Dietary supplementation with 0.4% L-arginine between days 14 and 30 of gestation enhances NO and polyamine syntheses and water transport in porcine placentae
Background: Most embryonic loss in pigs occurs before d 30 of gestation. Dietary supplementation with L-arginine (Arg) during early gestation can enhance the survival and development of conceptuses (embryo/fetus and its extraembryonic membranes) in gilts. However, the underlying mechanisms remain largely unknown.
Methods: Between d 14 and 30 of gestation, each gilt was fed daily 2 kg of a corn- and soybean-meal based diet (12% crude protein) supplemented with either 0.4% Arg (as Arg-HCl) or an isonitrogenous amount of L-alanine (Control). There were 10 gilts per treatment group. On d 30 of gestation, gilts were fed either Arg-HCl or L-alanine 30 min before they were hysterectomized, followed by the collection of placentae, embryos, fetal membranes, and fetal fluids. Amniotic and allantoic fluids were analyzed for nitrite and nitrate [NOx; stable oxidation products of nitric oxide (NO)], polyamines, and amino acids. Placentae were analyzed for syntheses of NO and polyamines, water and amino acid transport, concentrations of amino acid-related metabolites, and the expression of angiogenic factors and aquaporins (AQPs).
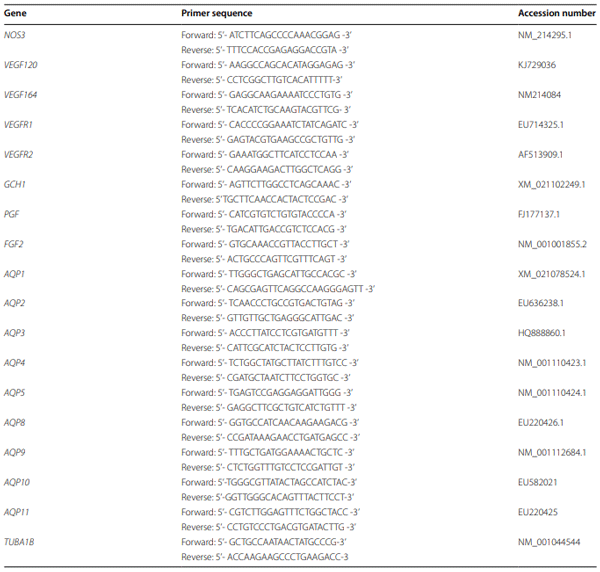
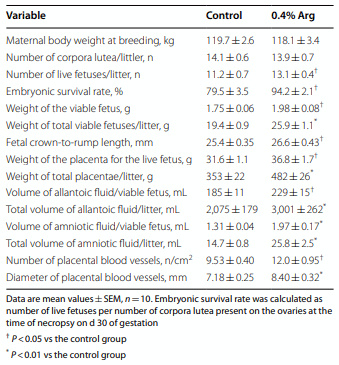
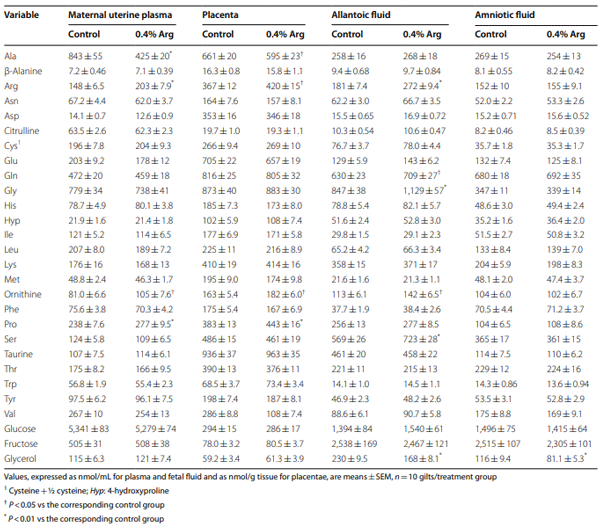
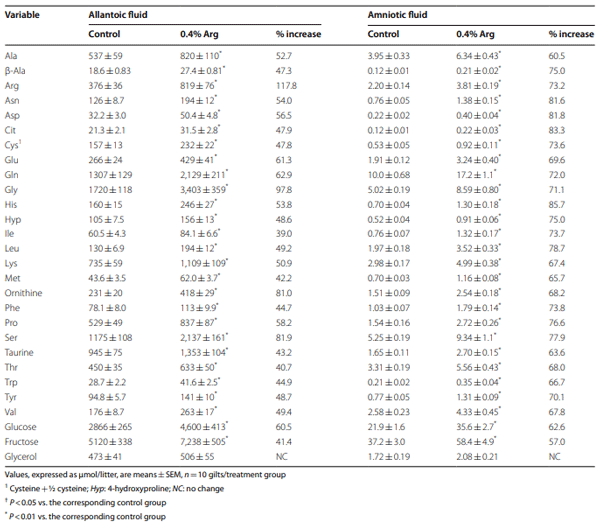
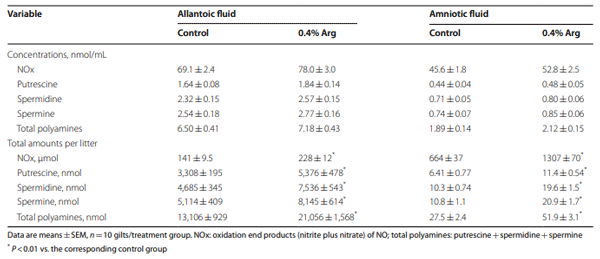
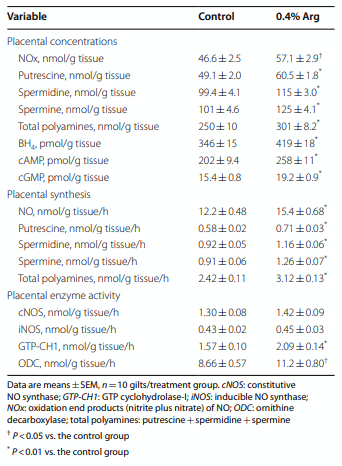
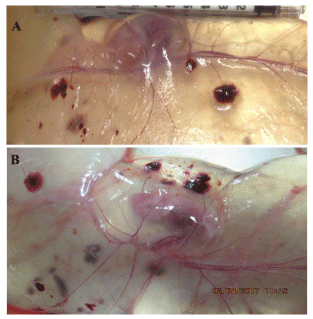
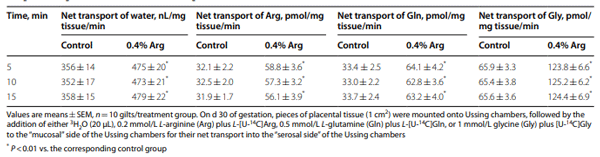
