuated vaccine, Variant-2, Vaccination regimes, Middle East.

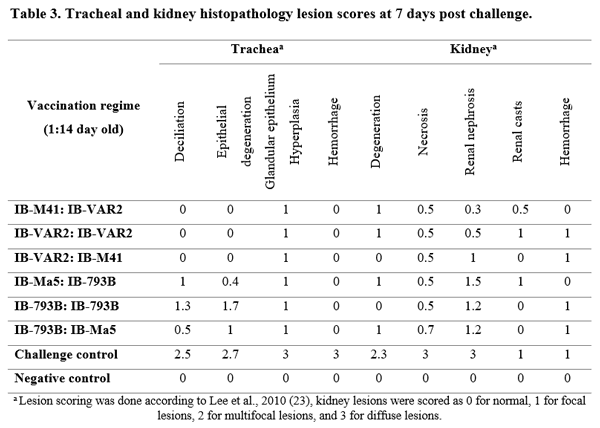
- Sid H, Benachour K, Rautenschlein S. Co-infection with Multiple Respiratory Pathogens Contributes to Increased Mortality Rates in Algerian Poultry Flocks. Avian Dis (2015) 59:440–446. doi:10.1637/11063-031615-Case.1
- Hassan KE, Shany SAS, Ali A, Dahshan AHM, El-Sawah AA, El-Kady MF. Prevalence of avian respiratory viruses in broiler flocks in Egypt. Poult Sci (2016) 95:1271–1280. doi:10.3382/ps/pew068
- Huang Q, Wang K, Pan L, Qi K, Liu H, Chen H. Co-infection of H9N2 subtype avian influenza virus and infectious bronchitis virus decreases SP-A expression level in chickens. Vet Microbiol (2017) 203:110–116. doi:10.1016/j.vetmic.2017.02.015
- Shehata AA, Sedeik ME, Elbestawy AR, Zain El-Abideen MA, Ibrahim HH, Kilany WH, Ali A. Co-infections, genetic, and antigenic relatedness of avian influenza H5N8 and H5N1 viruses in domestic and wild birds in Egypt. Poult Sci (2019) 98:2371–2379. doi:10.3382/ps/pez011
- Anonymus. Family - Coronaviridae. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ, editors. Virus Taxonomy. San Diego: Elsevier; 2012. p. 806-828. doi:https://doi.org/10.1016/B978-0-12-384684-6.00068-9
- Cook JKA. “Coronaviridae,” in Poultry Diseases (Edinburgh: W.B. Saunders), 340–349. doi:10.1016/B978-0-7020-2862-5.50033-7
- Cook JKA, Jackwood M, Jones RC. The long view: 40 years of infectious bronchitis research. Avian Pathol (2012) 41:239–250. doi:10.1080/03079457.2012.680432
- Jia W, Karaca K, Parrish CR, Naqi SA. A novel variant of avian infectious bronchitis virus resulting from recombination among three different strains. Arch Virol (1995) 140:259–271. doi:10.1007/BF01309861
- Liu S, Han Z, Chen J, Liu X, Shao Y, Kong X, Tong G, Rong J. S1 gene sequence heterogeneity of a pathogenic infectious bronchitis virus strain and its embryo-passaged, attenuated derivatives. Avian Pathol (2007) 36:231–234. doi:10.1080/03079450701338730
- Abdel-Moneim AS, Afifi MA, El-Kady MF. Emergence of a novel genotype of avian infectious bronchitis virus in Egypt. Arch Virol (2012) 157:2453–2457. doi:10.1007/s00705-012-1445-1
- Gough RE, Cox WJ, Winkler CE, Sharp MW, Spackman D. Isolation and identification of infectious bronchitis virus from pheasants. Vet Rec (1996) 138:208–209. doi:10.1136/vr.138.9.208
- Callison SA, Jackwood MW, Hilt DA. Molecular characterization of infectious bronchitis virus isolates foreign to the United States and comparison with United States isolates. Avian Dis (2001) 45:492–9. Available at: http://www.ncbi.nlm.nih.gov/pubmed/11417834
- Gelb J, Weisman Y, Ladman BS, Meir R. S1 gene characteristics and efficacy of vaccination against infectious bronchitis virus field isolates from the United States and Israel (1996 to 2000). Avian Pathol (2005) 34:194–203. doi:10.1080/03079450500096539
- Mahmood ZH, Sleman RR, Uthman AU. Isolation and molecular characterization of Sul/01/09 avian infectious bronchitis virus, indicates the emergence of a new genotype in the Middle East. Vet Microbiol (2011) 150:21–27. doi:10.1016/j.vetmic.2010.12.015
- Cook JKA, Chesher J, Baxendale W, Greenwood N, Huggins MB, Orbell SJ. Protection of chickens against renal damage caused by a nephropathogenic infectious bronchitis virus. Avian Pathol (2001) 30:423–426. doi:10.1080/03079450120066421
- Feng K, Xue Y, Wang J, Chen W, Chen F, Bi Y, Xie Q. Development and efficacy of a novel live-attenuated QX-like nephropathogenic infectious bronchitis virus vaccine in China. Vaccine (2015) 33:1113–1120. doi:10.1016/j.vaccine.2015.01.036
- Ali A, Kilany WH, Zain El-Abideen MA, Sayed M El, Elkady M. Safety and efficacy of attenuated classic and variant 2 infectious bronchitis virus candidate vaccines. Poult Sci (2018) doi:10.3382/ps/pey312
