Introduction
For economic reasons, the pig industry is marked today by a phenotypic inbreeding for production parameters that might result in a genotypic linkage with an altered (compromised) immune responsiveness. A well-developed immune system and optimal immune responsiveness remain important for the welfare and productivity. Indeed these qualities can only be obtained if the health status of the animal scores high. Therefore a lot of energy and money is invested in prophylactic measures such as vaccinations and chemoprophylaxis against infectious diseases. However, health and immune responsiveness is not only maintained and improved by vaccinations and hygiene but also by an adequate supply of nutritional components to the animal.
In order to understand and improve prophylaxis by vaccination, it is imperative that we understand as much as possible the induction of an immune response by vaccination and the important communication (cell talk) between cells, such as antigen-presentation and cytokine production. A good comprehension of what is happening at the induction site of an immune response will help us to understand the difference between the use of live or dead vaccines as well as the interference of active (from a previous infection or vaccination) or passive immunity on the vaccination.
The induction of an immune response
1. The immune orchestra
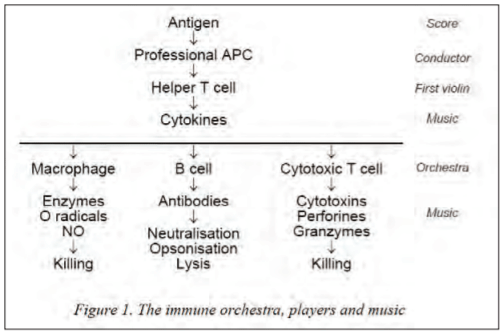
An immune response is like music, produced by an orchestra (the immune-competent cells) under the guidance of a conductor (the professional antigen-presenting cell such as the interdigitating dendritic cell, IDC) who interprets the score (antigen) (figure 1). The immune responses can be divided in innate immunity, responsible for the first line of defense and induction of immune responses, and acquired immunity, responsible for antigen-specific memory. Innate immunity is mediated by the “so-called” antigen a-specific immune cells such as the IDC (the conductor), macrophages and neutrophils (microphages). However, their important immune role is not restricted to a first-line defense but lays also in the triggering of alarm signals (interpretation of the score and direction of the music) which are crucial for initiating and directing subsequent antigen a-specific and specific defense reactions (the music). Moreover, recent data have shown that these “so-called” a-specific cells are not so a-specific as they recognize and differentiate specific pathogen-associated molecular patterns (PAMPs) with pathogen recognition receptors, such as Toll-like receptors (TLRs). Depending on the PAMPs recognized, the APC can direct the immune response into opposite ways with important implications on the inflammatory response, production and protection (see later).
The antigen-specific cells (members of the orchestra) which have an immunological memory and secrete immunoactive molecules (the music), are divided in two main populations, the B and T cells (figure 1). Their antigen specificity is mediated by cell surface receptors (their instruments), the B cell receptor (membrane-bound immunoglobulin, BCR) and the T cell receptor (TCR), respectively. The repertoire of each of these receptors for their different antigens is clonally distributed, i.e. each individual T cell and B cell has only one idiotype set of antigen receptors and can thus only recognize one specific determinant or epitope of the antigen. BCR recognizes directly a specific natural epitope present on unprocessed antigen while TCR recognize epitopes of a processed antigen presented on MHC molecules of an antigen-presenting cell (conductor). Indeed, the antigen is processed intracellular (interpretation of the score) by an antigen-presenting cell (APC) into short peptides: linear epitopes of 8 to 20 amino acids long) that associate intracellular with major histocompatibility complex (MHC) molecules. These MHC molecules will, on subsequent expression on the cell membrane of the antigen-presenting cell, present the epitope to the T cell.