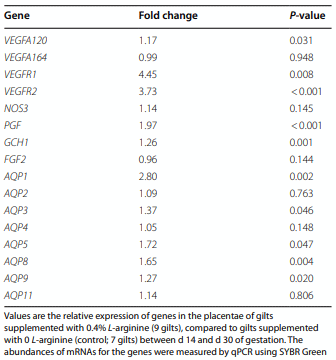
Results: Compared to the control group, Arg supplementation increased (P<0.05) the number of viable fetuses by 1.9 per litter, the number and diameter of placental blood vessels (+25.9% and+17.0% respectively), embryonic survival (+18.5%), total placental weight (+36.5%), the total weight of viable fetuses (+33.5%), fetal crown-to-rump length (+4.7%), and total allantoic and amniotic fluid volumes (+44.6% and+75.5% respectively). Compared to control gilts, Arg supplementation increased (P<0.05) placental activities of GTP cyclohydrolase-1 (+33.1%) and ornithine decarboxylase (+29.3%); placental syntheses of NO (+26.2%) and polyamines (+28.9%); placental concentrations of NOx (+22.5%), tetrahydrobiopterin (+21.1%), polyamines (+20.4%), cAMP (+27.7%), and cGMP (+24.7%); total amounts of NOx (+61.7% to+96.8%), polyamines (+60.7% to+88.7%), amino acids (+39% to + 118%), glucose (+60.5% to+62.6%), and fructose (+41.4% to+57.0%) in fetal fluids; and the placental transport of water (+33.9%), Arg (+78.4%), glutamine (+89.9%), and glycine (+89.6%). Furthermore, Arg supplementation increased (P<0.05) placental mRNA levels for angiogenic factors [VEGFA120 (+117%), VEGFR1 (+445%), VEGFR2 (+373%), PGF (+197%), and GCH1 (+126%)] and AQPs [AQP1 (+280%), AQP3 (+137%), AQP5 (+172%), AQP8 (+165%), and AQP9 (+127%)].
Conclusion: Supplementing 0.4% Arg to a conventional diet for gilts between d 14 and d 30 of gestation enhanced placental NO and polyamine syntheses, angiogenesis, and water and amino acid transport to improve conceptus development and survival.
