Introduction
The increase in litter size over recent decades has reduced the uterine space available for fetal growth and development, thus reducing individual piglet birth weight. Consequently, lower birth weight reduces piglet survivability, wean weight, and market weight. However, few nutritional options have been identified to mitigate reduced birth weight. There is evidence that increasing feed intake in late gestation improves piglet birth weight (Cromwell et al., 19894 , Shelton et al., 20095 , Soto et al., 20116 ). However, the effect of increased amounts of feed offered during late gestation on piglet birth weight in high-performing herds (> 14.5 total piglets born/sow) is unclear. Further, because all studies reported increased feed intake, which increased both energy and amino acids (AA), it is unclear if the influences on piglet birth weight are due to dietary AA or energy content. The objective of this study was to determine the effects of AA and energy intake during late gestation on piglet birth weight and reproductive performance of high-performing gilts and sows housed under commercial conditions. The hypothesis was that both maternal dietary AA and energy in the late gestation period would affect piglet birth weight.
Procedures
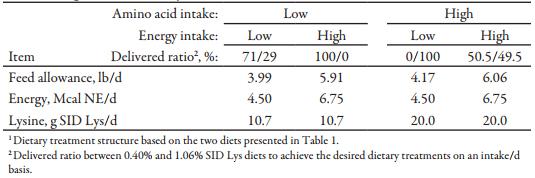
The Kansas State University Institutional Animal Care and Use Committee approved the protocol used in this experiment. The experiment was conducted at a commercial sow farm in northern Ohio during the summer. Females were individually housed from d 0 to 40 of gestation, and then were group-housed (20 and 18 sq. ft/animal for gilts and sows, respectively) from d 40 to 111 of gestation. Each group pen was equipped with an electronic sow feeding station (Schauer, Prambachkirchen, Austria) and two cup waterers. All females had ad libitum access to water. From d 0 to 89 of gestation, females were fed a common diet with 0.59% standardized ileal digestible (SID) Lys according to body condition: Thin, ideal, and fat females were fed 7.0, 4.4, and 4.0 lb/d, respectively. To be eligible for enrollment in the experiment, at d 90 of gestation females must not have: 1) returned to estrus during the previous cycle, 2) had an abortion during the previous cycle, 3) moderate or greater severity lameness, or 4) body condition score less than 2. At d 90 of gestation, a total of 1,102 females (PIC 1050, Hendersonville, TN; 741 gilts and 361 sows) were housed in pens by parity group (P1 or P2+), blocked by weight within each pen, and each female was randomly assigned to dietary treatments within weight block. The average parity for P2+ (sows) after farrowing was 4.1 ± 1.9 cycles (mean ± SD). Dietary treatments consisted of combinations of 2 SID AA intakes (10.7 or 20.0g SID Lys/d and other AA met or exceeded the NRC [2012] recommendations as a ratio to Lys) and 2 energy intakes (4.50 or 6.75 Mcal/d intake of NE) in a 2 x 2 factorial arrangement. All other nutrients met or exceeded the NRC (2012) recommendations.
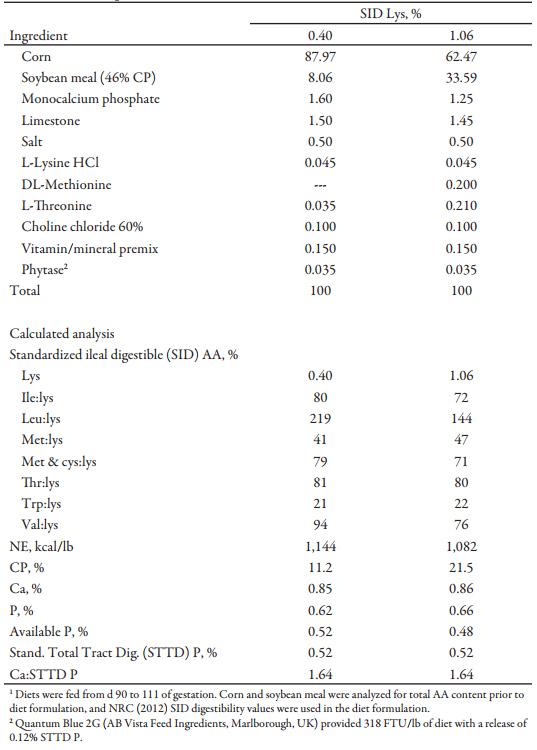
Two diets were formulated (Table 1) and delivered at 4 different ratios and intake levels to achieve the desired dietary treatments for the 2 × 2 factorial (Table 2). Diets were balanced on a Ca to standardized total tract digestible (STTD) P ratio basis. Phytase was included in both diets with release considered to be 0.12% for STTD P. No AA release was considered for phytase.
