The metabolic basis of feed-energy efficiency in swine
Published: June 10, 2013
By: N. A. Gutierrez; John Patience (Iowa State University, Ames, Iowa)
Introduction
It is estimated that feed comprises up to 75% of livestock production costs, and energy is the greatest single cost of diets in swine (Noblet et al., 1993). Energy yielding substances make up 70 to 90% of dietary DM, and carbons from these compounds are oxidized by the animal to produce energy needed to perform biochemical work, in the form of biosynthesis of protein and lipids, active ion transport and regulation of membrane potential, and mechanical work (muscle movement). Understanding energy supply and metabolic transformations of dietary energy in the whole animal is, therefore, of critical importance to adequately supply energy to the animal for production. It is also critical to our understanding of feed efficiency in swine.
In general, animals eat to fulfill their energetic needs when voluntary feed intake is not limited by essential nutrient deficiencies, gut capacity, disease, the physical environment and social environment. Genotype and live weight determine the potential rate of energy retention in the body and, as a consequence, the energetic needs of the pig. Energy intake and its modulation, as a consequence, often affect utilization of other nutrients and limit performance.
Prediction, optimization, and formulation of diets to achieve maximum performance, and the ability to alter product composition highlight the importance of knowledge of nutritional energetics in animal production. Additionally, many human health issues center around energy intake, such as obesity, heart disease, type II diabetes, and malnutrition.
In this chapter, aspects of partition of intake energy to fill maintenance and production demands, systems used to account for delivery and use of dietary energy, and sources of metabolic variation of energy required for maintenance and growth that will impact the efficiency of use of dietary energy are discussed. Special emphasis will be placed on sources of variation in current energy systems used for diet formulation, which will have an impact on the estimation of energy requirements and determination of usable energy from a feedstuff. For simplicity, we will restrict our review to the energy needs and metabolic efficiency of use of energy in a growing pig, and will obviate other physiological states such as lactation and reproduction.
Laws of thermodynamics
The laws of thermodynamics are fundamental in nutritional energetics for the understanding of energy flux from feed ingredients to animal tissue and fuel to support life. These laws are the basis for the development of energy models used in commercial farm animal production with the objective of supplying energy to the animal most efficiently and achieving predictable performance outcomes.
As outlined by Baldwin (1995), the first law of thermodynamics states that energy can change form, but cannot be created nor destroyed. This law is essential for assumptions and calculations utilized in nutrition, and gives rise to the factorial approach to metabolic partitioning of energy intake:
MEI = TE + HE (1)
where MEI is defined as total metabolizable energy (ME) intake, or energy consumed by an animal, and not excreted in feces or urine or lost in the form of hindgut fermentation gases; TE is energy retained in animal tissues (primarily adipose and protein); HE is the total heat released by the animal.
The second law states that all forms of energy are convertible to heat or, in the context of nutritional energetics, the driving force in all systems is to release energy as heat. In addition to the second law, the law of Hess states that heat lost in a net chemical transformation is independent of metabolic path. These are the basis of all measurements made in nutritional energetics. In brief, when these laws are taken together, "if one were to measure the heat release from a gram of fat upon total oxidation to CO2 and H2O in a laboratory calorimeter, the result will be exactly equal to the heat released when a gram of fat is total oxidized by an animal" (Baldwin, 1995).
Classical partitioning of energy
For optimization of energy content in a feed ingredient and its delivery for metabolic use in farm animal production, different energy systems have been developed and used in practice.
The concentration of gross energy (GE) in a feed ingredient is usually expressed as kilocalories (kcal) per gram, and depends on the concentration of carbohydrates, lipids, proteins and minerals in the ingredient. The heat released after combustion of 1 g of carbohydrates, lipids, or protein is 4.15, 9.50, and 5.65 kcal/g, respectively (Ewan, 2001). Gross energy is usually measured using bomb calorimetry. Gross energy intake is the total amount of energy consumed by an animal, and it represents the maximum amount of energy that is available for use by the animal.
