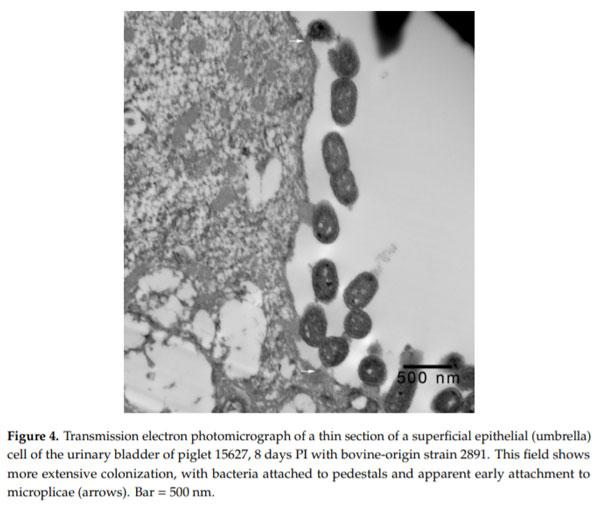
Intimate Attachment of Escherichia coli O157:H7 to Urinary Bladder Epithelium in the Gnotobiotic Piglet Model
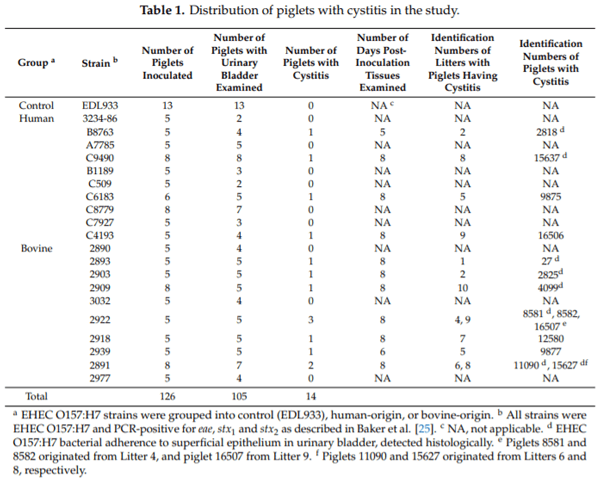
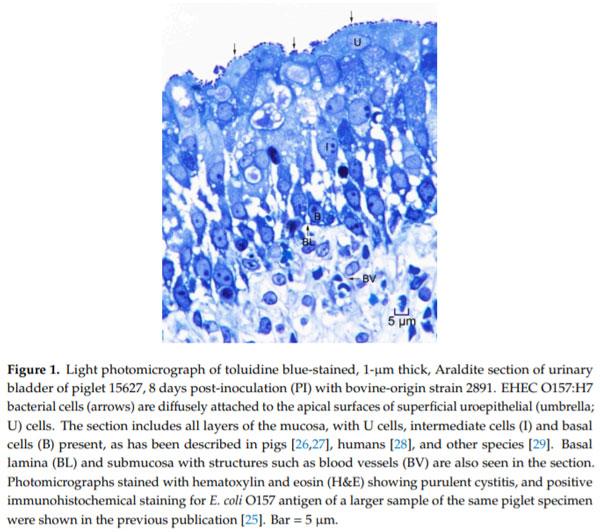
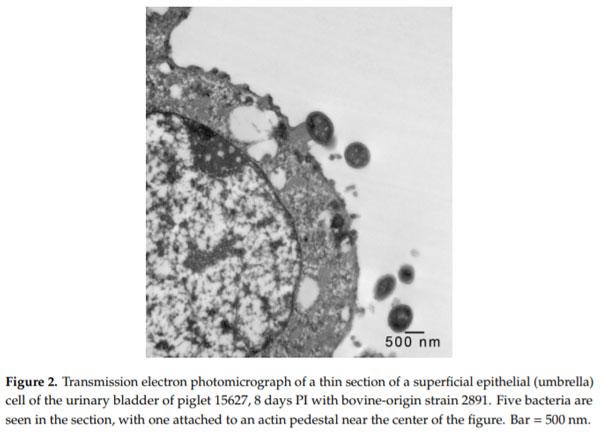
Enterohemorrhagic Escherichia coli (EHEC), a pathogenic subset of Shiga toxin-producing E. coli (STEC), is an important cause of hemorrhagic colitis and hemolytic–uremic syndrome (HUS), and a rare cause of urinary tract infections (UTIs) with associated HUS. EHEC strains attach intimately to intestinal epithelium with formation of actin pedestals (attaching-effacing (A/E) lesions); however, the mechanism of EHEC attachment to the uroepithelium is unknown. We conducted a retrospective study on archived urinary bladder specimens from gnotobiotic piglets that naturally developed cystitis associated with EHEC O157:H7 infection following oral inoculation and fecal shedding. Paraffin-embedded bladder tissues from three piglets with cystitis and immunohistochemical evidence of EHEC O157:H7 adherence to the uroepithelium were processed for and examined by transmission electron microscopy. EHEC O157:H7 bacteria were found in one of three piglets, intimately attached to pedestals on the apical surfaces of the superficial urothelium (umbrella cells). Cystitis was significantly associated with the length of survival of the piglets post-inoculation (p = 0.0339; estimated odds ratio = 2.6652). This is the first report of E. coli causing A/E-like lesions in the uroepithelium, and also evidence of the utility of the gnotobiotic piglet as a model for studies of the pathogenesis of EHEC UTIs.