Keywords: Angiogenesis, Arginine, Fetus, Placenta, Reproduction.
![Fig. 2 Proposed mechanisms responsible for benefcial efects of dietary L-arginine supplementation in improving embryonic/fetal growth and survival in gestating swine. L-Arginine stimulates the synthesis of tetrahydrobiopterin [BH4, a required co-factor for nitric oxide (NO) synthase)] from GTP via the GTP cyclohydrolase-I (GCH1) pathway, thereby augmenting NO production by placental tissue. L-Arginine also increases the activity of ornithine decarboxylase (a key enzyme for the synthesis of polyamines). Both NO and polyamines, as well as growth factors (such as placental growth factor, vascular endothelial growth factor A120, and vascular endothelial growth factor receptors 1 and 2) promote placental angiogenesis and growth (including vascular growth) to increase rates of transfer of non-water nutrients [including amino acids (AAs)] and oxygen across the placenta from mother to embryo/fetus. In addition, L-arginine elevates the concentrations of both cGMP and cAMP in the placenta to increase the expression of aquaporins (AQPs) to promote the placental transport of water from mother to embryo/fetus. Ultimately, the coordinate actions of L-arginine result in improvements in the growth and survival of embryos/fetuses](/_next/image/?url=https%3A%2F%2Fimages.engormix.com%2FE_articles%2F51735_162.gif&w=1080&q=75)
1. Bazer FW, Song GW, Kim JY, Dunlap KA, Satterfeld MC, Johnson GA, et al. Uterine biology in sheep and pigs. J Anim Sci Biotechnol. 2012;3:23.
2. Edwards AK, Wessels JM, Kerr A, Tayade C. An overview of molecular and cellular mechanisms associated with porcine pregnancy success or failure. Reprod Dom Anim. 2012;47:394–401.
3. Bazer FW, Johnson GA. Pig blastocyst-uterine interactions. Difer. 2014;87:52–65.
4. Johnson GA, Bazer FW, Seo H. The early stages of implantation and placentation in the pig. In: Geisert RD, Spencer T, editors. Placentation in mammals. Advances in anatomy, embryology and cell biology, vol. 234. Cham: Springer; 2021. p. 61–89. https://doi.org/10.1007/ 978-3-030-77360-1_5.
5. Moreira RHR, Pérez Palencia JY, Moita VHC, Caputo LSS, Saraiva A, Ferreira RA, et al. Variability of piglet birth weights: A systematic review and meta-analysis. J Anim Physiol Anim Nutr (Berl). 2020;104:657–66.
6. Palencia JYP, Lemes MAG, Garbossa CAP, Abreu MLT, Pereira LJ, Zangeronimo MG. Arginine for gestating sows and foetal development: a systematic review. J Anim Physiol Anim Nutr (Berl). 2008;102:204–13.
7. Wu G, Bazer FW, Satterfeld MC, Li X, Wang X, Johnson GA, et al. Impacts of arginine nutrition on embryonic and fetal development in mammals. Amino Acids. 2013;45:241–56.
8. Herring CM, Bazer FW, Johnson GA, Wu G. Impacts of maternal dietary protein intake on fetal survival, growth and development. Exp Biol Med. 2018;243:525–33.
9. Ji Y, Wu Z, Dai Z, Wang X, Li J, Wang B, et al. Fetal and neonatal programming of postnatal growth and feed efciency in swine. J Anim Sci Biotech. 2017;8:42.
10. Hou YQ, Yin YL, Wu G. Dietary essentiality of “nutritionally nonessential amino acids” for animals and humans. Exp Biol Med. 2015;240:997–1007.
11. Wu G, Bazer FW, Burghardt RC, Johnson GA, Kim SW, Li XL, et al. Impacts of amino acid nutrition on pregnancy outcome in pigs: mechanisms and implications for swine production. J Anim Sci. 2010;88:195–204.
12. Wu G, Bazer FW, Johnson GA, Hou Y. Arginine nutrition and metabolism in growing, gestating, and lactating swine. J Anim Sci. 2018;96:5035–51.
13. Meininger CJ, Wu G. Regulation of endothelial cell proliferation by nitric oxide. Methods Enzymol. 2022;352:280–95.
14. Chen DB, Zheng J. Regulation of placental angiogenesis. Microcirc. 2014;21:15–25.
15. Elmetwally MA, Li XL, Johnson GA, Burghardt RC, Herring CM, Kramer AC, et al. Dietary supplementation with L-arginine between Days 14 and 25 of gestation enhances NO and polyamine syntheses and the expression of angiogenic proteins in porcine placentae. Amino Acids. 2022;54:193–204.