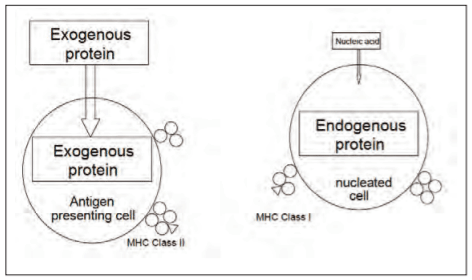
The T cell population is divided in CD4+ and CD8+ T cells (in swine there exist also double positive and double negative T cells) depending on the class of antigen-presenting MHC molecule they recognise. CD4+ T cells recognize their epitope in association with MHC class II molecules while CD8+ T cells recognize their epitope in association with MHC class I molecules. MHC class II molecules present epitopes from exogenous antigens, i.e. phagosomal antigens after uptake and processing by the APC (IDC, macrophage or B cell), while MHC class I molecules present epitopes from endogenous antigens, i.e. proteasomal antigens which have been transcribed inside the antigen-presenting cell as is the case for any virus-infected cell (figure 2). This difference has very important consequences as CD4+ T cells (or T helper cells, Th) compensate their antigen-presenting target cells with beneficial cytokines, while CD8+ T cells kill their infected target cells with cytotoxic factors and signal transductions.
Figure 2. Presentation of processed antigens (peptides: T cell epitopes) of exogenous and endogenous antigens in association with MHC class II and MHC class I molecules, respectively
Recently however, it has been demonstrated that in dendritic cells soluble antigens can be redirected during phagocytosis towards MHC class I presentation which has been named “cross-presentation”. For this cross-presentation, there appears to be a delay in the maturation of endosomes towards lysosomes under the control of NADPH oxidase-associated reactive oxygen radicals and endosomal antigen release by lipid peroxidation, enabling presentation by class I. Based on their cytokine-producing profiles, CD4+ T cells can be subdivided in different subtypes: the inflammatory T helper cell or Th1 is characterized by the secretion of interleukin-2 (IL-2), interferon- (IFN- ) and IL-12 while the non-inflammatory T helper cell or Th2 is characterized by IL-4, IL-5, IL-10 and IL-13. More recently other T helper cells have been identified such as the antibacterial Th17 responsible for the attraction of neutrophils and the regulatory T cell (Treg) responsible for the production of immune-suppressive cytokines such as transforming growth factor (TGF- ). The kind of CD4+ T cell that will be induced, is determined by the APC (IDC the conductor), the strength of stimulus and amount of antigen, and the kind of cytokine(s) produced by the environmental cells, all of which will have important modulatory implications on the type of resultant immune responses (music).
2. The concert hall: secondary lymphoid structures, the place of induction of B and T cells
Primary induction of immune responses occurs in the secondary lymphoid organs. These structures are filters for antigen (or antigen bearing APC) and naive lymphocytes and are optimally organized for intercellular communication between antigen-presenting cells, T cells and B cells. These organs are strategically placed on the lymph (lymph nodes) and blood circulation (spleen) to capture antigen and antigen-presenting cells draining from the tissues. By specific adhesion molecules, naive lymphocytes are directed to emigrate from the blood circulation into these secondary lymphoid organs in search for their specific antigen.
Conversely to other mammals, the draining lymph nodes of pigs appear rather an agglomerate of a few lymph nodes which are stuck together in an inverted position with the medulla being external to the cortex. Nevertheless, the physiology of the T and B cell areas are broadly conventional, with the T cell zone located in the paracortex and the B cell zone with its follicles located in the cortex. T and B cells enter via paracortical postcapillary venules, of which many are high endothelial venules (HEV). The major difference between pigs and other mammals lies in the emigration route of the lymphocytes: instead of leaving by sinuses through the medulla in the afferent lymph, they emigrate directly into the blood through the HEV. As a consequence, efferent lymph of pigs contains very few lymphoid cells as compared to other species.