At d 111 of gestation, females were moved to the farrowing house and fed 8.0 lb/d of a common lactation diet with 1.25% SID Lys provided until farrowing and then provided the same diet ad libitum thereafter. Both gestation and lactation diets were corn-soybean meal-based and presented in meal form. After weighing individual piglets, litter size was equalized regardless of dietary treatment.
On the subsequent cycle, no dietary treatments were applied, and females were fed a common diet with 0.59% SID Lys according to body condition. Thin, ideal, and fat females were fed 7.0, 4.4, and 4.0 lb/d, respectively, until d 90 of gestation, and then feed allowance was increased by 2.0 lb/d for thin and ideal condition females.
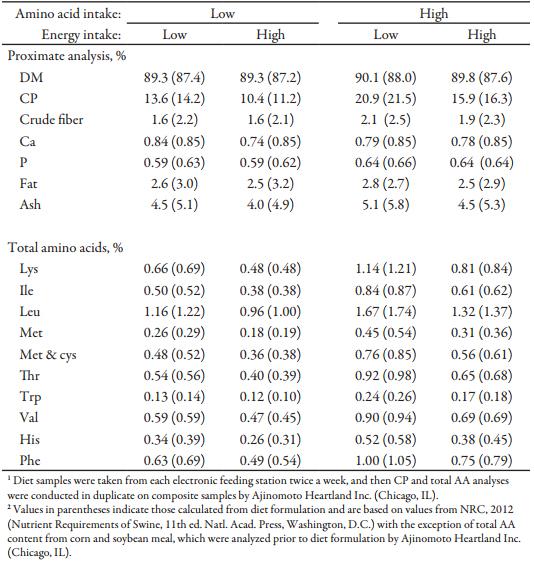
Prior to diet formulation, 5 samples of corn and soybean meal were submitted for proximate and total AA analysis. The analyzed values were used in formulation in conjunction with NRC (2012) digestibility coefficients. Samples of the diets were submitted for analysis of DM, CF, ash, crude fat, Ca, and P (Ward Laboratories, Inc., Kearney, NE). Diet samples were taken from each electronic feeding station twice a week, and then CP and total AA analyses were conducted in duplicate on composite samples (Ajinomoto Heartland Inc., Chicago, IL). Feeding station calibration was monitored twice a week by weighing 10 samples from each dispenser in each station.
Data were analyzed using generalized linear mixed models with parity group and dietary treatments as the linear predictor and random effects of pen as the experimental unit for parity and the individual female as the experimental unit for dietary treatments. Models of non-normal responses were initially fitted with Laplace method of approximation to evaluate for overdispersion (Stroup, 2012)7 . Then, models were fit using the restricted maximum likelihood estimation for inference. Response variables were fitted accordingly with distribution. Bonferroni adjustment was used for multiple comparisons. Statistical models were fitted using the GLIMMIX procedure of SAS (SAS Institute Inc., Cary, NC). Results were considered significant at P ≤ 0.05 and marginally significant at 0.05 < P ≤ 0.10.