After ingestion, only a part of the GE will be absorbed, and the remaining portion is excreted in the feces. The amount of GE in the feed that is absorbed from the gastrointestinal tract and is available for utilization by the pig is called digestible energy (DE), and is calculated by subtracting the total energy in feces from the GE content of the feed ingredient.
The metabolizable energy (ME) concentration of a feed ingredient is calculated by subtracting urinary energy (UE) and energy from fermentation gases (FE) from DE (Ewan, 2001):
ME = DE – UE – FE (2)
The ME content of a feedstuff can be calculated after placing an animal in a closed cage designed to individually deliver feed and allowing for the determination of energy lost in the urine and in the feces. Fermentation energy represents the energy in methane produced by the animal, but in most cases, the loss of energy in gas is ignored when ME is calculated, because it is negligible and difficult to measure (Chiba, 2000). The ME of a feed ingredient is, therefore, a measure of the amount of energy that is available for metabolic processes in the pig (Just, 1982b).
The ME from the feed can be lost as heat or recovered as body tissue energy (TE). The energy lost as heat is commonly referred to as heat increment (HI). This division requires either the measurement of heat loss or retained energy. Heat increment is the increase in heat production following consumption of feed when the animal is in a thermoneutral environment (Baldwin, 1995). Brody (1945) described the components of heat increment as the heat produced from the work associated with physical and chemical processes involved in mastication, digestion, absorption, and metabolism of ingested food. Baldwin (1995) divided the heat increment into 2 portions: i) the heat needed for digestion and assimilation of feed for maintenance and ii) the heat increment associated with maintaining a constant body temperature and with product synthesis.
By definition, the net energy (NE) content of a feedstuff is the energy needed for maintenance of body tissues and that accumulated in products of gain (muscle and fat). It can be calculated as the difference between ME and HI, or the heat lost in the metabolic utilization of ME (HI is an expression of the metabolic efficiency of use of ME to support maintenance functions and growth):
NE = ME – HI (3)
Heat increment includes energy lost as heat due to costs associated with digestion, absorption and assimilation of meals used for maintenance and growth, energy costs of product synthesis, synthesis of waste products, and energy lost in fermentation gases.
The NE from a feedstuff can be used for maintenance or production. Net energy for maintenance is energy used to maintain the physiological functions of the animal, e.g., to keep the animal alive and maintain a constant body temperature. The NE for production represents the NE supplied in excess of the NE of maintenance and that is used for growth or milk production (Baldwin, 1995).
The total NE of a given feed intake can, therefore, be expressed as:
NE = M + P (4)
Where M (maintenance) is an expression of NE intake used to fuel maintenance functions, and P (production) is an expression of NE intake deposited in animal tissue.
The NE system is believed to more accurately predict the energy value of high fiber feedstuffs than DE and ME systems (Patience et al., 2006) because it accounts for the lower efficiency of utilization of ME originating from hindgut fermentation than for enzymatically degraded carbohydrates (Noblet et al., 1994). Lower efficiency of use of ME from protein in feedstuffs with high protein concentration is also accounted for in the NE system. Although the NE system predicts more accurately the true energy value of a feed ingredient in pigs (Noblet et al., 1994), the ME system is most frequently used in the U.S. Given the above, it is increasingly accepted that the NE system will result in a more predictable feed efficiency outcome than either DE or ME (Patience et al., 2004).
Current estimation of maintenance requirements and costs of production
Partitioning of heat (energy used for maintenance and deposited in animal tissues) into meaningful physiologic or metabolic components is the most complex and controversial aspect of all systems and nomenclature (Baldwin, 1995).
There are multiple definitions, and dissimilar assumptions, for an animal to be considered "at maintenance". The maintenance requirement represents the ME intake required to maintain an energy balance of zero, when the animal is not thermo regulating (Baldwin, 1995). Armsby and Moulton (1925) describe maintenance as "the idea of conserving the existing status of the animal while doing no work and producing no product" and restricted maintenance to a true steady-state where "strictly there should be no translocation of material within the animal" so that protein and lipid deposition rates are zero". For animals at maintenance, Emmans (1994) also assumed a steady state with zero protein and lipid deposition. Close and Fowler (1982) stated that maintenance relates to an animal in energy equilibrium, neither losing nor gaining energy, and the energetic equivalents of body protein and lipid deposition (energy retention) is zero, but not necessarily that protein and lipid deposition rates are zero. They also reference studies that have shown the tendency in immature animals to deposit some protein and catabolize body lipids when fed near to maintenance.
