Feed additives and feed efficiency in the pork industry
Published: May 30, 2010
By: F.R. Dunshea1,2, T-Y. Hung 1,2, H. Akit1,2 and C.V. Rikard-Bell1,3
(1CRC for an Internationally Competitive Pork Industry; 2 Department of Agriculture and Food Systems, Melbourne School of Land and Environment, The University of Melbourne; 3 Murdoch University, Australia)
Summary
Although metabolic modifiers that require daily injection, such as porcine somatotropin, are available for the Australian pork industry, producers are interested in the potential of orally administered compounds to improve body composition, especially to reduce fat depth. The purpose of this review is to discuss dietary additives that can be used to manipulate growth, body composition or meat quality in growing pigs, viz., ractopamine, cysteamine, chromium, lecithin, betaine and dietary narcoleptics. Ractopamine increases lean tissue deposition in all sexes after 60 kg live weight and recent evidence suggests that 0.56 g and 0.60 g available lysine per MJ DE should be sufficient to maximize performance when 5 ppm of ractopamine is included in the diets of boars and gilts, respectively. Cysteamine may stimulate somatotropin secretion in pigs and under some circumstances may increase growth rate and decrease back fat depth. However, responses are variable and further work is needed to define the dose-response curve. Responses to dietary chromium are variable and this may in part be due to inefficient digestion and absorption of chromium. Micro- and nano-particles of chromium may increase the efficacy of dietary chromium and improve insulin sensitivity and growth performance in pigs. Dietary lecithin offers a means of reducing skeletal muscle collagen content and improving meat quality. It may also improve dressing percentage, but this effect needs to be verified. Dietary betaine has the potential to reduce maintenance requirements and, in some circumstances, the additional energy made available may be used to support protein deposition. Dietary narcoleptics offer a means of reducing sexual and aggressive behaviours in finisher pigs, specifically boars, and of improving growth performance. Responses to dietary feed additives are generally not as great as responses to porcine somatotropin, but they do offer an alternative means of manipulating growth and carcass quality.
Introduction
It has been known for almost 60 years that injection of growing pigs with pituitary tissue extracts containing porcine somatotropin (pST) increases protein deposition and decreases fat accretion (Turman and Andrews, 1955). Advances in biotechnology have now provided a means of producing pST on a commercial scale and the efficacy of daily injection of recombinant pST for improving the productive performance of swine is beyond doubt (Campbell et al., 1991). Porcine somatotropin has now been approved for use in the Australian pig industry as a daily injectable metabolic modifier (Dunshea et al., 2002; Dunshea, 2005). Because daily injection with pST is problematic, alternative regimens involving less frequent injections have been investigated and have been demonstrated to have similar efficacy to daily injections (Dunshea, 2002). Nonetheless, pork producers are still interested in orally administered compounds for improving body composition, particularly for reducing fat depth. The purpose of this review is to discuss dietary additives that can be used to manipulate growth, body composition or meat quality in growing pigs, viz., ractopamine, cysteamine, chromium, lecithin, betaine and dietary neuroleptics.
Ractopamine
Ractopamine is a β-agonist and has been shown to increase lean tissue deposition, although its effects on back fat are small and variable (Dunshea et al., 1993a,b;1998a) and responses may diminish with time due to downregulation of β-receptors (Dunshea et al., 1998b). The efficacy and mode of action of ractopamine has been much reviewed and the reader is referred to Dunshea (1993), Dunshea and Gannon (1995) and Mersmann (1998) for comprehensive discussions. However, since the early registration studies, there have been changes in management, genetics and slaughter weights as well as in pricing structures, which necessitated the commission of further studies by the Australian Pork Cooperative Research Centre (CRC) to ensure optimum exploitation of the benefits of ractopamine. These studies were focused on the effects of sex, live weight, dietary lysine, timing and dosage on the temporal response to ractopamine.
Rikard-Bell et al. (2009 and unpublished data) investigated interactions between starting weight (65, 80 and 95 kg) and dose of dietary ractopamine (0, 5, 10 and 20 ppm) in two separate experiments with boars and gilts. These studies clearly showed that most measures of productive performance and carcass composition, except feed intake and back fat depth, improved with increasing doses of ractopamine in all weight and sex classes. For lighter animals, average daily gain (ADG) was maximised at 5 ppm, whereas higher doses were required for heavier classes of pigs and for maximal responses in carcass weight and dressing percentage. This scenario may suit the Australian and New Zealand markets because pigs are marketed at light weights; thus only 5 ppm of dietary ractopamine is required to achieve maximum ADG responses in boars. However, greater doses of ractopamine are required for further improvements in carcass attributes.