Results and Discussion
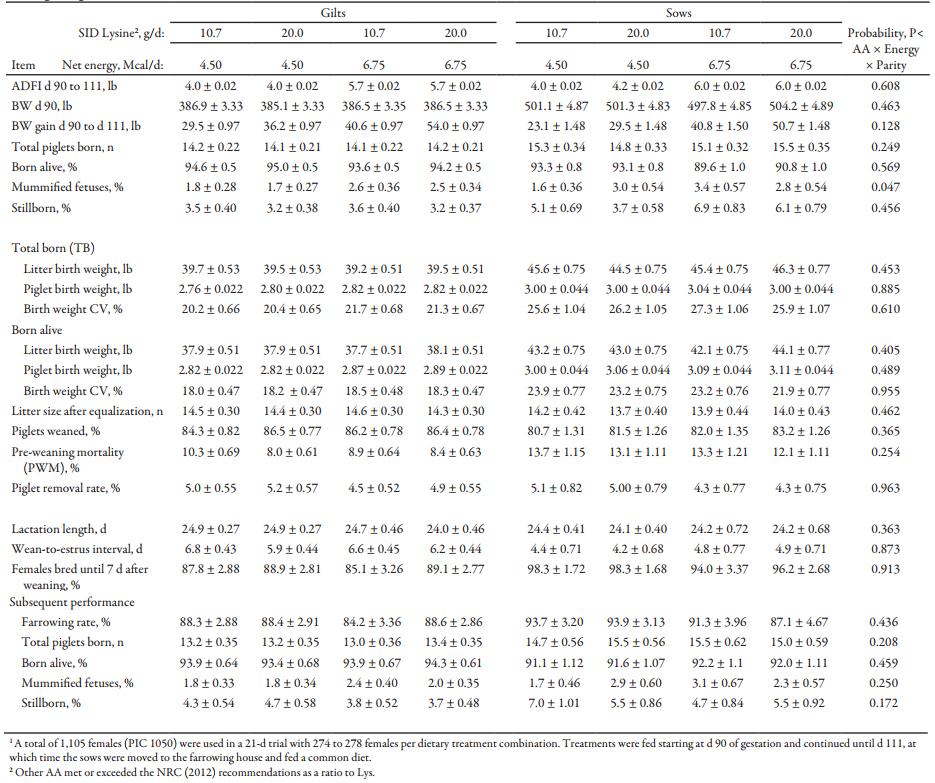
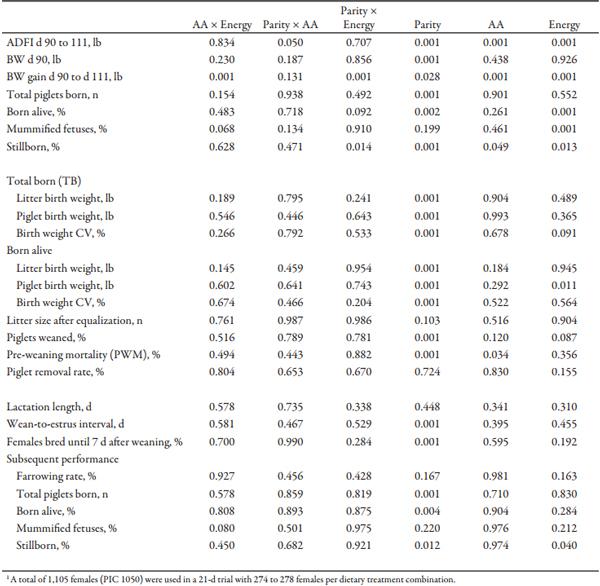
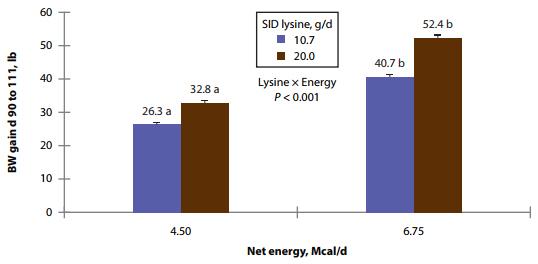
Chemical analysis of feed was similar to formulated values (Table 3). In sows and gilts fed high-energy intake, the magnitude of BW gain was greater with increasing AA intake (Fig. 1A) compared to increasing Lys intake at low-energy intake (AA × energy, P < 0.001; Tables 4 and 5). Gilts gained more weight at low energy intake than sows (Fig. 1B; parity × energy, P < 0.001); however, there was no evidence for differences (P = 0.601) in weight gain between gilts and sows at high-energy intake.
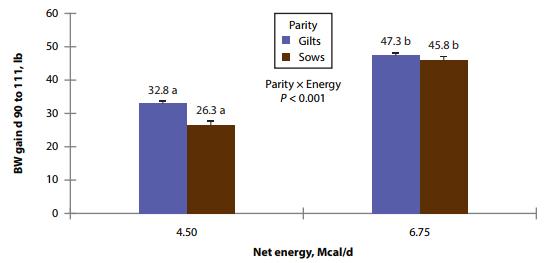
There was no evidence of differences in the number of piglets total born between the dietary treatments. However, sows had an increased number (P < 0.001) of total piglets born, as compared with gilts. Sows fed the high-energy intake had marginally reduced probability of piglets born alive (parity × energy, P = 0.001), but there was no evidence for differences in gilts. This was due to the increased probability (parity × energy, P = 0.014) of stillborns to be greater in sows fed high-energy intake (Fig. 2). The probability of stillborns was reduced (P = 0.049) in females fed high AA intake. There was a AA × energy × parity (P = 0.047) interaction: Sows fed low-energy and high-AA intake had increased probability of mummified fetuses, as compared to sows fed low-energy but low-AA intake.