For practical reasons and convenience of application, the "surface law" has traditionally been applied to calculate the maintenance requirement for energy. Heat loss is believed to be proportional to the surface area of the animal. Therefore, heat production is also proportional to the surface area because heat loss equals heat production (Brody, 1945) in homoeothermic animals. The surface law dictates that basal heat produced per unit of body weight decreases with increasing body weight (BW); hence, larger animals have lower basal metabolic rates per kilogram than do small animals (Kleiber, 1975). The allometric relationship between FHP and surface area can be described as:
FHP = ( a)BW(b) (5)
Where, FHP is a function of body weight (BW) raised to a power, and (a) is a constant. The (b) exponent varies when determined in animals of different species, sex, and age (Brody, 1945), and is determined based on actual data.
For a number of years, in nutritional energetics, the exponent (b), and as a consequence the coefficient (a), were allowed to vary from study to study and comparisons were difficult because results from the literature had to be converted to a common base. For convenience, the exponent (b) was forced, in order to attribute all variance to the coefficient (a) and to base the discussion on coefficient (a) rather than (a) and (b) (Baldwin, 1995). An initial exponent (b) of 0.75 was then used as a convention for a wide range of species. The NRC (2012) later adopted an exponent of 0.60 (Noblet et al., 1994) to estimate FHP in growing pigs but not in sows. These exponents were obtained from fit of data that best described the relationship of FHP and body weight. Fixed exponents were adopted to standardize the expression of data. However, this approach is based on empirical best-fit relationships of data and, therefore, is inappropriate to speculate on the physiological meaning of the exponents and coefficients.
To determine the maintenance energy requirement in pigs, measurements of heat production can be obtained either directly or indirectly using calorimeters (respiration chambers). Another approach is to estimate energy retention by comparing the total energy content of groups of animals before and after an experimental period using comparative slaughter (Just et al., 1982a,b).
When values from indirect calorimetry and comparative slaughter are compared, it should be considered that animals are restrained in environmentally controlled respiration chambers, limiting activity, social interaction, and temperature regulation. As a consequence, energy retention tends to be lower if measured using comparative slaughter procedures compared with indirect calorimetry (Reynolds, 2000). Values of FHP determined in respiration chambers tend to overlook major environmental factors that may influence heat production, such as ambient temperature and physical activity of the animal (NRC, 2012). Therefore, a decrease in environmental temperature below the critical temperature results in an increase in energy expenditure because of the necessity to produce heat (Noblet et al., 1985).
In order to undertake statistical analysis of relationships between ME intake (MEI) and body energy changes, basic equations have been developed. According to Baldwin (1995):
MEm = FCAT/ km (6)
Where MEm is the ME required for maintenance; fasting catabolism (FCAT) is an estimate of total energy loss of a fasting animal and comprises FHP plus fecal and urine energy of endogenous origin, and costs of waste synthesis of an animal fed at maintenance; km is the efficiency of use of ME for maintenance.
TEg = ( MEI-MEm)/kg (7)
Where TEg represents the energy from feed, supplied above maintenance, that is retained in muscle and adipose tissue; kg is the efficiency of use of ME provided above maintenance for tissue energy gain. Thus, from equations (6) and (7), the intake of ME (MEI) of a growing pig can be expressed as:
MEI = FCAT/km + TEg/ kg (8)
The factorial approach, as is widely used in nutritional energetics, is a statistical model designed to interpret the energy content of a feedstuff in relation to its metabolic use.
The empirical nature of the values of FCAT and TE, and the methodology used to determine the values, restrict the application of energy partition models to similar conditions under which they were determined. Efficiency of utilization of energy in a feedstuff or diet can be affected by the poor interpretation or application of energy systems.