- Habibi M, Karimi V, Langeroudi AG, Ghafouri SA, Hashemzadeh M, Farahani RK, Maghsoudloo H, Abdollahi H, Seifouri P. Combination of H120 and 1/96 avian infectious bronchitis virus vaccine strains protect chickens against challenge with IS/1494/06 (variant 2)-like infectious bronchitis virus. Acta Virol (2017) 61:150–160. doi:10.4149/av_2017_02_04
- Awad F, Hutton S, Forrester A, Baylis M, Ganapathy K. Heterologous live infectious bronchitis virus vaccination in day-old commercial broiler chicks: clinical signs, ciliary health, immune responses and protection against variant infectious bronchitis viruses. Avian Pathol (2016) 45:169–177. doi:10.1080/03079457.2015.1137866
- Bru T, Vila R, Cabana M, Geerligs HJ. Protection of chickens vaccinated with combinations of commercial live infectious bronchitis vaccines containing Massachusetts, Dutch and QX-like serotypes against challenge with virulent infectious bronchitis viruses 793B and IS/1494/06 Israel variant 2. Avian Pathol (2017) 46:52–58. doi:10.1080/03079457.2016.1203393
- Lim TH, Kim MS, Jang JH, Lee DH, Park JK, Youn HN, Lee JB, Park SY, Choi IS, Song CS. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poult Sci (2012) 91:89–94. doi:10.3382/ps.2011-01739
- Terregino C, Toffan A, Serena Beato M, De Nardi R, Vascellari M, Meini A, Ortali G, Mancin M, Capua I. Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens, and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol (2008) 37:487–493. doi:10.1080/03079450802356938
- Lee HJ, Youn HN, Kwon JS, Lee YJ, Kim JH, Lee JB, Park SY, Choi IS, Song CS. Characterization of a novel live attenuated infectious bronchitis virus vaccine candidate derived from a Korean nephropathogenic strain. Vaccine (2010) 28:2887–2894. doi:10.1016/j.vaccine.2010.01.062
- Zhao Y, Cheng JL, Liu XY, Zhao J, Hu YX, Zhang GZ. Safety and efficacy of an attenuated Chinese QX-like infectious bronchitis virus strain as a candidate vaccine. Vet Microbiol (2015) 180:49–58. doi:10.1016/j.vetmic.2015.07.036
- Huo YF, Huang QH, Lu M, Wu JQ, Lin SQ, Zhu F, Zhang XM, Huang YY, Yang SS, Xu CT. Attenuation mechanism of virulent infectious bronchitis virus strain with QX genotype by continuous passage in chicken embryos. Vaccine (2016) 34:83–89. doi:10.1016/j.vaccine.2015.11.008
- Bande F, Arshad SS, Hair Bejo M, Moeini H, Omar AR. Progress and challenges toward the development of vaccines against avian infectious bronchitis. J Immunol Res (2015) doi:10.1155/2015/424860
- Abozeid HH, Paldurai A, Khattar SK, Afifi MA, El-Kady MF, El-Deeb AH, Samal SK. Complete genome sequences of two avian infectious bronchitis viruses isolated in Egypt: Evidence for genetic drift and genetic recombination in the circulating viruses. Infect Genet Evol (2017) 53:7–14. doi:10.1016/j.meegid.2017.05.006
- Hassan KE, Ali A, Shany SAS, El-Kady MF. Experimental co-infection of infectious bronchitis and low pathogenic avian influenza H9N2 viruses in commercial broiler chickens. Res Vet Sci (2017) 115:356–362. doi:10.1016/j.rvsc.2017.06.024
- Zanaty A, Naguib MM, El-Husseiny MH, Mady W, Hagag N, Arafa AS. The sequence of the full spike S1 glycoprotein of infectious bronchitis virus circulating in Egypt reveals evidence of intra-genotypic recombination. Arch Virol (2016) 161:3583–3587. doi:10.1007/s00705-016-3042-1
- Elhady MA, Ali A, Kilany WH, Elfeil WK, Ibrahim H, Nabil A, Samir A, El Sayed M. Field Efficacy of an Attenuated Infectious Bronchitis Variant 2 Virus Vaccine in Commercial Broiler Chickens. Vet Sci (2018) 5:49. doi:10.3390/vetsci5020049
- Cavanagh D, Ellis MM, Cook JKA. Relationship between sequence variation in the S1 spike protein of infectious bronchitis virus and the extent of cross-protection in vivo. Avian Pathol (1997) 26:63–74. doi:10.1080/03079459708419194
