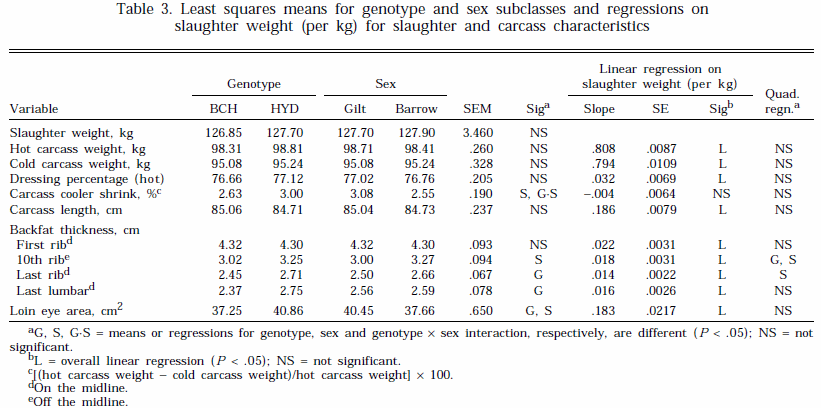
slaughter weight between the sexes for 10th rib and last rib backfat and between the genotypes for 10th rib backfat (Table 3). However, these quadratic relationships were of limited practical significance. The regressions of first rib and last lumbar fat depths and loin eye area on SLW were linear with no effect of sex or genotype. These data suggest that the overall rate of backfat deposition did not increase with SLW. Gu et al. (1992) also found a linear effect of live weight on carcass length, backfat depths, and loin eye area in five genotypes of pigs slaughtered between 59 and 127 kg live weight. However, they also reported a curvilinear increase in carcass fat weight relative to SLW, suggesting an increase in fat accretion and carcass fat percentage with animal weight. This highlights the limitations with using linear measurements taken at a limited number of positions on the carcass to predict body composition.
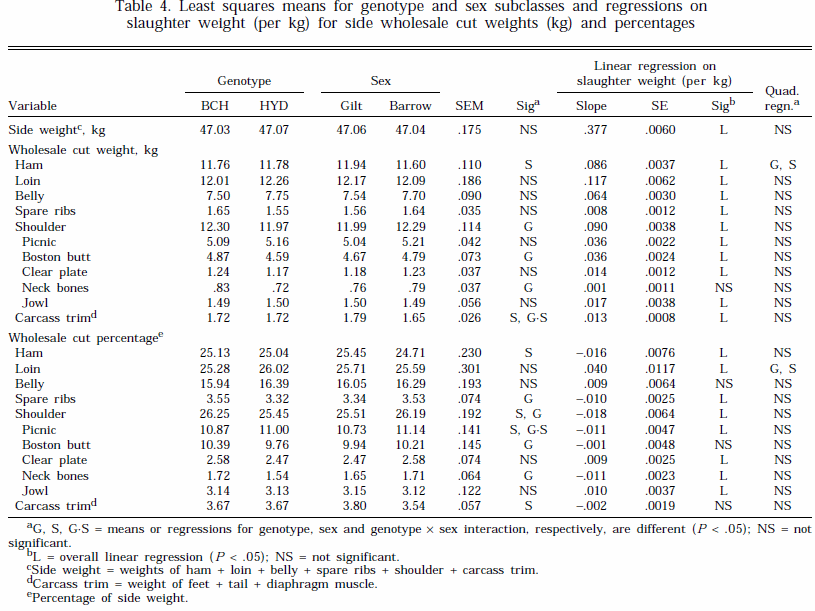
Results for wholesale cut weights and percentages are presented in Table 4. Side weights were similar across all treatment groups. There were genotype differences for the weight of shoulder, boston butt, and neck bones, and the percentages of spare ribs, shoulder, boston butt, and neck bones. The sexes differed for the weight and percentage of ham and carcass trim and the percentage of shoulder and picnic (Table 4). In addition, there were genotype ´ sex interactions for carcass trim weight and percentage of picnic ( P < .001). Gilts from BCH had greater carcass trim than barrows (1.82 vs 1.62 kg, respectively, P < .001), whereas there was no difference between the sexes for HYD (1.76 vs 1.69 kg, respectively). Picnic percentage of HYD barrows was greater than for gilts (11.43 vs 10.57%, respectively, P < .05); however, there was little difference between the sexes for BCH (10.85 vs 10.90%, respectively). Overall, however, the genotype and sex differences and the quadratic regressions on SLW for cut weights and proportions observed in this study were of limited practical significance.
As expected, the weight of all of the wholesale cuts increased with slaughter weight (Table 4). In addition, the percentage of loin, clear plate, and jowl increased and ham, shoulder, picnic, and spare rib percentages decreased with slaughter weight. The percentage of belly, boston butt, and carcass trim did not change with slaughter weight (Table 4). Overall, the changes in wholesale cut percentages with SLW observed in this study were relatively small. A number of other studies have also found little effect of slaughter weight on primal cut distribution (Martin et al., 1981; Albar et al., 1990).