Considering the total number of piglets born, litter birth weight and individual piglet birth weight were heavier in sows (P < 0.001) than gilts, with marginally greater (P = 0.091) within-litter birth weight CV in females fed high energy compared to low energy, independent of parity. Within-litter birth weight, CV was greater (P < 0.001) in sows than gilts. However, there was no evidence for differences between the dietary treatments for litter birth weight and individual piglet birth weight of total piglets born.
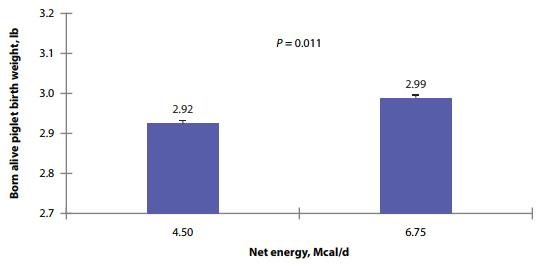
When litter birth weight and individual birth weight for piglets born alive were considered, weights were heavier in sow litters (P < 0.001) compared to gilt litters. Individual born-alive birth weight (Fig. 3) was heavier (P = 0.011) for females fed the high-energy intake compared to low-energy intake fed females, regardless of AA intake or parity. Born-alive piglets from sows were heavier (P < 0.001) than those from gilts. There was no evidence for differences in the within litter birth weight CV between the dietary treatments. Within-litter birth weight CV was greater (P < 0.001) in sows than gilts.
Piglets suckling from females fed high AA intake showed a lower percentage (P = 0.034) of pre-weaning mortality (PWM) compared to those born to females fed low AA intake during late gestation, regardless of energy level.
There was no evidence of differences in lactation length between dietary treatment or parity. The percentage of females bred 7 d after weaning was greater (P < 0.001) for sows than for gilts due to shorter (P < 0.001) wean-to-estrus interval (WEI) in sows compared to gilts. However, there was no evidence for differences in percentage of females bred by 7 d after weaning or WEI between the dietary treatments.
There was no evidence for differences on subsequent farrowing rate between the dietary treatments or parity level. Sows had greater (P < 0.001) number of total piglets born compared to gilts on the subsequent cycle. However, gilts had increased (P < 0.004) probability of more piglets born alive compared to sows on the subsequent cycle. There was no evidence for differences between the dietary treatments on the number of total piglets born, and probability of born-alive piglets on the subsequent cycle. However, when stillborns were analyzed separately from mummified fetuses, females fed high energy had lower (P = 0.040) probability of stillborn piglets in the subsequent cycle compared to those fed low energy.
In conclusion, 1) body weight gain of gilts and sows depends not only on energy but also AA intake, 2) sows fed increased amount of energy had an increased stillborn rate, 3) pre-weaning mortality was reduced in piglets suckling from females with high AA intake, and 4) the positive effect of increased amount of feed during late gestation on individual piglet birth weight, 30 g per pig, was due to energy rather than AA intake.
This article was originally published in Kansas Agricultural Experiment Station Research Reports: Vol. 1: Iss. 7. https://doi.org/10.4148/2378-5977.1112.
Table 1. Diet composition (as-fed basis)1
Table 2. Experimental dietary treatment structure1
Table 3. Chemical analysis of the diets (as-fed-basis)1
Table 4. Least square mean estimates (and corresponding SEM) for reproductive performance of gilts and sows fed different lysine and energy intake levels during late gestation under commercial conditions1
Table 5. Probability values of the effects of different lysine and energy intake levels during late gestation on reproductive performance of gilts and sows under commercial conditions1
Figure 1A. Effects of different lysine and energy intake levels fed from d 90 to d 111 of gestation on BW gain of gilts and sows. Means with a different superscript within SID Lys level differ (P < 0.05).
Figure 1B. Effects of different energy intake levels fed from d 90 to d 111 of gestation on BW gain of gilts and sows. Means with a different superscript within parity level differ (P < 0.05).
Figure 2. Effects of different energy intake levels fed from d 90 to d 111 of gestation on stillborn rate of gilts and sows. Means with a different superscript within parity level differ (P < 0.05).
Figure 3. Effects of different energy intake fed from d 90 to d 111 of gestation on individual born live piglet birth weight of gilts and sows.











.jpg&w=3840&q=75)