Research conducted 20 years ago indicated that the lysine requirements of gilts fed 0 or 20 ppm ractopamine between 60 kg and 90 kg live weight were 0.65 g and 0.75 g available lysine per MJ DE, respectively (Dunshea et al., 1993a). There has been considerable genetic selection for lean growth since then as well as a general industry decision to use a lower inclusion rate of ractopamine than initially suggested. Furthermore, there were no data on the ractopamine requirements of boars. Rikard-Bell et al. (unpublished data) recently investigated the lysine requirements of boars and gilts fed 0, 5 or 10 ppm ractopamine from 65 kg live weight and found that 0.56 g of available lysine per MJ DE was sufficient to maximize ADG, feed conversion ratio (FCR) and carcass weight in gilts fed 0, 5 or 10 ppm of ractopamine. On the other hand, there were interactions between dietary ractopamine and lysine in boars such that 0.56 g and 0.60 g of available lysine per MJ DE were sufficient to maximize ADG and FCR in boars consuming diets containing 5 ppm and 10 ppm ractopamine, respectively, whereas there was no sign of a plateau in the control boars up to 0.72 g of available lysine per MJ DE. These data suggest that the ractopamine-treated pigs were more efficient at utilizing lysine and other amino acids than the control boars, perhaps through a reduced protein turnover rate.
Cysteamine
The release of endogenous pST from the pituitary is regulated by several secretagogues and by inhibitors such as somatostatin. Several researchers have attempted to increase pST secretion by immunizing against somatostatin, but results have been inconsistent, possibly because of low antibody titres (Dubreuil et al., 1989; McCauley et al., 1995). Others have reported that the sulfhydryl compound, cysteamine hydrochloride (CSH), increases somatotropin secretion in rats (Tannenbaum et al., 1990), sheep (McLeod et al., 1995a,b) and fish (Xiao and Lin, 2003), presumably through inhibition of somatostatin secretion (Wakabayashi et al., 1985). A few recent studies have investigated the effects of CSH on growth performance in pigs. Zhou et al. (2005) fed CSH (0.6 g/kg diet) to finisher pigs for 5 weeks and reported that it increased ADG (+4.8%) and decreased back fat depth (–20%). Dunshea (2007) found that dietary CSH (0.7 g/kg diet) increased ADG (+12%) and decreased FCR (–9.4%) over the first 2 weeks of supplementation. Although the response was not as pronounced over the entire 5-week supplementation period, nevertheless dietary CSH supplementation resulted in a modest increase in ADG (+7.4%). As a result, final live weight (+2.0 kg) and carcass weight (+1.6 kg) were increased and back fat depth was decreased (–1.0 mm) by CSH. Yang et al. (2005) found that daily gain was increased (+13.7%) by a low dose of CSH (0.03 g/kg feed) but not by a higher dose of CSH (0.05 g/kg feed); these doses are very low compared with those used in other studies. Back fat depth at the P2 site was reduced in a dosedependent manner by dietary CSH supplementation (15.7, 12.7 and 10.4 mm for pigs fed 0, 0.03 and 0.05 g/kg CSH, respectively). Liu et al. (2009) found that dietary CSH (0.07 g/kg) increased ADG (+19%), feed intake (+15%) and nitrogen retention (+63%) but did not change FCR. In a recent study conducted within the Pork CRC, dietary CSH (0.7 mg/kg) had no effect on growth performance but decreased P2 back fat depth (–1.9 mm) (David Miller, personal communication). Cysteamine had no effect on serum somatotropin level but decreased ghrelin level after 3 weeks of treatment. The inhibitory effect of CSH on ghrelin may be a feedback mechanism and may partially explain the apparent transitory nature of the growth response (Dunshea, 2007). An area of research that needs to be investigated is the dose response to CSH, as the dose used in studies reported above varies 10-fold and downregulation may be more rapid at higher doses.
Chromium
Cr is an essential element for humans and animals (Mertz, 1969; 1993), trivalent Cr3+ being the most stable form. Cr functions as a cofactor for insulin and enhances the ability of insulin to regulate glucose, protein and fat metabolism (Amoikon et al., 1995). However, Cr3+ is normally poorly absorbed and utilized. The digestibility of the inorganic and organic forms of Cr is 0.5–2% and 10–25%, respectively (Mertz, 1969; Underwood, 1977). Currently, there is no recommendation for dietary Cr supplementation (NRC, 1998) for pigs and, as most pig diets are primarily formulated using ingredients of plant origin, which are usually low in Cr content (Giri et al., 1990), it is possible that many pig diets have a low Cr content.