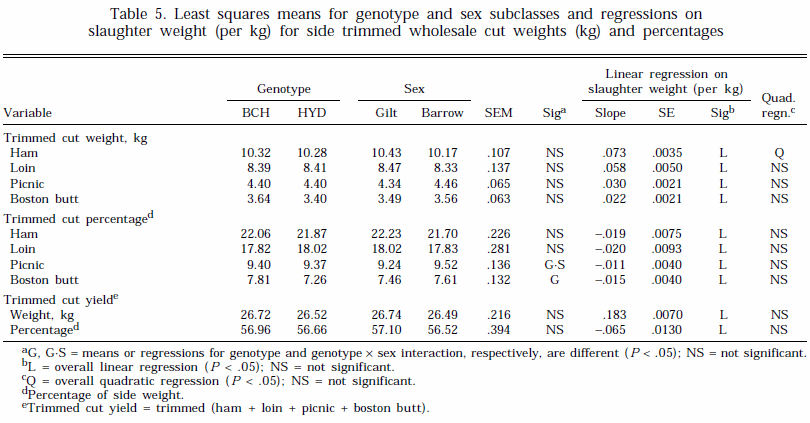
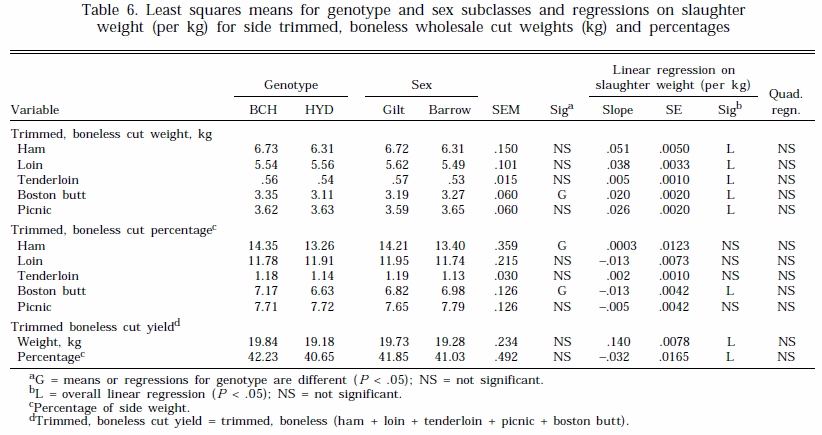
Treatment effects on weight and percentages of trimmed wholesale cuts and trimmed, boneless wholesale cuts are summarized in Tables 5 and 6, respectively. Differences between the genotypes and sexes for trimmed cut weights and percentages were generally small (Table 5). However, genotype BCH had a greater weight and percentage of trimmed, boneless boston butt and a higher percentage of trimmed, boneless ham than genotype HYD (Table 6). There were, however, no effects of genotype or sex on the total yields of trimmed or trimmed, boneless cuts (Tables 5 and 6). A number of studies have shown an advantage in lean cut yields for gilts compared with castrates (Fortin, 1980; Geri et al., 1990).
The weight of trimmed cuts and trimmed, boneless cuts increased linearly with slaughter weight (Tables 5 and 6). The regression of trimmed ham weight was related curvilinearly to SLW ( P < .05, Table 5). The percentage yields for trimmed cuts decreased with increasing slaughter weight (Tables 5). In addition, the percentage of trimmed, boneless boston also decreased with increasing weight, but there was no change in the percentage of the other trimmed, boneless cuts (Table 6). Overall, total side trimmed and trimmed, boneless cut weights increased by 1.8 and 1.4 kg, respectively, for each 10-kg increase in SLW weight. However, when expressed as a percentage of side weight, these changes were equivalent to decreases of .65 and .32 percentage units, respectively.
Carr et al. (1978) and Neely et al. (1979) also reported a small decrease in percentage trimmed, boneless yields in pigs slaughtered between 45 and 136 kg and 100 and 127 kg, respectively. Other workers have shown little change in percentage lean cut yield with slaughter weight (Fortin, 1980; Martin et al., 1981). The results of this experiment, in which pigs were taken to heavier weights than in other studies, suggests that reductions in percentage yields of the shoulder, loin, and ham with SLW will be relatively small.