It can therefore be seen that measurement of feed efficiency is rather more complex and simply dividing growth rate for a given period by feed consumption during the same period. Differences in feed efficiency can quite simply be the result of changing dietary energy content, when the more important considerations are how much energy is used for maintenance and how much dietary energy consumed is actually available for growth.
Problems of the factorial approach
When estimating energy required for maintenance functions by the pig, the factorial/empirical approach can describe carefully collected experimental data very well, because estimates of FHP are the product of best fit relationships with BW. The use of the allometric relationship to express energy needed to support maintenance functions is, however, a major over-simplification of reality. Maintenance involves several significant body processes including those associated with FHP measured when the pig is lying in a recumbent position, normal activity, and activity associated with feeding, the maintenance of body temperature, and changes related to disease (Black, 1995). Additionally, Brody (1945) argued that surface area might not be the driving force in evolution because of the big differences among species in subcutaneous fat, skin thermo conductivity, covering of hair, fur and/or feathers, and mechanisms developed for thermoregulation.
Young et al. (1989) reported elevated metabolic rates during the stage of highest growth of growing sheep. These elevated physiological process are mainly related to increased nutrient intake (foraging and feed intake activity; digestion and its associate enzyme production; sloughing and turnover of GIT; and excretion) and intensified cellular functions such as membrane transport and associated protein turnover (Knap, 2009). In pigs, modern fast growing genotypes selected to deposit more protein have been associated with an increased metabolic intensity, and as a consequence, the metabolic costs of maintenance have also increased (Kolstad and Vangen, 1996).
Current estimates of FHP using calorimetry, however, are carried out in animals fed at maintenance, or at zero energy retention. Animals fed at zero energy retention, however, can make metabolic adjustments to mimic mature animals because of the necessity to reduce energy needs due to growth, and can be considered at "steady state". Therefore, energy models constructed with data from FHP at zero energy retention, and designed to include all the energy needs for maintenance work at zero energy retention well for an animal in "steady state".
But this is not the case of a normal immature animal, because the steady state situation excludes ongoing dynamic processes such as growth, reproduction, lactation, and physical work beyond that of apprehension of feed. As a consequence, maintenance needs of immature animals fed to maintain zero energy retention give suspicious estimates, or more representative of animals at maturity rather than the elevated metabolic intensity of growing animals (Knap, 2009).
The variability of maintenance requirements is high and also known to be affected by sex, age, genetics, and environment. The exponents and coefficients of the allometric relationship are based on a pure empirical fit of data, resulting in the impossibility of assigning a biological meaning to the coefficients. The empirical nature of the values for FHP, and the methodology used for determination, restrict the application of energy partition models to conditions similar to which they were derived. The same situation occurs for empirically derived efficiencies of utilization of lipids and protein for growth when feed is supplied above maintenance. As a consequence, efficiency of utilization of energy in a feedstuff is affected by the poor interpretation or application of energy systems.
One solution has been to develop more complex statistical models, including more variables, with the objective to account for factors which may contribute to variation in energy required for maintenance functions. Parameters accompanying these variables are derived empirically from multiple regression analysis of large data sets. The statistical approach, however, has some limitations, as outlined by Knap (2009): 1) The number of parameters that can be incorporated into the model to make it more mechanistic is limited; 2) the independent variables (i.e. km, kg) are highly inter-correlated, violating the assumption of independence of variance of the multiple regression analysis 3) as a result of the inter-correlation, partial protein and lipid accretion are highly variable (larger error of independent variables). These disadvantages cause the associated parameter estimates of the model to be confounded and biased, which makes interpretation hazardous.
Additionally, actions such as immune response, thermoregulation and coping with other stressors are usually excluded from FHP because of the methodology used for determination. A similar situation applies to physical activity beyond the basic level (especially in young animals) and for physiological service and cell maintenance functions in feeding levels above maintenance. This implies a difficulty of separating out the costs involved in maintenance (FHP) and growth in the factorial approach.
