Reducing mycotoxin impact: the science behind Mycosorb
Mycotoxins are secondary metabolites secreted by various moulds such as Aspergillus, Penicillium and Fusarium. They are synthesised during the production, transport and storage of food supplies in certain environmental conditions. Detectable levels of nearly 300 potentially toxic metabolites secreted by moulds contaminate 25 to 40% of the cereals and cereal products worldwide (Pittet, 1998; Yiannikouris and Jouany, 2002).
Despite recent food crises (e.g., bovine spongiform encephalitis, listeriosis, salmonellosis, dioxins), consumers are still largely unaware of the risk associated with the natural presence of toxins or their metabolites in foods for human and animal consumption (Guerre et al., 2000).
However, these contaminants can cause numerous disorders and diseases, which sometimes prove fatal in animals (Pfohl-Leszkowicz, 2000) or humans (CAST, 2003; Krishnamachari et al., 1977), demonstrating the potential danger associated with mycotoxins. This risk has existed from the dawn of organised agricultural practices (Pittet, 1998).
For example the Old Testament speaks of ergotism (Schoental, 1984) and indirectly attributes the decline of the Etruscan civilisation to T-2 toxin and/or to zearalenone (Schoental, 1991). The first modern mycotoxicosis due to aflatoxins was reported in the 1960s, following the death of 100,000 turkeys in Great Britain from liver necrosis and biliary hyperplasia.
Mycotoxins may be hepatotoxic, haematotoxic, nephrotoxic, immunotoxic, neurotoxic, mutagenic, genotoxic, reprotoxic, teratogenic and carcinogenic.
Furthermore, multiple contaminations often result in synergy, increasing the danger associated with individual mycotoxins.
Integrated strategies to monitor mycotoxin risks
The effective control of food contamination levels requires various complementary strategies. An integrated strategy can be applied to fight mould growth and mycotoxin production by using resistant crops and rotating crops at field level; by using good agricultural practices and accurate pesticide/insecticide application during plant growth; by avoiding physical damage and over-contamination and by eliminating damaged grain during harvest; by controlling the level of contamination and environmental conditions, using effective preservatives and by applying physical, chemical and biological decontamination procedures during storage; and by avoiding damage to grain during distribution and processing.
These preventive measures might protect both animals and humans from mycotoxin contamination and also the contamination of food animal products that are consumed by humans. However, it is impossible to eliminate fungi and mycotoxins totally (CAST, 2003).
Strategies involving the sorting (Jemmali, 1990; Pasikatan and Dowell, 2001) and/or destruction of contaminated foods (Karlovsky, 1999; Scott, 1998) are also unrealistic, given their cost. The dilution of contaminated foods such that contamination falls below authorised levels has been forbidden in Europe since July 2003.
The lack of practical solutions to totally avoid mycotoxin contamination of feeds explains why inorganic materials such as clays, bentonites and aluminosilicates, known for their adsorptive properties, were at first proposed to step down the toxic effect of aflatoxins (Galvano et al., 2001; Grant and Phillips, 1998; Huwig et al., 2001; Lemke et al., 2001; Patterson and Young, 1993; Ramos and Hernandez, 1997), but they showed limited efficacy against other toxins.
Furthermore, they reduced the biological value of certain nutrients and could contain dioxin and heavy metals. Organic compounds such as yeast cell walls (Dawson et al., 2001; Devegowda et al., 1998a; Freimund et al., 2003; Newman, 2000) have therefore been proposed as alternative solutions to complex several mycotoxins without impairing nutrient bioavailability or inducing environmentally detrimental effects. They can be added to contaminated food or feeds to selectively bind mycotoxin, allowing the toxins to pass through the digestive tract without negative effects on animals or carryover to edible animal products (Dawson et al., 2001; Devegowda et al., 1998a; Devegowda et al., 1998b; Newman, 2000).
Numerous studies have evaluated the functions of ligands from yeast cell walls in vivo. Functional efficiency has been assessed indirectly, not by the characterisation of complex formation, but instead by determining appropriate biological markers in animals given foods naturally or artificially contaminated with mycotoxins (Devegowda et al., 1998a; Dvorska, 2003; Dvorska and Surai, 2001; Dvorska et al., 2003; Newman, 2000; Smith et al., 2001; Smith et al., 2002; Surai et al., 2002; Swamy et al., 2002a; Swamy et al., 2002b; Whitlow et al., 2000; Whitlow and Hagler, 2002; Whitlow and Hagler, 1999).
However, few studies have investigated the intrinsic mechanisms underlying the formation of complexes between toxins and yeast cell wall components (Dawson et al., 2001; Diaz et al., 2002). Only recent studies on inorganic ligands have looked at the chemical and quantitative aspects of these processes.
More analytical studies have been conducted on aluminosilicates. These involved the modeling of adsorption curves and the determination of physical factors, making it possible to calculate adsorption capacity, the number of sites involved, the strength of the interaction and thermodynamic parameters (Grant and Phillips, 1998; Lemke et al., 2001).