It has been suggested that dietary Cr increases insulin sensitivity in pigs (Steele et al., 1977; Matthews et al., 2001b) and over the past three decades, Cr has been investigated as a potential means of manipulating fat deposition in humans and farm animals. Although chromium chloride has been successful in improving the growth rate of turkeys (Steele and Rosbrough, 1979) and protein-deficient rats (Mertz and Roginski, 1969), effects in pigs have been equivocal (Page et al., 1993; Mooney and Cromwell, 1995; Mooney and Cromwell, 1997; Zhang et al., 2010). Page et al. (1993) conducted three experiments to determine the efficacy of Cr chloride, Cr picolinate and picolinate alone for grower-finisher pigs. Although Cr and picolinate had no effects on growth rate or FCR, Cr picolinate had dose-dependent effects on loin-eye area and back-fat depth. Others have also observed little or no effect of Cr picolinate on ADG and FCR (Evock-Clover et al., 1993; Mooney and Cromwell, 1995; Kim et al., 2009). Zhang et al. (2010) observed no effect of dietary Cr picolinate in grower pigs but observed a small increase in ADG in finisher pigs. Lean tissue deposition /loin-eye area was increased and fat deposition/back-fat depth was decreased by Cr picolinate in some studies (Page et al., 1993; Jackson et al., 2009) but not in others (Evock- Clover et al., 1993; Matthews et al., 2001c).
At least part of the variation in response to Cr may be related to the low and variable digestion, absorption and availability of Cr, which could be improved by using nano- or micro-sized Cr. For example, in rodents, the efficiency of uptake of 100 nm particles by the intestinal tissue was 15- to 250-fold higher compared with large micro-particles (Desai et al., 1996). The digestibility of nano-Cr picolinate was threefold higher than that of native Cr picolinate in rats (Lien et al., 2009). Recently, we conducted a meta-analysis of seven studies conducted on nano-Cr during the past 6 years and found that ADG and FCR were improved by 7% and 4%, respectively (Hung et al., 2010). Pork CRC scientists have also conducted research on dietary nano- and micro-Cr to determine their efficacy and mechanisms of action.
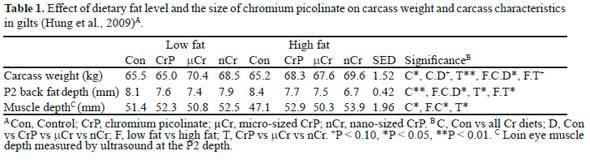
Hung et al. (2009) investigated interactions between dietary fat (2% vs 6% fat) and the form of dietary Cr picolinate (400 ppb of normal, micro- or nano-sized particles) for 6 weeks in finisher gilts. Nano- and micro- Cr was produced using the dry polish method in a dry cryo-nanonization grinding system integrated with a size separator. Briefly, the particle size of Cr picolinate was reduced using a grinder and it was then passed through appropriately sized sieve end-plates to collect the nano- and micro-sized particles. No solvent was used and the temperature was restricted to less than 40 °C during milling.
Over the first 21 days of the experiment, there was a significant main effect of Cr on ADG (944 vs 1011 g/d, respectively, P = 0.021) but not of Cr size (P = 0.17). Dietary Cr increased carcass weight and loin muscle depth and responses were greatest for nano-Cr (Table 1). Dietary Cr also decreased P2 back fat depth and the greatest response was observed in pigs fed nano-Cr and a high-fat diet. Furthermore, dietary Cr decreased plasma insulin level (7.0 vs 5.1 mU/mL, P = 0.04) without changing plasma glucose level (3.6 vs 3.5 mmol/L, P = 0.60) and increased muscle phosphatidylinositol 3-kinase and Akt mRNA expression, which are indicative of an improvement in insulin sensitivity in muscle through enhanced signalling downstream of the insulin receptor. A more recent Pork CRC study conducted under commercial conditions with very lean gilts found that final live weight was increased by 400 ppb of micro-Cr picolinate (94.0 vs 95.4 kg, P = 0.09) but back fat depth did not change (8.0 vs 8.0 mm, P = 0.94). These data suggest that dietary Cr picolinate increases ADG and improves carcass traits in lean genotypes and that responses are most pronounced in pigs fed a high fat diet containing micro-Cr or nano- Cr. Further studies on dose-response curves are being conducted to determine the effects of low nano- and micro-Cr picolinate inclusion rates.