The picture that arises is one of confusion. There is neither general agreement about what maintenance represents nor about its components, and most descriptions are of qualitative nature. Rather than best fit relationships of data, Baldwin (1995) favors, over the long term, the explicit consideration of physiological and metabolic functions to better explain variance in FHP due to the influence of age, physiological state, previous and current planes of nutrition etc., and its interaction with growth processes. Progress has been made, but the goal of understandingmechanisms and relationships has not yet been achieved. The current situation is not likely to change in the future and, therefore, the main issue is to understand which metabolic processes contribute to variation in main maintenance and growth supporting functions (Knap, 2009).
Metabolic sources of variation in energy expenditure
Costs of digestion and assimilation
The energy lost as heat due to the cost of digestion, absorption and assimilation (storage) of a carbohydrate meal fed at maintenance in monogastric animals were outlined by Baldwin (1995) and are presented in Table 1. From the table, it can be concluded that the energy costs of absorption of a meal varies depending on its carbohydrate, lipid, and AA composition. The number of high energy phosphates (ATP) expended per mole of nutrient absorbed vary from 0.5 ~ P per mole of AA, 1 ~ P per mole of glucose, to 4 ~ P per mole of TAG. As a result, the energy lost as a % of the GE in the nutrient is 2.6, 0.6, and 1.9 % for glucose, tripalmitin, and AA, respectively. The energy cost of synthesis of digestive proteins is minor, but changes depending on the methodology used to calculate it (indirect calorimetry vs. comparative slaughter).
Table 1: Estimated costs of digestion, absorption, and assimilation of meals fed at maintenance.1,2,3
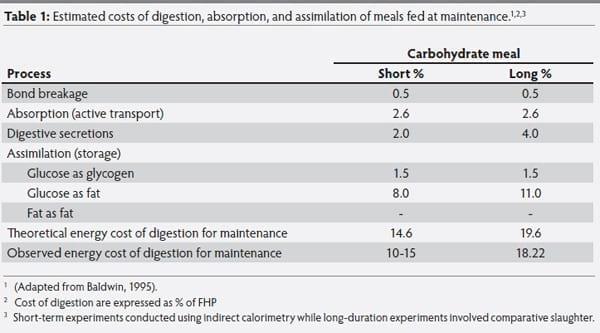
To exemplify the effect on energy costs of assimilation/ storage of different dietary nutrients, Baldwin (1995) calculated theoretically, for single stomached animals, the energy losses due to metabolic costs for synthesis of end products, and found that when glucose is stored as glycogen in liver and muscle, 5.2 and 2.6% of glucose energy is lost, respectively. The energy losses associated with the conversion of glucose to fat, are 16.2 and 22.8% in short term and long term studies, respectively. On the other hand, when AA are used to produce high energy phosphates, the associated energy costs were estimated at 15% of the GE contained in AA, mostly driven by the cost of gluconeogenesis in the liver.
After digestion and absorption, when nutrients are used to fulfill maintenance functions, the heat released varies depending on the substrate oxidized. This is an important source of variation in feed efficiency, because heat released from dietary glucose, lipids or protein is used to produce ATP, which is the energy currency to fuel maintenance functions. The amount of energy needed to produce ATP can vary significantly dependent upon the substrate oxidized to support that process (Table 2). The organization of substrate oxidation presented in Table 2 clearly illustrates the effect of differing nutrients used to fill energy requirements. For example, when protein and acetate are used as energy sources to produce ATP, respective heat losses relative to glucose are 28 and 18% greater.
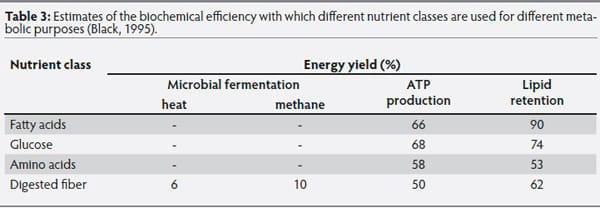
Black (1995), summarized the theoretical efficiencies of utilization of major nutrients absorbed for maintenance functions and lipid retention in growing pigs (Table 3). He based his calculations on the stoichiometry of biochemi-cal reactions to calculate the average efficiency of use of the total energy of different nutrients for different body functions. This type of calculation is of critical importance because energy models used today assume a constant efficiency of use of available energy (from dietary nutrients) for maintenance and growth. The initial approach proposed by Black is important in understanding possible ways to improve feed efficiency, because it is necessary for energy models to be capable of predicting which nutrients are used for particular purposes and to apply the appropriate energetic efficiencies.