Keywords: Shiga toxin-producing E. coli; enterohemorrhagic E. coli; attaching-effacing; pedestals; urinary tract infection; cystitis; pigs; gnotobiotic piglets
1. Majowicz, S.E.; Scallan, E.; Jones-Bitton, A.; Sargeant, J.M.; Stapleton, J.; Angulo, F.J.; Yeung, D.H.; Kirk, M.D. Global incidence of human Shiga toxin-producing Escherichia coli infections and deaths: A systematic review and knowledge synthesis. Foodborne Pathog. Dis. 2014, 11, 447–455. [CrossRef] [PubMed]
2. Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [CrossRef]
3. Kavanagh, D.; Raman, S.; Sheerin, N.S. Management of hemolytic uremic syndrome. F1000Prime Rep. 2014, 6, 119. [CrossRef] [PubMed]
4. Centers for Disease Control. Isolation of E. coli O157:H7 from sporadic cases of hemorrhagic colitis—United States. Morb. Mortal. Wkly. Rep. 1982, 31, 580–585.
5. Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 1983, 308, 681–685. [CrossRef]
6. Remis, R.S.; MacDonald, K.L.; Riley, L.W.; Puhr, N.D.; Wells, J.G.; Davis, B.R.; Blake, P.A.; Cohen, M.L. Sporadic cases of hemorrhagic colitis associated with Escherichia coli O157:H7. Ann. Intern. Med. 1984, 101, 624–626. [CrossRef]
7. Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [CrossRef]
8. Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [CrossRef]
9. Gould, L.H.; Mody, R.K.; Ong, K.L.; Clogher, P.; Cronquist, A.B.; Garman, K.N.; Lathrop, S.; Medus, C.; Spina, N.L.; Webb, T.H.; et al. Increased recognition of non-O157 Shiga toxin-producing Escherichia coli infections in the United States during 2000–2010: Epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 2013, 10, 453–460. [CrossRef]
10. Luna-Gierke, R.E.; Griffin, P.M.; Gould, L.H.; Herman, K.; Bopp, C.A.; Strockbine, N.; Mody, R.K. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol. Infect. 2014, 142, 2270–2280. [CrossRef]
11. Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [CrossRef] [PubMed]
12. Lavrek, D.; Lava, S.A.G.; Milani, G.P.; Simonetti, G.D.; Bianchetti, M.G.; Giannini, O. Hemolytic-uremic syndrome after Escherichia coli urinary tract infection in humans: systematic review of the literature. J. Nephrol. 2018, 31, 919–924. [CrossRef] [PubMed]
13. Kalita, A.; Hu, J.; Torres, A.G. Recent advances in adherence and invasion of pathogenic Escherichia coli. Curr. Opin. Infect. Dis. 2014, 27, 459–464. [CrossRef] [PubMed]
14. Nielubowicz, G.R.; Mobley, H.L.T. Host-pathogen interactions in urinary tract infection. Nat. Rev. Urol. 2010, 7, 430–441. [CrossRef] [PubMed]
15. Robino, L.; Scavone, P.; Araujo, L.; Algorta, G.; Zunino, P.; Pírez, M.C. Intracellular bacteria in the pathogenesis of Escherichia coli urinary tract infection in children. Clin. Infect. Dis. 2014, 59, e158–e164. [CrossRef] [PubMed]
16. Mulvey, M.A.; Schilling, J.D.; Martinez, J.J.; Hultgren, S.J. Bad bugs and beleaguered bladders: interplay between uropathogenic Escherichia coli and innate host defenses. Proc. Natl. Acad. Sci. USA 2000, 97, 8829–8835. [CrossRef]
17. Mulvey, M.A. Adhesion and entry of uropathogenic Escherichia coli. Cell. Microbiol. 2002, 4, 257–271. [CrossRef]
18. Rosen, D.A.; Hooton, T.M.; Stamm, W.E.; Humphrey, P.A.; Hultgren, S.J. Detection of intracellular bacterial communities in human urinary tract infection. PLoS Med. 2007, 4, e329. [CrossRef]
19. Wiles, T.J.; Kulesus, R.R.; Mulvey, M.A. Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp. Mol. Pathol. 2008, 85, 11–19. [CrossRef]
20. Mobley, H.L.; Donnenberg, M.S.; Hagan, E.C. Uropathogenic Escherichia coli. EcoSal Plus 2013. [CrossRef]
21. Tamadonfar, K.O.; Omattage, N.S.; Spaulding, C.N.; Hulgren, S.J. Reaching the end of the line: urinary tract infections. Microbiol. Spectrum 2019, 7, BAI-0014-2019. [CrossRef]