Lecithin
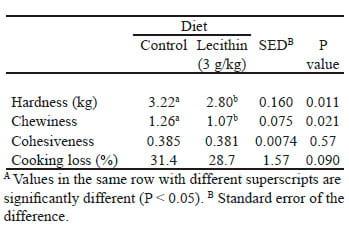
Tenderness and texture are considered the most important organoleptic or sensory attributes of pork by consumers and are affected by the connective tissue, myofibril and sarcoplasmic protein components of meat (Lawrie, 1998). Collagen, a major component of intramuscular connective tissue, is a key determinant of the tenderness of meat (Light et al., 1985). The stability of collagen is dependent on the cross-linking of collagen fibrils (Rowe, 1981) and the degree of cross-linking of collagen is dependent on the amount of hydroxyproline present in collagen. The enzyme responsible for the hydroxylation of proline is prolyl- 4-hydroxylase (Bailey and Light, 1989). It has been hypothesized that polyenylphosphatidylcholine (PPC), a phospholipid present in lecithin extracted from soya bean, may decrease the cross-linking of collagen fibrils. Several studies have shown that the primary mode of action of PPC on collagen involves inhibition of prolyl-4-hydroxylase, resulting in reduced collagen fibril cross-linking (Lieber, 1997, 1999). Lieber (1997, 1999) also found that PPC promotes the action of collagenase and increases the breakdown of collagen in non-human primates. Previous Australian research has shown that dietary lecithin decreases the compression characteristic of pork (D'Souza et al., 2005; Edmunds et al., 2005) (Table 2), possibly by decreasing the amount of collagen or the extent of collagen crosslinking. Dietary lecithin supplementation may reduce cholesterol absorption and LDL cholesterol. The use of dietary lecithin supplementation to improve pork quality and the health benefits of pork represents a significant marketing opportunity for the industry (Bellhouse et al., 2010). This would be particularly appealing if carcass weights and characteristics could also be improved by dietary lecithin. Hence, the Pork CRC has commissioned research to investigate the effect of dietary lecithin on carcass characteristics and meat quality in pigs.
Table 2. The effect of dietary lecithin supplementation on some aspects of meat quality of individually housed female pigs (after D'Souza et al., 1995)A
The first study commissioned by the Pork CRC investigated the dose response to dietary lecithin under commercial conditions. Briefly, 36 gilts were randomly allocated at 55 kg live weight to individual pens in an experimental finisher shed for acclimatization one week before the beginning of the experiment, during which time they were fed the control diet at 80% of ad libitum intake. Subsequently, the pigs were fed a commercial finisher diet plus 0% (control diet), 0.4%, 2% or 8% lecithin. All pigs had ad libitum access to feed and water for 6 weeks. Dietary lecithin had no effect on shear force (3.1 vs 3.8, 3.4 and 3.3 kg for 0, 0.4, 2 and 8% lecithin, respectively) but increased dressing percentage (73.6 vs 75.5, 75.5 and 75.5% for 0, 0.4, 2 and 8% lecithin, respectively; P = 0.009). Dietary lecithin also made the pork more red and darker as indicated by a linear increase in a* (5.1 vs 5.3, 6.0 and 6.2 for 0, 0.4, 2 and 8% lecithin, respectively; P = 0.004) and a decrease in L* (51.1 vs 49.9, 49.4 and 49.1 for 0, 0.4, 2 and 8% lecithin, respectively; P = 0.048). These effects were associated with changes in the expression of genes involved in collagen synthesis and degradation.
Pork from pigs fed dietary lecithin had lower intramuscular fat than pork from the control pigs (2.1 vs 1.5, 1.6 and 1.8% for 0, 0.4, 2 and 8% lecithin, respectively; P = 0.07), although there was no significant effect on total cholesterol. Pork from pigs fed diets supplemented with lecithin had higher (P < 0.05) levels of polyunsaturated fatty acids (PUFAs), linoleic acid and α-linolenic acid compared with pork from pigs fed the control diet. Levels of long-chain PUFAs such as C20:2n-6 and C20:3n-3 were also higher in pork (P 0.05 for both) from pigs fed dietary lecithin compared with pork from pigs fed the control diet. There was a dose-dependent increase in the ratio of PUFA to saturated fatty acid in pork from pigs that consumed lecithin (0.46 vs 0.56, 0.60 and 0.73 for 0, 0.4, 2 and 8% lecithin, respectively; P < 0.001).