The porcine spleen is composed of white and red pulp. The white pulp contains mainly leukocytes while the red pulp consists of erythrocytes and lymphocytes. In the white pulp lymphocytes are organised around the arterioles in periarteriolar lymphoid sheets (PALS). The peri-arteriolar sheet consists of a mantel of T cells (CD4+> CD8+) surrounded by a marginal zone containing macrophages and B cells. T cells are also found in the red pulp (CD8+> CD4+). These cells are already present at birth, but a large increase occurs after 2 weeks of life. At the same time, a similar increase occurs in B cells, mainly IgM+, with the formation of small follicles in the PALS as a consequence. In 4-week-old pigs the percentage of B cells in regard to the total amount of lymphoid cells, is similar to that in peripheral blood, approximately 35%. Most IgM-producing cells are located in the red pulp, whereas IgG- and IgA-producing cells are predominantly present in the periphery of the PALS. At 10 months of age however, IgG and IgA-producing cells can also be found in the red pulp.
3. Immunological defence of the gut and the gut-associated lymphoid tissue (GALT)
The immune defence system of the gut consists of lymphoid tissues and cells distributed along the gastrointestinal tract (reviewed in Brandtzaeg and Pabst, 2004). The lymphoid tissue localized along the gastrointestinal tract constitutes quantitatively the major part of the immune system of the whole body. This extremely developed gastrointestinal immune system reflects the importance of the mucosal immune defence system against the continuous attack of antigens and pathogens. Moreover, the major development of this local immune tissue as well as the individual immune reactivity seems to be induced by the continuous contact of the gastro-intestinal mucosa with the gastrointestinal flora, as well with pathogens, as evidenced by the atrophic mucosal immune system in axenic (germ-free) animals.
Some important features characterize the mucosal immune system:
- it possesses mucosa-associated lymphoid tissue (MALT or GALT for the gut-associated lymphoid tissue) as well as local and regional draining lymph nodes where the induction of immune responses is established, such as the Peyer’s patches (PP) and the mesenterial lymph nodes, respectively,
- certain subpopulations of lymphoid cells predominate at the mucosal surfaces,
- there is a specific recirculation of mucosal lymphocytes towards mucosae, known as mucosal homing, and
- the predominant mucosal immunoglobulin is dimeric IgA which is secreted at the mucosal surface.
All these elements of the mucosal immune system are working together to generate immune responses that protect the host against mucosal invaders but also render the host tolerant against ubiquitous dietary antigens and the beneficial microbial flora of the mucosae. The elucidation of the mechanisms that determine immune tolerance or immune induction forms today a major topic of study where the type of activation state of the antigen-presenting IDC will determine whether T cells will or will not differentiate in T regulatory cells playing a role in immunotolerance.
The gastrointestinal immune system consists of lymphoid cells in organised sites like PP and mesenterial lymph nodes, and lymphocytes spread over the stromal tissues in the lamina propria and the epithelium (the intra-epithelial lymphocytes, IEL) of the intestine. There exists evidence that these organized structures are the places (or one of the places) where antigen enters the mucosal immune system to initiate subsequently the immune reactions: therefore these places are quite often called the inductive lymphoid sites of the mucosal immune system. Lymphocytes located in the lamina propria and between the epithelial cells of the intestinal tract are attributed with effector functions, such as antibody production, cytokine production and cytotoxicity, and these places are therefore referred to as the effector sites of the intestinal tract. These inductive and effector sites are interconnected by selective migration of lymphocytes (homing) whereby cells that have been activated in the inductive sites migrate specifically to effector sites of the intestinal tract. This interconnection assures that mucosal responses are primarily directed and localised against antigens that have been recognized at mucosal surfaces.