22. Eberly, A.R.; Beebout, C.J.; Tong, C.M.C.; Van Horn, G.T.; Green, H.D.; Fitzgerald, M.J.; De, S.; Apple, E.K.; Schrimpe-Rutledge, A.C.; Codreanu, S.G.; et al. Defining a molecular signature for uropathogenic versus urocolonizing Escherichia coli: The status of the field and new clinical opportunities. J. Mol. Biol. 2019. [CrossRef]
23. Moon, H.W.; Whipp, S.C.; Argenzio, R.A.; Levine, M.M.; Giannella, R.A. Attaching and effacing activities of rabbit and human enteropathogenic Escherichia coli in pig and rabbit intestines. Infect. Immun. 1983, 41, 1340–1351. [CrossRef] [PubMed]
24. Lai, Y.; Rosenshine, I.; Leong, J.M.; Frankel, G. Intimate host attachment: Enteropathogenic and enterohaemorrhagic Escherichia coli. Cell. Microbiol. 2013, 15, 1796–1808. [PubMed]
25. Baker, D.R.; Moxley, R.A.; Steele, M.B.; LeJeune, J.T.; Christopher-Hennings, J.; Chen, D.-G.; Hardwidge, P.R.; Francis, D.H. Differences in virulence among Escherichia coli O157:H7 strains isolated from humans during disease outbreaks and from healthy cattle. Appl. Environ. Microbiol. 2007, 73, 7338–7346. [CrossRef] [PubMed]
26. Scheidegger, G. Der aufbau des übergangsepithels der harnblase bei schwein, schaf, ratte und spitzmaus (Structure of the transitional epithelium in the urinary bladder of the pig, sheep, rat and shrew). Acta Anat. 1980, 107, 268–275. [CrossRef] [PubMed]
27. Liebhold, M.; Wendt, M.; Kaup, F.-J.; Drommer, W. Licht- und elektronenmikroskopische studien zur struktur des normalen blasenepithels beim weiblichen schwein (Light- and electron-microscope study of the structure of the normal bladder epithelium in female pigs). Anat. Histol. Embryol. 1995, 24, 47–52. [CrossRef]
28. Khandelwal, P.; Abraham, S.N.; Apodaca, G. Cell biology and physiology of the uroepithelium. Am. J. Physiol. Renal Physiol. 2009, 297, F1477–F1501. [CrossRef]
29. Hicks, R.M. The mammalian urinary bladder: An accommodating organ. Biol. Rev. Camb. Phil. Soc. 1975, 50, 215–246. [CrossRef]
30. Staley, T.E.; Jones, E.W.; Corley, L.D. Attachment and penetration of Escherichia coli into intestinal epithelium of the ileum in newborn pigs. Am. J. Pathol. 1969, 56, 371–392.
31. Takeuchi, A.; Inman, L.R.; O’Hanley, P.D.; Cantey, J.R.; Lushbaugh, W.B. Scanning and transmission electron microscopic study of Escherichia coli O15 (RDEC-1) enteric infection in rabbits. Infect. Immun. 1978, 19, 686–694. [CrossRef] [PubMed]
32. Ulshen, M.H.; Rollo, J.L. Pathogenesis of Escherichia coli gastroenteritis in man—Another mechanism. N. Engl. J. Med. 1980, 302, 99–101. [CrossRef] [PubMed]
33. Rothbaum, R.; McAdams, A.J.; Giannella, R.; Partin, J.C. A clinicopathologic study of enterocyte-adherent Escherichia coli: A cause of protracted diarrhea in infants. Gastroenterology 1982, 83, 441–454. [CrossRef]
34. Knutton, S.; Baldwin, T.; Williams, P.H.; McNeish, A.S. Actin accumulation at sites of bacterial adhesion to tissue culture cells: Basis of a new diagnostic test for enteropathogenic and enterohemorrhagic Escherichia coli. Infect. Immun. 1989, 57, 1290–1298. [CrossRef] [PubMed]
35. Geerlings, S.E. Clinical presentations and epidemiology of urinary tract infections. Microbiol. Spectrum 2016, 4, UTI-0002-2012. [CrossRef] [PubMed] 36. Oelschlaeger, T.A.; Barrett, T.J.; Kopecko, D.J. Some structures and processes of human epithelial cells involved in uptake of enterohemorrhagic Escherichia coli O157:H7 strains. Infect. Immun. 1994, 62, 5142–5150. [CrossRef]
37. Turner, A.M.; Subramaniam, R.; Thomas, D.F.M.; Southgate, J. Generation of a functional, differentiated porcine urothelial tissue in vitro. Eur. Urol. 2008, 54, 1423–1432. [CrossRef]