Table 2: Heat production associated with the formation and use of high energy ATP bonds in support of maintenance processes1.
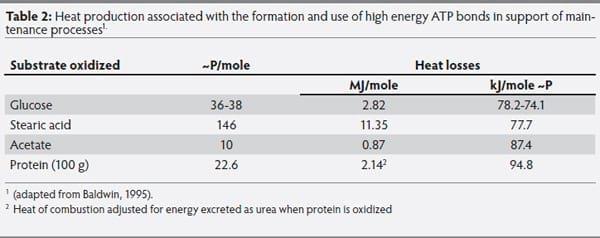
Table 3: Estimates of the biochemical efficiency with which different nutrient classes are used for different metabolic purposes (Black, 1995).
Components of fasting heat production
Breed, age, sex, physiological state, previous and current planes of nutrition, etc., contribute to big differences observed in requirements for MEm in pigs. Differences in efficiency may be explained by determining components of FHP as well as their contribution to the aggregate. They can also be explained by analysis of tissue-by-tissue functions and its relation with any of the factors contributing with variance of ME m.
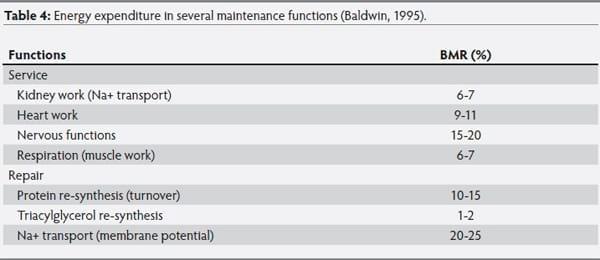
Baldwin (1995) reported the energy expenditures in several maintenance functions, calculated for a 70 kg man at a basal metabolic rate (equivalent of FHP) of 7.1 MJ/d, as summarized in Table 4. These functions agree with the tentative partitioning of MEm reported by Knap (2009), after conducting simulation studies in growing pigs under thermoneutral, welfare-friendly, and healthy individual housing conditions. The author reported five main component processes of the maintenance aggregate: 1)
Service functions, 2) membrane transport, 3) protein turnover, 4) basal activity and 5) additional functions, such as social stress, extra activity, immune response, and cold stress. Service functions are performed by an organ or tissue, in support of another organ function or performance of the whole organism. From Table 4, heat production from service functions accounts for approximately 40% of total energy expenditure. Ion transport is a significant contributor to FHP and constitutes approximately 25%. The sum of service, ion transport, and turnover functions is approximately 80% of FHP.
According to Baldwin, (1995), energy expenditures for service functions such as blood flow and respiration are dynamic, because they vary with food intake, activity, and rate of production. Both functions vary continuously and manifest as components of both maintenance energy expenditure and cost of production in animals on low and high planes of nutrition. Protein turnover and active transport of ions (both part of repair functions) have the same problem. Others functions such as kidney work and nervous functions remain constant. In the statistical approach, blood flow and protein turnover are usually partitioned between maintenance and costs associated with production. This constitutes a problem, because it is not possible to relate the dynamic effects of growth on cardiac output and protein turnover to the estimates of efficiency for maintenance or for growth.
Table 4: Energy expenditure in several maintenance functions (Baldwin, 1995).