16. Blachier F, Davila AM, Benamouzig R, Tome D. Channelling of arginine in NO and polyamine pathways in colonocytes and consequences. Front Biosci (Landmark Ed). 2011;16:1331–43.
17. Jobgen WS, Fried SK, Fu WJ, Meininger CJ, Wu G. Regulatory role for the arginine-nitric oxide pathway metabolism of energy substrates. J Nutr Biochem. 2006;17:571–88.
18. Agostinelli E. Biochemical and pathophysiological properties of polyamines. Amino Acids. 2020;52:111–7.
19. Durante W. Amino acids in circulatory function and health. Adv Exp Med Biol. 2020;1265:39–56.
20. Hou YQ, Hu SD, Jia SC, Nawaratna G, Che DS, Wang FL, et al. Wholebody synthesis of L-homoarginine in pigs and rats supplemented with L-arginine. Amino Acids. 2016;48:993–1001.
21. Li H, Meininger CJ, Hawker JR Jr, Haynes TE, Kepka-Lenhart D, Mistry SK, et al. Regulatory role of arginase I and II in nitric oxide, polyamine, and proline syntheses in endothelial cells. Am J Physiol. 2001;280:E75-82.
22. Wu G, Bazer FW, Johnson GA, Herring C, Seo H, Dai Z, et al. Functional amino acids in the development of the pig placenta. Mol Reprod Dev. 2017;84:870–82.
23. Li XL, Johnson GA, Zhou HJ, Burghardt RC, Bazer FW, Wu G. Microarray analysis reveals an important role for dietary L-arginine in regulating global gene expression in porcine placentae during early gestation. Front Biosci (Landmark ed). 2022;27(1):033.
24. Li X, Bazer FW, Johnson GA, Burghardt RC, Frank JW, Dai Z, et al. Dietary supplementation with L-arginine between days 14 and 25 of gestation enhances embryonic development and survival in gilts. Amino Acids. 2014;46:375–84.
25. Zhang Q, Hou YQ, Bazer FW, He WL, Posey EA, Wu G. Amino acids in swine nutrition and production. Adv Exp Med Biol. 2020;1285:81–107.
26. Mateo RD, Wu G, Bazer FW, Park JC, Shinzato I, Kim SW. Dietary L-arginine supplementation enhances the reproductive performance of gilts. J Nutr. 2007;137:652–6.
27. Zeng XF, Wang FL, Fan X, Yang WJ, Zhou B, Li PF, et al. Dietary arginine supplementation during early pregnancy enhances embryonic survival in rats. J Nutr. 2008;138:1421–5.
28. Zeng XF, Mao X, Huang Z, Wang FL, Wu G, Qiao S. Arginine enhances embryo implantation in rats through PI3K/PKB/mTOR/NO signaling pathway during early pregnancy. Reprod. 2013;145:1–7.
29. Li X, Bazer FW, Johnson GA, Burghardt RC, Erikson DW, Frank JW, et al. Dietary supplementation with 0.8% L-arginine between Days 0 and 25 of gestation reduces litter size in gilts. J Nutr. 2010;140:1111–6.
30. Bazer FW, Goldstein MH, Barron DH. Water and electrolyte transport by pig chorioallantois. In: Mastroainni L, Biggers JD, editors. Fertilization and embryonic development in vitro. New York: Plenum; 1981. p. 200–21.
31. Bazer FW, Thatcher WW, Martinat-Botte F, Terqui M. Conceptus development in large white and prolifc Chinese Meishan pigs. J Reprod Infertil. 1988;84:37–42.
32. Markou A, Unger L, Abir-Awan M, Saadallah A, Halsey A, Balklava Z, et al. Molecular mechanisms governing aquaporin relocalisation. Biochim Biophys Acta. 2022;1864:183853.
33. Zhu C, Jiang Z, Bazer FW, Johnson GA, Burghardt RC, Wu G. Aquaporins in the female reproductive system of mammals. Front Biosci (Landmark ed). 2015;20:838–71.
34. Laloux T, Junqueira B, Maistriaux LC, Ahmed J, Jurkiewicz A, Chaumont F. Plant and mammal aquaporins: same but diferent. Int J Mol Sci. 2018;19:521.