4. The overture: the acute phase response and the arachidonic acid cycle
Inflammation is the answer of tissue to irritation, injury and infection, and is quite often needed for the induction of strong immune responses. It consists of a complex cascade of nonspecific events, known as the acute-phase response, which confer early protection by limiting tissue injury to the place of infection or destruction. One important function of this reaction is to recruit more phagocytic cells to the site of injury. Moreover, it initiates the specific immune response against the invader. The localized reaction is induced by clotting factors and pro-inflammatory cytokines released by the activated resident sentinels, the tissue macrophages. The combined actions of their pro-inflammatory cytokines IL-1, IL-6, IL-12 and TNF- and the release of chemokines by activated macrophages and activated structural tissue cells (keratinocytes, fibroblasts, endothelial and epithelial cells) are responsible for changes in the surrounding capillaries, inducing an influx of neutrophils, monocytes and effector lymphocytes into the site of inflammation. Indeed, there is an increased expression of inflammatory adhesion molecules on the endothelial cells, which trap the circulating leukocytes with subsequent diapedesis and migration towards the place of tissue injury in order to fulfill their duties. However, activated phagocytes release also other proteins with potent local effects, such as toxic radicals, peroxides, nitric oxide, plasminogen activator and enzymes. Phospholipase A2 cleaves the fatty acid arachidonic acid (C20:4n6) from the glycerol backbone of membrane-bound phospholipids. The liberated arachidonic acid can then undergo controlled oxidative metabolism to form a variety of eicosanoids (C20) with different physiological and immune effects.
The cyclooxygenase pathway yields prostaglandins (PG), prostacyclin and thromboxanes while the lipoxygenase pathway the leukotrienes (LT) and lipoxins. The eicosanoids act as autocrine/paracrine regulators, since most of their biological effects are limited to the site of biosynthesis. LTB4 stimulates chemotaxis of neutrophils and an increased expression of their C3b receptors while LTC4, LTD4 and LTE4 are collectively known as the slow reactive substance of anaphylaxis (SRS-A), being more than 1000 times more potent than histamine in smooth muscle contraction and vasodilatation. The latter is important in view of the increased blood flow in inflamed tissues inducing an even higher influx of leukocytes into the inflammation site. After exerting their effect, eicosanoids are rapidly catabolized to inactive compounds in the liver and lungs.
Vaccination: induction of a specific protective immunological memory
1. Live versus dead (inactivated, non-proliferating) vaccine
The absolute difference between live and dead vaccines lays in the fact that the vaccine dose of a dead vaccine required for successful immunization was made outside the host (the lab), while it is made inside the host for live vaccines. In other words, if a live vaccine fails to proliferate inside the host and fail to produce enough antigen to reach its immune dose, no immune induction or vaccination will occur.
The immunologically relevant difference however, between dead and live vaccines lays in the fact that a dead vaccine delivers exogenous antigens while a live vaccine delivers exogenous as well as endogenous antigens (figure 3). Although both vaccines induce good B cell (antibody) protective responses, dead vaccines induce only CD4 T cells while live vaccines induce CD8 as well as CD4 T cells. In view of intracellular pathogens, dead vaccines exert their effect on the infected cell indirectly by secreted cytokines while live vaccines do that by secreted cytokines and direct cell-mediated CD8 killing of the infected cell.
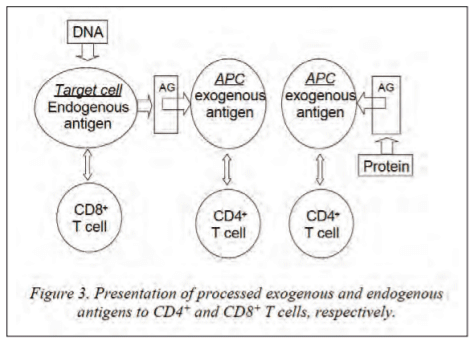
Another immunological relevant difference is that live vaccines can be administered systemically as well as mucosally while dead vaccines can for the moment only be administered systemically by local injection. As a live vaccine is delivered locally or mucosally, it will migrate to its predilection site and cells for proliferation, where it will induce a mucosal and systemic protective response. Conversely a dead vaccine will only induce a systemic response. However, major efforts in recent research are directed towards the mucosal delivery of dead vaccines or modulation of immune responses towards mucosae by systemic delivery of death vaccines. The last twenty years a lot of efforts have been directed towards oral delivery of antigens (in the feed) in order to immunize and protect animals mucosally at the gastrointestinal tract with dead (inactivated, non-proliferating) vaccines. The major challenge here is to avoid induction of antigen-specific tolerance which is a major mechanism operative against the multitude of antigens encountered at the gastrointestinal epithelium. One of the most promising ways of such oral antigen delivery for immunization is the receptor-mediated uptake at the gastro-intestinal tract which is and has thoroughly been researched, reviewed and discussed by our group (Devriendt et al. 2012). This mechanism is based on the uptake of molecules by receptors/ligands present on the intestinal epithelium (inclusive M cells of the PP) for lectins, immunoglobulins and even for some pathogen antigens and other molecules. The antigens of interest for immunization can then be recombinantly of covalently coupled to these molecules for receptor-mediated uptake in the body. Such an optimised antigen uptake system opens ways for oral immunization via animal feed, where plants are then recombinantly transformed for production of the antigens of interest in leaves or seeds for inclusion in the feed. We tested already a recombinant system in plants for passive protection however, by specific expression of sIgA antibodies specific for F4 in seeds and inclusion of the latter in porcine feed with successful protection against weaning diarrhoea (Vikram et al. 2014).