The effect of dietary lecithin on dressing percentage (+2 percentage units) was initially unanticipated but, if proven correct, could improve feed efficiency. The study of D'Souza et al. (2005) did not indicate any effect of lecithin on dressing percentage. However, further interrogation of data from the study of Edmunds et al. (2005) suggests that dressing percentage may have increased with an increase in the dose of lecithin (Mullan, unpublished). Larger on-farm studies are currently being conducted under the auspices of the Pork CRC to identify the effects of dietary lecithin on carcass dressing percentage.
Betaine
Betaine is a methyl donor and has a lipotropic effect (Barak et al., 1993). Betaine has been investigated as a partial replacement for choline in pig and poultry diets, as it is less expensive and less corrosive than choline. When incorporated into pig diets, betaine has been reported to improve growth performance by reducing the maintenance energy requirement of the animal, perhaps by reducing the need for ion pumping involved with maintaining intracellular osmolarity (Schrama et al., 2003). In addition, dietary betaine has been reported to increase protein deposition and carcass leanness (Fernandez-Figares et al., 2002; Matthews et al., 2001a; Suster et al., 2004) and to decrease back-fat depth (Cadogan et al., 1993). There is evidence that betaine has a more pronounced effect when dietary energy is limiting (Suster et al., 2004), thus offering a means of improving growth performance and meat quality by increasing the availability of energy. The additional energy may improve or maintain performance and meat quality under conditions in which performance may be limited by energy intake, e.g., in genetically improved pigs housed under commercial conditions and in pigs treated with metabolic modifiers such as pST (Suster et al., 2004) or ractopamine (Dunshea et al., 2009). Although betaine is used in many animal production systems, there are relatively few studies on the effects of betaine on meat quality. In one study, betaine supplementation resulted in improved carcass quality and improved subjective colour and marbling scores (Xu et al., 1999). Pork from pigs fed betaine was darker and had elevated ultimate pH and thaw loss and decreased cooking loss in another study (Matthews et al., 2001b). In a companion study, fresh pork from pigs fed betaine had increased initial pH and decreased drip loss (Matthews et al., 2001a). Cooking loss and total loss from frozen pork were decreased by dietary betaine in pigs with adequate pen space but were increased by dietary betaine in pigs with inadequate pen space. Shear force was not significantly altered in either of these studies (Matthews et al., 2001a,b). In another study, there was no effect of betaine supplementation on objective meat quality (Overland et al., 1999). Although it is difficult to make any conclusions on the effects of betaine on meat quality, there do appear to be some benefits in terms of water-holding capacity, which may be associated with the osmotic properties of betaine.
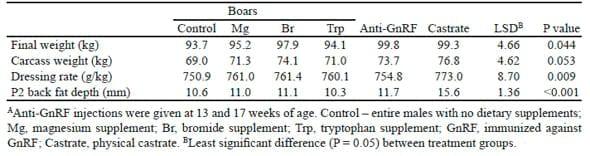
Table 3. Effect of sex and dietary additive between 17 and 22 weeks of age on final weight and carcass characteristics at slaughter (after McCauley et al., 2003 and unpublished)A
Dietary neuroleptics
In Australia, the pork industry has traditionally not castrated male pigs to take advantage of the improved carcass leanness and feed efficiency associated with entire males. Apart from the risk of boar taint, another disadvantage of entire male pig production is the likelihood of aggressive behaviour and sexual activity among group-housed boars (Cronin et al., 2003; Rhydmer et al., 2006; 2010), which may prevent them from performing to their potential. Cronin et al. (2003) found that immunization against gonadotrophin releasing factor (GnRF) to reduce boar taint also decreased sexual and aggressive behaviour to levels observed in physical castrates. In addition, dietary tryptophan reduces aggression in boars subjected to mixing during lairage (Pethick et al., 1997) and dietary magnesium reduces plasma catecholamine concentrations and the incidence of pale soft exudative pork in stressed pigs (D'Souza et al., 1998). Potassium bromide is a dietary neuroleptic that has been shown to decrease sexual and aggressive activities without altering growth rate in growing bulls (Genicot et al., 1991). In the early 2000s, our group conducted a series of studies to test the efficacy of dietary neuroleptics (McCauley et al., 2003; 2004), and there is now renewed interest in this approach within the Pork CRC.