35. Nesverova V. Törnroth-Horsefeld S (2019) Phosphorylation-dependent regulation of mammalian aquaporins. Cells. 2019;8(2):82.
36. Zhu C, Ye JL, Bai YS, Hu SD, Tan CQ, Bazer FW, et al. Arginine promotes the expression of aquaporin-3 and water transport in porcine trophectoderm cells through NO- and cAMP-dependent mechanisms. Front Biosci (Landmark ed). 2022;27(3):083.
37. Zhu C, Jiang ZY, Johnson GA, Bazer FW, Wu G. Nutritional and physiological regulation of water transport in the conceptus. Adv Exp Med Biol. 2022;1354:109–25.
38. McLendon BA, Kramer AC, Seo H, Burghardt RC, Bazer FW, Wu G, et al. Temporal and spatial expression of aquaporins 1, 5, 8, and 9: Potential transport of water across the endometrium and chorioallantois of pigs. Placenta. 2022;124:28–36.
39. Wu G. Nutrition and metabolism: foundations for animal growth, development, reproduction, and health. Adv Exp Med Biol. 2022;1354:1–24.
40. Dai ZL, Wu ZL, Jia SC, Wu G. Analysis of amino acid composition in proteins of animal tissues and foods as pre-column o-phthaldialdehyde derivatives by HPLC with fuorescence detection. J Chromatogr B. 2014;964:116–27.
41. Wu G, Bazer FW, Tuo W, Flynn SP. Unusual abundance of arginine and ornithine in porcine allantoic fuid. Biol Reprod. 1996;54:1261–5.
42. Kong XF, Wang XQ, Yin YL, Li XL, Gao HJ, Bazer FW, et al. Putrescine stimulates the mTOR signaling pathway and protein synthesis in porcine trophectoderm cells. Biol Reprod. 2014;91:106.
43. Li H, Meininger CJ, Wu G. Rapid determination of nitrite by reversedphase high-performance liquid chromatography with fuorescence detection. J Chromatogr B. 2000;746:199–207.
44. Jobgen WS, Jobgen SC, Li H, Meininger CJ, Wu G. Analysis of nitrite and nitrate in biological samples using high-performance liquid chromatography. J Chromatogr B. 2007;851:71–82.
45. Wu G, Bazer FW, Hu J, Johnson GA, Spencer TE. Polyamine synthesis from proline in the developing porcine placenta. Biol Reprod. 2005;72:842–50.
46. Dillon EL, Wu G. Cortisol enhances ctrulline synthesis from proline in enterocytes of suckling piglets. Amino Acids. 2021;53:1957–66.
47. Furukawa K, He WL, Bailey CA, Bazer FW, Toyomizu M, Wu G. Polyamine synthesis from arginine and proline in tissues of developing chickens. Amino Acids. 2021;53:1739–48.
48. Haynes TE, Li P, Li XL, Shimotori K, Sato H, Flynn NE, et al. L-Glutamine or L-alanyl-L-glutamine prevents oxidant- or endotoxin-induced death of neonatal enterocytes. Amino Acids. 2009;37:131–42.
49. He WL, Furukawa K, Bailey CA, Wu G. Oxidation of amino acids, glucose, and fatty acids as metabolic fuels in enterocytes of post-hatching developing chickens. J Anim Sci. 2022;100:skac053.
50. Jobgen WJ, Meininger CJ, Jobgen SC, Li P, Lee MJ, Smith SB, et al. Dietary L-arginine supplementation reduces white-fat gain and enhances skeletal muscle and brown fat masses in diet-induced obese rats. J Nutr. 2009;139:230–7.
51. Meininger CJ, Marinos RS, Hatakeyama K, Martinez-Zaguilan R, Rojas JD, Kelly K, et al. Impaired nitric oxide production in coronary endothelial cells of the spontaneously diabetic BB rat is due to tetrahydrobiopterin defciency. Biochem J. 2000;349:353–6.
52. Meininger CJ, Cai S, Parker JL, Channon KM, Kelly KA, Becker EJ, et al. GTP cyclohydrolase I gene transfer reverses tetrahydro-biopterin defciency and increases nitric oxide synthesis in endothelial cells and isolated vessels from diabetic rats. FASEB J. 2004;18:1900–2.
53. Zhu C, Li XL, Bazer FW, Johnson GA, Burghardt RC, Jiang ZY, et al. Dietary L-arginine supplementation during days 14–25 of gestation enhances aquaporin expression in the placentae and endometria of gestating gilts. Amino Acids. 2021;53:1287–95.