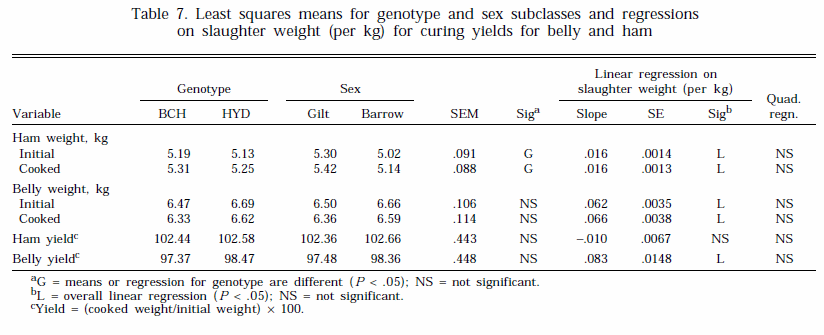
Curing yields for the ham and belly are presented in Table 7. There were no differences between the genotypes or sexes for ham or belly yield. In addition, there was no effect of slaughter weight on ham yields (Table 7). However, belly yields increased ( P < .001) with slaughter weight; the increase was equivalent to .83 percentage units increase in yield for each 10-kg increase in slaughter weight. Fredeen (1980) suggested that carcass weight was the major determinant of belly curing yields and that increases in yield with slaughter weight were primarily the result of an increase in the fat content of the belly. The range of meat quality measurements that were taken on the longissimus thoracis et lumborum muscle are presented in Table 8. Subjective color, firmness, and marbling scores were higher ( P < .05) for barrows, indicating that they had darker, firmer muscle with more visible marbling compared to that of gilts. There were no differences between the sexes for any of the other muscle quality traits measured. Most other studies that have compared muscle quality in barrows and gilts have shown little difference between them (Barton-Gade, 1987). There was an effect of genotype on 45-min pH ( P < .01); HYD had higher values. However, there were no genotype effects for any other trait (Table 8). Increasing slaughter weight was associated with reductions in longissimus thoracis color ( P < .05) and firmness ( P < .01) scores and lower 24-h pH ( P < .05), tenderness scores ( P < .05), and moisture content ( P < .001) in the longissimus lumborum. However, longissimus thoracis drip loss and longissimus lumborum fat content increased (P < .05). The increase in drip loss was equivalent to around .3% for a 10-kg increase in slaughter weight, a change that would result in a significant increase in commercial loss at higher weights. These results suggest that heavier pigs may be more prone to the development of the pale, soft, and exudative (PSE) condition. The development of PSE is normally associated with a rapid drop in muscle pH immediately postmortem. This, however, was not the case in this study, as evidenced by 45-min pH values, which did not change with slaughter weight. On the other hand, the development of PSE is temperature-dependent, and elevated muscle temperatures can also produce this condition (Fernandez et al., 1994; McCaw, 1994). One possibility is that muscle cooling rates are slower in heavier carcasses and, thus, muscle temperatures remain high in the early postmortem period, leading to an increase in the incidence of PSE. Further work looking at muscle temperature changes in heavy carcasses immediately after slaughter is warranted. Other studies have generally observed limited effects of slaughter weight on meat quality parameters, including water-holding capacity (Shuler et al., 1970; Martin et al., 1981). Differences between studies in the changes in meat quality with increasing slaughter weight may reflect differences in genotypes and(or) pre- and post-slaughter handling procedures. These factors are known to influence meat quality and they may interact with slaughter weight.
The changes in longissimus lumborum composition with increasing slaughter weight noted here are consistent with the findings of Shuler et al. (1970), who reported that ether extract fat increased and moisture content decreased with slaughter weights between 45 and 113 kg. However, Martin et al. (1981) found no change in these two components with increasing slaughter weights between 73 and 137 kg. Meat tenderness, as determined by a trained taste panel, decreased with slaughter weight in the present study (Table 8), which is in agreement with the findings of Ellis et al. (unpublished data), who showed an increase in taste panel assessed toughness and in shear force with increases in slaughter weight from 80 to 120 kg. However, in the present study, there was no change ( P = .18) in shear force values with weight and the change in taste panel scores with weight ( -.015 units/kg, P < .02) was relatively small. Overall, the results of this study suggest limited changes are observed in most muscle quality parameters with increased slaughter weight.
Implications
This study evaluated slaughter weights in excess of current commercial levels in two genotypes with medium lean growth potential reared on a single diet and under space allowances in excess of those used in commercial practice. The results suggest limited effects of slaughter weight on growth rate, feed efficiency, and cutting and curing yields. The tendency for meat quality to deteriorate with slaughter weight noted in this study, particularly in terms of drip loss, is of concern and merits further research.
Literature Cited
Albar, J., P. Latimier, and R. Grainier. 1990. Poids d’abbatage: Evolution des performances d’engraissement et de la carcasse de porcs abattus au dela` de 100 kg. J. Rech. Porc. Fr. 22:119.
American Meat Science Association. 1978. Guidelines for cookery and sensory evaluation of meat. American Meat Science Association— National Live Stock and Meat Board, Chicago, IL. AOAC. 1984. Official Methods of Analysis (14th Ed.). Association of Official Analytical Chemists, Arlington, VA.