The first study involved 200 entire boars and 40 contemporary castrates (physical castration at 2 weeks of age) (McCauley et al., 2003). Entire boars were allocated to the treatments at 13 weeks of age and housed in 10 pens of 20 pigs each (two pens per subsequent treatment). The physical castrates were housed in two pens of 20 pigs each. The immunized boars were given the first dose of an anti-GnRF vaccine (Improvac™, Pfizer Animal Health, Parkville) at this time. All pigs received a standard grower ration containing 14.0 MJ DE and 9.8 g true ileal digestible lysine per kg ad libitum until 17 weeks of age. Feed consumed and ADG were determined from 13 to 17 weeks of age. At 17 weeks of age, each group of 20 pigs was further divided into groups of 10 pigs (predetermined at 13 weeks of age) and moved into pens in the finisher shed. Immunized pigs (four pens of 10 pigs each) were given their second dose of the anti-GnRF vaccine at 17 weeks of age and dietary treatments began. Control boars, immunized boars and physical castrate boars were offered a commercial finisher ration containing 13.5 MJ DE and 7.4 g true ileal digestible lysine per kg. The three other dietary treatments offered to entire boars were a finisher diet supplemented with magnesium (5 g Mg proteinate per kg), bromide (140 mg Br chloride per kg) and tryptophan (Trp; 5 g/kg). All diets were offered ad libitum and feed intake and live weight were determined weekly on a per-pen basis.
There was no significant effect of sex or dietary treatment on ADG between 17 and 19 weeks or between 19 and 22 weeks. Over the entire period (17 to 22 weeks), the immunized boars had a greater ADG than all other classes of pigs. The ADG of physical castrates tended to be higher than that of the entire boars (P = 0.087). Feed intake of the control and Trp boars was less (P < 0.05) than that of the other groups over the first 2 weeks after transfer into the finisher shed. For the Trp pigs, this effect was most pronounced during the first week, whereas for the control boars, it was most pronounced during the second week (data not shown). Feed intake of the physical castrates was higher than that of the other groups over the first 2 weeks of the finishing period. There was no effect of any of the dietary additives on feed intake of entire boars over the latter part of the finishing phase (19–22 weeks). Boars immunized against GnRF increased their feed intake after the first week after secondary immunization to a level similar to that of the surgical castrates. There was no effect of sex or dietary additive on FCR over the first 2 weeks of the finishing period. However, over the latter part of the finishing phase, the FCR of the physical castrate pigs was 21% higher than that of the control boars. Over the entire finishing period, the FCR of the Trp boars was 10% lower than that of the control boars, whereas the FCR of the surgical castrates was 17% higher than that of the control boars. The FCR of boars immunized against GnRF and those offered diets containing Br or Mg were not different from that of the control boars.
The final live weights of castrates and boars immunized against GnRF were greater (P < 0.05) than those of control boars and boars fed Trp (Table 3). Relative to the control boars, carcass weight was significantly increased in the castrates, immunized boars and boars fed Br (Table 3). In the surgical castrates and the boars fed Br, the increase in carcass weight resulted from an increase in live weight as well as an increase in dressing rate, whereas for the immunized boars, the increase in carcass weight was the result of an increase in live weight but not in dressing rate. Dressing out rate was also increased by dietary Mg and Trp. Surgical castrates had greater P2 fat depth (+5 mm) than the control boars, but none of the dietary additives affected P2 fat depth, nor did immunization.
A subsequent study (McCauley et al., 2004), investigated the effects of Br and Trp, alone or in combination, in pigs that were either light, average or heavy for age. Immunization against GnRF was used as a positive control. Over the period from 17 to 22 weeks, the immunized males grew more quickly than all other classes of pig. In particular, the immunized males grew 19% (+153 g/d) faster than the control boars. However, there was an interaction between treatment and weight such that growth response was greatest in the medium weight class of pigs treated with Improvac and least in the light pigs. Whereas there were no significant individual effects of either Br (+1.8%) or Trp (+2.2%) on daily gain, pigs treated with both compounds grew significantly faster than the controls (10.3%, P < 0.05). Pigs from all treatment groups grew faster than the control boars, but there was an interaction between treatment and weight class such that this effect was most pronounced in the heavy pigs.
Conclusions
Dietary additives offer a means of manipulating growth performance and carcass quality. However, responses are often variable and inconsistent and further research is needed to identify the circumstances under which they are most effective. Although responses to dietary feed additives are generally not as great as responses to porcine somatotropin, they do offer an alternative means of manipulating growth performance and carcass quality.
This paper was presented at the Recent Advances in Animal Nutrition- Australia, July 2011.
Related topics
Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post




You may be interested in
.jpg&w=3840&q=75)
Evonik Animal Nutrition