- Chen YN, Wu CC, Lin TL. Identification and characterization of a neutralizing-epitope-containing spike protein fragment in turkey coronavirus. Arch Virol (2011) 156:1525–1535. doi:10.1007/s00705-011-1020-1
- Liu S, Chen J, Han Z, Zhang Q, Shao Y, Kong X, Tong G. Infectious bronchitis virus: S1 gene characteristics of vaccines used in China and efficacy of vaccination against heterologous strains from China. Avian Pathol (2006) 35:394–399. doi:10.1080/03079450600920984
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol (2018) 35:1547–1549. doi:10.1093/molbev/msy096
- Cook JKA, Orbell SJ, Woods MA, Huggins MB. Breadth of protection of the respiratory tract provided by different live-attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol (1999) 28:477–485. doi:10.1080/03079459994506
- King DJ, Hopkins SR. Evaluation of the hemagglutination-inhibition test for measuring the response of chickens to avian infectious bronchitis virus vaccination. Avian Dis (1983) 27:100–12. doi:10.2307/1590376
- Callison SA, Hilt DA, Boynton TO, Sample BF, Robison R, Swayne DE, Jackwood MW. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J Virol Methods (2006) 138:60–65. doi:10.1016/j.jviromet.2006.07.018
- Lee CW, Suarez DL. Application of real-time RT-PCR for the quantitation and competitive replication study of H5 and H7 subtype avian influenza virus. J Virol Methods (2004) 119:151–158. doi:10.1016/j.jviromet.2004.03.014
- Chhabra R, Chantrey J, Ganapathy K. Immune Responses to Virulent and Vaccine Strains of Infectious Bronchitis Viruses in Chickens. Viral Immunol (2015) 28:478–488. doi:10.1089/vim.2015.0027
- Valastro V, Holmes EC, Britton P, Fusaro A, Jackwood MW, Cattoli G, Monne I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect Genet Evol (2016) 39:349–364. doi:10.1016/j.meegid.2016.02.015
- Jackwood MW, Hilt DA, Brown TP. Attenuation, Safety, and Efficacy of an Infectious Bronchitis Virus GA98 Serotype Vaccine. Avian Dis (2003) 47:627–632. doi:10.1637/6094
- Benyeda Z, Mató T, Süveges T, Szabó É, Kardi V, Abonyi-Tóth Z, Rusvai M, Palya V. Comparison of the pathogenicity of QX-like, M41 and 793/B infectious bronchitis strains from different pathological conditions. Avian Pathol (2009) 38:449–456. doi:10.1080/03079450903349196
- Benyeda Z, Szeredi L, Mató T, Süveges T, Balka G, Abonyi-Tóth Z, Rusvai M, Palya V. Comparative Histopathology and Immunohistochemistry of QX-like, Massachusetts and 793/B Serotypes of Infectious Bronchitis Virus Infection in Chickens. J Comp Pathol (2010) 143:276–283. doi:10.1016/j.jcpa.2010.04.007
- Gelb J, Killian SL. Serum Antibody Responses of Chickens Following Sequential Inoculations with Different Infectious Bronchitis Virus Serotypes. Avian Dis (1987) 31:513. doi:10.2307/1590733
- Zabihipetroudi MT, Ghalyanchilangeroudi A, Karimi V, Khaltabadifarahani R, Hashemzadeh M. Virus neutralization study using H120, H52, 793/B antisera against Iranian infectious bronchitis virus genotypes. Acta Virol (2018) 62:374–378. doi:10.4149/av_2018_404
- Moreno A, Franzo G, Massi P, Tosi G, Blanco A, Antilles N, Biarnes M, Majó N, Nofrarías M, Dolz R, Lelli D, Sozzi E, Lavazza A, Cecchinato M. A novel variant of the infectious bronchitis virus resulting from recombination events in Italy and Spain. Avian Pathol (2017) 46:28–35. doi:10.1080/03079457.2016.1200011
- Eldemery F, Joiner KS, Toro H, van Santen VL. Protection against infectious bronchitis virus by spike ectodomain subunit vaccine. Vaccine (2017) 35:5864–5871. doi:10.1016/j.vaccine.2017.09.013
- Ismail ZM, El-Deeb AH, El-Safty MM, Hussein HA. Enhanced pathogenicity of low-pathogenic H9N2 avian influenza virus after vaccination with infectious bronchitis live attenuated vaccine. Vet World (2018) 11:977–985. doi:10.14202/vetworld.2018.977-985









.jpg&w=3840&q=75)