54. Wu G, Li X, Seo H, McLendon BA, Kramer AC, Bazer FW, et al. Osteopontin (OPN)/secreted phosphoprotein 1 (SPP1) binds integrins to activate transport of ions across the porcine placenta. Front Biosci (Landmark ed). 2022;27:117–27.
55. Steinhauser CB, Wing TT, Gao HJ, Li XL, Burghardt RC, Wu G, et al. Identifcation of appropriate reference genes for qPCR analyses of placental expression of SLC7A3 and induction of SLC5A1 in porcine endometrium. Placenta. 2017;52:1–9.
56. Elmetwally MA, Halawa AA, Tang W, Wu G, Bazer FW. Efects of bisphenol A on expression of genes related to amino acid transporters, insulin- like growth factor, aquaporin and amino acid release by porcine trophectoderm cells. Reprod Toxicol. 2020;96:241–8.
57. Fu WJ, Hu J, Spencer T, Carroll R, Wu G. Statistical models in assessing fold changes of gene expression in real-time RT-PCR experiments. Comput Biol Chem. 2006;30:21–6.
58. Steel RGD, Torrie JH. Principles and procedures of statistics. New York: McGraw-Hill; 1980.
59. Bazer FW, Burghardt RC, Johnson GA, Spencer TE, Wu G. Mechanisms for the establishment and maintenance of pregnancy: synergies from scientifc collaborations. Biol Reprod. 2018;99:225–41.
60. Bérard J, Bee G. Efects of dietary L-arginine supplementation to gilts during early gestation on foetal survival, growth and myofber formation. Animal. 2010;4:1680–7.
61. Li J, Xia HA, Yao W, Wang TT, Li JL, Piao XS, et al. Efects of arginine supplementation during early gestation (d 1 to 30) on litter size and plasma metabolites in gilts and sows. J Anim Sci. 2015;93:5291–303.
62. Wu ZL, Hou YQ, Hu SD, Bazer FW, Meininger CJ, McNeal CJ, et al. Catabolism and safety of supplemental L-arginine in animals. Amino Acids. 2016;48:1541–52.
63. Wu G. Amino Acids: Biochemistry and Nutrition. 2nd ed. Boca Raton: CRC Press; 2022.
64. Reynolds LP, Caton JS, Redmer DA, Grazul-Bilska AT, Vonnahme KA, Borowicz PB, et al. Evidence for altered placental blood fow and vascularity in compromised pregnancies. J Physiol. 2006;572:51–8.
65. Vonnahme KA, Wilson ME, Ford SP. Relationship between placental vascular endothelial growth factor expression and placental/endometrial vascularity in the pig. Biol Reprod. 2001;64:1821–5.
66. Roskoski R. VEGF receptor protein-tyrosine kinases: structure and regulation. Biochem Biophys Res Comm. 2008;375:287–91.
67. Sanchis EG, Cristofolini AL, Merkis CI. Porcine placental immunoexpression of vascular endothelial growth factor, placenta growth factor, Flt-1 and Flk-1. Biotech Histochem. 2015;90:486–94.
68. Li H, Meininger CJ, Bazer FW, Wu G. Intracellular sources of ornithine for polyamine synthesis in endothelial cells. Amino Acids. 2016;48:2401–10.
69. McKnight JR, Satterfeld MC, Jobgen WS, Smith SB, Spencer TE, Meininger CJ, et al. Benefcial efects of L-arginine on reducing obesity: Potential mechanisms and important implications for human health. Amino Acids. 2010;39:349–57.
70. Anthony TL, Brooks HL, Boassa D, Leonov S, Yanochko GM, Regan JW, et al. Cloned human aquaporin-1 is a cyclic GMP-gated ion channel. Mol Pharmacol. 2000;57:576–88.
71. Baetz NW, Stamer WD, Yool AJ. Stimulation of aquaporin-mediated fuid transport by cyclic GMP in human retinal pigment epithelium in vitro. Invest Ophthalmol Vis Sci. 2012;53:2127–32.
72. Wu G, Bazer FW, Johnson GA. Regulation of water and ion transport by arginine in porcine conceptuses. USDA-NIFA Meeting Abstract. July 10, New Orleans, LA. 2018.
73. Wu G. Functional amino acids in growth, reproduction and health. Adv Nutr. 2010;1:31–7.
74. Wu G, Li P. The “ideal protein” concept is not ideal in animal nutrition. Exp Biol Med. 2022;247:1191–201.







.jpg&w=3840&q=75)