2. Adjuvants
The generation of a successful immune response upon vaccination is dependent on several factors. We could summarize these factors in two categories, namely the antigen delivery, amount and presentation and secondly the creation of the correct immunological environment (inflammatory) or overture for inducing immune responses.
With respect to dead vaccines it imperative that the antigen is given over a prolonged period in the correct three-dimensional configuration (at least for correct and high affinity antibody formation) in order to have a correct and long-lasting antigen delivery and recognition. Therefore dead vaccines are delivered in precipitated forms or oil/water emulsions to avoid quick clearance from the body and to allow slow release of the antigen to the immune system. In order to get a good environment for immune induction, i.e. an inflammatory environment, inflammatory products are added in order to activate the innate immune system and antigen-presenting cells and increase the chemotaxis of immune-competent cells at the site of antigen delivery and immune induction (draining lymph node or spleen). Therefore adjuvants contain quite often ligands or pathogen-associated molecular patterns (PAMPs) which activate antigen-presenting cells as well as lymphocytes by binding to their pathogen recognition receptors such as Toll-like receptors, NOD receptors, RXR and c-type lectin receptors which belong to the innate immune system and converse with the acquired immunity to generate humoral and cell mediated immune responses.
In order to get maximal uptake of antigen by the antigen-presenting cells or the antigen-specific cells or to enable cross-linking of the antigen-specific receptors and as a consequence better activation of the latter, antigens are formulated in multimeric forms such as dendromers, iscoms, viroids, micells or liposomes.
A lot of efforts have also been focussed on the direction of immune responses of systemically delivered antigens towards the mucosae by selection and inclusion of specific adjuvants in immunization formulations (Cox et al. 2006).
With respect to live vaccines, the major point is that the vaccine is and stays alive before and upon administration allowing production of the required amounts of antigens for inducing the immune response inside the host and preferably at the entry and predilection site of the pathogen as the live vaccine (mostly attenuated pathogen) is migrating towards its preferred cells in that place. Antigens produced from live vaccines are normally in their correct tridimensional structure as well as in their multimeric form. When constructing live vaccines, immune-stimulating molecules can be introduced genetically by inserting coding genes or immune-stimulating nucleotide sequences (PAMPs) into the attenuated pathogen or live vector.
3. Maternal interference
One of the major problems in successful vaccination is the interference with maternal immunity or active immunity from previous vaccinations. Previous vaccinations or passive transfer of maternal antibodies via colostrum or milk can have a blocking effect on subsequent vaccination.
Indeed vaccination wants to induce or reactivate B cells and T cells via their membrane antigen-specific receptors, i.e. BCR and TCR respectively. When there are already secreted antigen-specific BCR i.e. antibodies (whether passive or active) circulating in the host, they can compete with the specific membrane BCR on B cells and as a consequence block the activation of the latter as the antigen epitopes are already opsonised by the circulating antibodies and cannot interact with the specific BCR for (re)activation of the B cells. As colostrogenic antibodies have been taking up in the systemic circulation while lactogenic antibodies remain in and protect the gastrointestinal tract, interference on systemic vaccination is mainly expected from colostrogenic antibodies.