An alternative approach to analyses of contributors to FHP is to undertake a tissue-by-tissue evaluation, in which estimates of weight of organs and/or tissues, and energy expenditures by organs and/or tissues are analyzed. Baldwin (1995) reported a tremendous range of heat production (as % FHP) per unit of weight of a given tissue or organ. Although muscle and liver account for almost 20% of total FHP each, and muscle accounts for almost 41% of empty body mass whereas liver accounts for only 1.6%, heat production of liver per unit weight is 25 times greater than muscle. Among tissues and organs, heat production per unit weight ranged from 0.22 to 23. The overall estimate of FHP is an average of energy expenditure per unit weight of diverse tissues, and nutritional and physiological factors that can cause changes in relative weights of organs and tissues can cause significant changes in the FHP estimate (Baldwin, 1995). Smith and Baldwin (1974) reported an increase of 10% in FHP due to the change in relative organ weights associated with lactation in cows.
The effects of feed intake on the maintenance requirements of energy were studied by Ferrel (1978) when rats previously provided high as compared to low planes of nutrition were studied. Rats under a high plane of nutrition had 38% greater maintenance requirements than rats under a low plane of nutrition. Besides the effect of increased organ size per se on FHP increase, changes in tissue function ma also impact FHP (previous plane of nutrition), physiological and hormonal state, and relative organ weights.
Conclusion
We have tried to identify, in spite of the enormous disparity and lack of knowledge in the use and partition of dietary energy, a large proportion of energy costs associated with energy requirements and causes of variation of these. Further experimentation and modeling exercises are necessary to develop systems in which energy costs of maintenance and production do not need to be considered separated and excluded, but as a continuous function of underlying processes, which are known and characterized to the extent that they can be expressed mathematically, and be used in the development of systems for optimization of feed resources for use in animal production.
With this in mind, we can see the tremendous challenge in understanding the scientific basis of feed efficiency. Since maintenance utilizes up to 35% of total energy intake (Oresanya et al., 2008), it is apparent that greater attention must be paid to barn management procedures that minimize maintenance energy utilization.
Finally, "feed efficiency" embraces the utilization of amino acids as well as energy in growth and maintenance. We have simply restricted this discussion to energy, because it appears to be the least understood component of the diet, but also by far the most costly to supply to the pig.
References
1. Armsby, H. P., C. R. Moulton. 1975. The animal as a converter or matter and energy. Chemical catalog, New York, NY.
2. Baldwin, R. L. 1995. Modeling Ruminant Digestion and Metabolism. Chapman and Hall, New York, NY.
3. Brody, S. 1945. Bioenergetics and Growth. Reinhold publishing Corp., New York, NY.
4. Close, W. H., V.R. Fowler. 1982. Energy requirements of pigs. Pages 1–16 in Recent Advances in Animal Nutrition. W. Haresign, ed. Butterworths, London.
5. Chiba, L. I. 2000. Feeding systems for pigs. Pages 181–209 in Feeding Systems and Feed Evaluation Models. M. K. Theodorou and J. France, eds. CAB International, Wallingford, United Kingdom.
6. Emmans, G. C. 1994. Effective energy: a concept of energy utilization applied across species. Br. J. Nutr. 71: 801–821.
7. Ewan, R. C. 2001. Energy utilization in swine nutrition. Pages 85–124 in Swine Nutrition. A. J. Lewis and L. L. Southern, ed. CRC Press, Boca Raton, FL.
8. Ferrel, C. L. 1984. Sources of variation in animal energy expenditures. Pages 100–109 in Modeling Ruminant Digestion and Metabolism. Proceedings of the Second International Workshop. R. L. Baldwin and A. C. Bywater, eds. Department of Animal Science, University of California, Davis, CA.
9. Just, A. 1982a. The net energy value of crude fat for growth in pigs. Livest. Prod. Sci. 9: 349–360.
10. Just, A. 1982b. The net energy value of crude (catabolized) protein for growth in pigs. Livest. Prod. Sci. 9: 569–580.
11. Kleiber, M. 1975. The fire of life: an introduction to animal energetics. John Wiley, New York, NY.
12. Kolstad, K., O. Vangen. 1996. Breed differences in maintenance requirements of growing pigs when accounting for changes in body composition. Livest. Prod. Sci. 47: 23–32.