Barton-Gade, P. 1987. Meat and fat quality in boars, castrates and gilts. Livest. Prod. Sci. 16:187.
Boland, M. A., A. P. Schinkel, and P. V. Preckel. 1993. Optimal slaughter weights and carcass value for three genotypes of hogs. Prof. Anim. Sci. 9:163.
Carr, T. R., L. E. Walters, and J. V. Whiteman. 1978. Carcass composition changes in growing and finishing swine. J. Anim. Sci. 47:615.
Fernandez, X., A. Forslid, and E. Tornberg. 1994. The effect of high post-mortem temperature on the development of pale, soft and exudative pork: Interaction with ultimate pH. Meat Sci. 37:133.
Fortin, A. 1980. The effect of slaughter weight on the carcass characteristics of Yorkshire barrows and gilts. Can. J. Anim. Sci. 60:265.
Fredeen, H. T. 1980. Yields and dimensions of pork bellies in relation to carcass measurements. J. Anim. Sci. 51:59.
Friesen, K. G., J. L. Nelssen, J. A. Unruh, R. D. Goodband, and M. D. Tokach. 1994. Effects of the interrelationship between genotype, sex, and dietary lysine on growth performance and carcass composition in finishing pigs fed to either 104 or 127 kilograms. J. Anim. Sci. 72:946.
Geri, G., O. Franci, B. M. Poli, G. Campodoni, and A. Zappa. 1990. Relationships between adipose tissue characteristics of newborn pigs and subsequent performance: II. Carcass traits at 95 and 145 kilograms live weight. J. Anim. Sci. 68:1929.
Gu, Y., A. P. Schinckel, and T. G. Martin. 1992. Growth, development, and carcass composition in five genotypes of swine. J. Anim. Sci. 70:1719.
Hahn, J. D. 1994. Lysine levels and ideal ratios of amino acids for finishing pigs. Ph.D. Dissertation. University of Illinois, Urbana, IL.
Hansson, I. 1974. Effect of sex and weight on growth, feed efficiency and carcass characteristics of pigs 1. Growth rate and feed efficiency of boars, barrows and gilts. Swed. J. Agric. Res. 4: 209.
Kanis, E., G. H. Nieuwhof, K. H. de Greef, W. van der Hel, M.W.A. Verstegen, J. Huisman, and P. van der Wal. 1990. Effect of recombinant porcine somatotropin on growth and carcass quality in growing pigs: Interactions with genotype, gender and slaughter weight. J. Anim. Sci. 68:1193.
Martin, A. H., H. T. Fredeen, G. M. Weiss, A. Fortin, and S. Sim. 1981. Yield of trimmed pork products in relation to weight and backfat thickness of the carcass. Can. J. Anim. Sci. 61:299.
McCaw, J. 1994. The effect of muscle condition and temperature on the color and water holding capacity of fresh pork. MS Thesis. University of Illinois, Urbana.
Morgan, J. B., G. C. Smith, J. Cannon, F. K. McKeith, and J. Heavner. 1994. Pork chain quality audit: Pork distribution channel audit report. National Pork Producers Council, Des Moines, IA.
NAMP. 1992. The Meat Buyers Guide (3rd printing). National Association of Meat Purveyors, Reston, VA. NCR-89 Committee on Confinement Management of Swine. 1993. Space requirements of barrows and gilts penned together from 54 to 113 kilograms. J. Anim. Sci. 71:1088.
Neely, J. D., R. K. Johnson, and L. E. Walters. 1979. Efficiency of gains and carcass characteristics of swine of two degrees of fatness slaughtered at three weights. J. Anim. Sci. 48:1049.
Neter, J., W. Wasserman, and M. H. Kutner. 1985. Applied Linear Statistics: Regression, Analysis of Variance and Experimental Designs. Richard D. Irwin, Homewood, IL. Novakofski, J., S. Park, P. J. Bechtel, and F. K. McKeith. 1989. Composition of cooked pork chops. Effect of removing subcutaneous fat before cooking. J. Food Sci. 54:15.
NPPC. 1991. Procedures to Evaluate Market Hogs (3rd Ed.). National Pork Producers Council, Des Moines, IA.
NRC. 1988. Nutrient Requirements of Swine (9th Ed.). National Academy Press, Washington, DC.
SAS. 1985. SAS User’s Guide: Statistics. SAS Inst. Inc., Cary, NC. Shuler, R. O., T. D. Pate, R. W. Mandingo, and L. E. Lucas. 1970. Influence of confinement, floor structure and slaughter weight on pork carcass characteristics. J. Anim. Sci. 31:31.
Tess, M. W., G. E. Dickerson, J. A. Nienaber, and C. L. Ferrell. 1986. Growth, development, and body composition in three genetic stocks of swine. J. Anim. Sci. 62:968