Here again we have to make a difference between vaccination with live or dead vaccine. As the immunizing amount of antigen in dead vaccines has already been produced outside the host, circulating antibodies depending on their concentration, will only neutralise a certain part of the antigens. This can have an influence on the activation of B cells but less on T cells as they are reacting with T cell epitopes presented by antigen-processing cells and have not been neutralised by antibodies on B cell epitopes. On the contrary opsonised antigens might be favoured for uptake by antigen-presenting cells. Conversely, live vaccines have to produce their immunizing antigen amount inside the host and circulating antibodies can hinder or abolish that production by neutralising (killing) the live vaccine so that immune induction never happens, in other words it becomes a dead vaccine but containing much too low amounts of antigen to induce immune responses.
Another way to avoid interference of systemic passive immunity on vaccination, is mucosal instead of systemic administration of the live vaccine. As the colostrogenic antibody level is much lower at mucosal sites than in the system, and as the mucosal site is quite often the predilection site for multiplication and transcription of the antigens, one would expect less interference by mucosal vaccination. This is probably the case for respiratory vaccination but not for oral administration as the gastrointestinal tract is full of lactogenic antibodies which might neutralise the vaccine.
Depending on the kind of passive immunity you want in the offspring, i.e. a high colostrogenic immunity or a high lactogenic immunity, vaccination time spots of the mother animal can be varied from distant to close to parturition date: 3 weeks prepartum will rather give a colostral immunity with high concentrations of IgG in the colostrum, while vaccination closer to partus will give longer and higher presence of IgG and IgA in the milk, enabling longer protection of gastrointestinal tract.
4. Interference between simultaneous vaccinations
Simultaneous vaccinations against different pathogens with separate dead vaccines might interfere with each other when injected in the same place, indeed when the injected vaccines are drained to the same draining lymph node where the immune induction is happening. This is all dependent on the kind of immune responses the vaccines are intended to induce. It is know that Th1 and Th2 responses interfere with each other in the kind of immune response they induce by their secretion of opposing cytokines. As some adjuvants are added to direct immune responses towards rather humoral or cell-mediated immune responses, they can have opposing effects on their respective immune inductions. As these different factors present in the vaccines or produced during vaccination might interfere with each other, it is advisable to test first in vivo if they are or are not interfering with each other in the induction of their respective intended immune responses.
When the vaccines are however injected in different places drained by different lymph nodes, no interference is expected. The only interference which might happen would be that one vaccine is so inflammatory that most immunocompetent naïve cells are drawn towards that vaccine-induced inflammation site so that there is a decreased blood flow in the other vaccination site with reduced chance or slower timing to find the correct antigen-specific cells to be stimulated.
Conversely, administration of a single vaccine containing antigens from different pathogens is safe and should not interfere in the induction of protective responses against the different pathogens as this has (should have been) been tested in the clinical trials by the company: the adjuvant for inducing the type of T cell response has been selected on the basis of what type of protective responses are desired against the respective pathogens. However, it is still possible that certain molecules of specific pathogens act as PAMP and might have their own effect of the direction of the immune response.
Conclusion
In order to understand and improve vaccine choice or vaccination schedule, it is imperative to comprehend the music of immunology. Live vaccines are immunologically speaking the best as they can be delivered at the entry port of the pathogen (i.e. mucosae) where the latter is best stopped to disseminate into the body and secondly as they induce cell-mediated, inclusive CD8 responses, as well as humoral immunity. However, vaccinations with live vaccines are more sensitive to failure than those with dead vaccines, as they can fail in reproduction and production of their necessary antigenic load for immune induction. In conclusion, vaccinate animals with live vaccines when, or in places where no interference is expected and vaccinate with dead vaccines when passive or active immunity is already present and might interfere.
Presented at the 24th International Pig Veterinary Society Congress. For information on the next edition, click here.







.jpg&w=3840&q=75)