13. Noblet, J., H. Fortune, X. S. Shi, and S. Dubois. 1994. Prediction of net energy value of feeds for growing pigs. J. Anim. Sci. 72: 344–353.
14. Noblet, J., H. Fortune, C. Dupire, S. Dubois. 1993. Digestible, metabolizable and net energy values of 13 feedstuffs for growing pigs: effect of energy system. J. Anim. Sci. 42: 131–149.
15. Noblet, J., J. Le Dividich, T. Bikawa. 1985. Interaction between energy level in the diet and environmental temperature on the utilization of energy in growing pigs. J. Anim. Sci. 61: 452–459.
16. N.R.C. 2012. Nutrient Requirements of Swine. 11th rev. ed. Natl. Acad. Press, Washington, DC.
17. Oresanya, T. F., A. D. Beaulieu, J. F. Patience. 2008. Investigations of energy metabolism in weanling barrows: The interaction of dietary energy concentration and daily feed (energy) intake. J. Anim. Sci. 86: 348– 363.
18. Patience, J. F., P. Leterme, A. D. Beaulieu. 2006. Advantages of the net energy system. Proc. Pre-symposium Worshop "Net Energy Systems For Growing And Finishing Pigs." 10th Int'l Symp. Digestive Physiology in Pigs, Vejle, Denmark.
19. Patience, J. F., A. D. Beaulieu, R. T. Zijlstra, T. Oresanya, R. Mohr. 2004. Energy systems for swine: A critical review of DE, ME and NE. Proc. Midwest Swine Nutr. Conf. Indianapolis, IN.
20. Knap, P. W. 2009. Allocation of resources to maintenance. Pages 118–136 in Resource allocation theory applied to farm animal production. W. M. Rauw, ed. CAB International, Wallingford, UK.
21. Reynolds, C. K. 2000. Measurement of energy metabolism. Page 87–107 in Feeding Systems and Feed Evaluation Models. M. K. Theodorou and J. France, eds. CAB International, Wallingford, UK.
22. Smith, N. E., R. L. Baldwin. 1974. Effects of breed, pregnancy, and lactation on weights of organs and tissues in dairy cattle. J. Dairy Sci. 57: 1055–1065.
23. Young, B. A., A. W. Bell, T. R. Hardin. 1989. Mass specific metabolic rate of sheep from fetal life to maturity. Pages 155–158 In Energy Metabolism of Farm Animals. Y. van der Honing, and W. H. Close, eds. EAAP publication 43. Pudoc, Wageningnen, The Netherlands.
This paper was presented at the Allen D. Leman Swine Conference, held Sept. 15-18 at the St. Paul River Center in St. Paul, Minnesota. Published by Veterinary Continuing Education, Volume 39 2012. Engormix thanks the university and the author for this huge contribution.
The University of Minnesota, Allen D. Leman Swine Conference is an annual educational event for the global swine industry. It is internationally acclaimed for bringing science-driven solutions to the complex challenges facing the industry. Each year hundreds of participants from over 20 countries attend the Leman Swine Conference held in St. Paul, Minnesota, USA. Major players in swine production, animal health management, and marketing from around the world exhibit their products and services.
To see 2013 edition, click here
Related topics
Authors:

Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post
IFF - International Flavors & Fragrances
29 de agosto de 2013
Hello Mr. Patience, my congratulations for this paper: excellent review of something which I believe most of us, practical Nutritionist, tend to, not forget, but not consider that much in our daily work. In any case I would be very interested in getting your wise answer on the following: as you know more and more pig diets are increasingly, especially in Europe, including more fibre-like ingredients (wheat by products, rapeseed meal, sunflower meal, wheat-DDGS...). We reformulate to isoenergetic diets by increasing the level of fat in diet, but level of starch vs NDF is reduced in these diets (I formulate in NE). My question is how would this higher NDF level would affect the HI - I would assume that this loss is not anymore negligible - thus NE for growth and what is your opinion in the role, in this more fibrous diets, of carbohydrases. Many thanks from Madrid, Spain.





You may be interested in
.jpg&w=3840&q=75)
Evonik Animal Nutrition