These advances led to the formulation of several hypotheses concerning the nature of the complexes formed between yeast cell walls and certain mycotoxins, raising the possibility of hydrophobic interactions in the case of zearalenone (ZEN) (Freimund et al., 2003). However, none of these studies described the finer mechanisms of the complexforming activity of these molecules.
Yeast cell wall organization
Yeasts, and particularly the cell wall of Saccharomyces cerevisiae, are an environmentally friendly alternative to inorganic adsorbents, which are not extensively biodegradable and are associated with the risk of contaminants.
The yeast cell wall fraction, which is mainly composed of polysaccharides (80-90%), makes up 15 to 30% of the dry weight of the whole cell. The mechanical strength of the wall is mainly due to the inner layer composed of ß-D-glucan chains (50-60% of the wall dry weight) made of ß-(1,3)-D-glucans with high complexity and degree of polymerisation (2 kDa for soluble forms up to 35 kDa for insoluble forms; Lee, 2001) and linear side chains of ß-(1,6)-D-glucans with low degree of polymerisation.
ß-D-glucans can be found under random coil or more organised conformations that constitute a network composed of single helix chains associated in triple helices more or less densely packed and nicely stabilised by inter- or intra-hydrogen bounds (Kogan, 2000). This fraction is firmly bound to the plasma membrane through linear chitin chains of N-acetylglucosamine (near to 190 units).
It contributes to the insolubility of the overall structure and the packing of ß-D-glucans, which both modulate the plasticity of the cell wall. Chitin molecules constitute 2 to 4% of the polysaccharides of the cell wall, and can reach 10% in some mutant laboratory strains. The outer layer is mainly composed of mannans (over 150 units of D-mannose), which are linked to ß-D-glucans through a GPI anchor or other protein-based links (Lipke and Ovalle, 1998).
This highly dynamic structure is strongly related to the strain nature because about 1200 genes drive its synthesis. The culture conditions such as pH, temperature, oxygenation rate, nature of the medium, concentration or nature of the carbon source, strongly modulate the amount and the structural properties of ß-D-glucans, mannans and chitin that are synthesised.
Moreover, the cell cycle stage also interacts with the cell wall composition due to budding; and thus due to the changes in the repartition of the structural components of the cell wall such as chitin.
What science is behind Mycosorb®?
The main goal of this study was to identify cell wall components responsible for the complexation of mycotoxins, more specifically of zearalenone (ZEN), known as a non-steroidal estrogenic mycotoxin, and to identify the chemical bonds involved in the formation of ‘mycotoxin-yeast cell wall’ complexes. The following aspects were dealt with:
1) methodological aspects of in vitro characterisation of the adsorption capacity of mycotoxins by yeast cell walls, cell wall extracts and pure components used as models of cell wall components; physical constants were used to characterise the nature and strength of the interactions;
2) spectroscopic studies (NMR, UV spectroscopy, X-ray diffraction) to assess the chemical interactions at the macromolecular level between the binder and mycotoxins; and
3) in silico analysis (molecular mechanics) of the toxin and ligand molecules carried out with the aim of modelling (3D) the complexes formed as a result of their interaction and to evaluate the stability of these complexes by calculating their potential energy. The nature of the chemical bonds within complexes was also studied.
MATERIALS AND METHODS
Yeast biomass production
Four strains of S. cerevisiae were grown in flasks containing a YPD medium (1% (w/v) yeast extract, 2% (w/v) bacteriological peptone and 2% (w/v) glucose) at 30°C, shaken at 200 rpm: the wild type wt292, the fks1 mutant type, the mnn9 mutant type, and sc1026 (Alltech Inc.). Cultures were stopped at 2 x 107 cells/mL.
Cell wall isolation and component characterisation
Cell walls were disrupted with glass beads and isolated (Dallies et al., 1998). Quantification of mannans and glucans was achieved by mannose and glucose analysis after 2N-H2SO4 hydrolysis at 100°C for 4 hrs. An enzymatic method was used to determine the chitin content (Popolo et al., 1997).
Alkali-extraction of yeast cell wall components Cell walls were fractionated by alkali extraction with 1 M NaOH and 0.5% NaBH4 for 24 hrs at 37°C under agitation (adapted from Fleet, 1991 and Catley, 1988).
The suspension was centrifuged (10,000 × g for 5 min) and the supernatant and pellet fractions were separated.
Supernatants were dialyzed (1:100, v/v) on celluloseester membranes (MWCO: 6 to 8,000) with 0.02 M Tris/HCl buffer (pH 7.4) for 16 hrs at 4°C with magnetic stirring. ß-D-glucans, which were separated from mannans using a concanavalin A sepharose column, were eluted with 0.02 M Tris/HCl buffer (pH 7.4), 0.5 M NaCl and stored at -20°C until use.
The pellets were thoroughly washed with 1 mL of 75% ethanol/10 mM Hepes buffer (pH 7.1) and then suspended in 2 mL of 0.1 M Tris/HCl buffer (pH 8.5) and finally stored at -20°C until use.
Quantitative analysis of carbohydrates Analyses were performed in triplicate on a Dionex Bio- LC system (Sunnyvale, CA) with a pulsed amperometric detector equipped with a gold electrode.
Separation of carbohydrates was performed on a CarboPac PA1 anion-exchange column (4 × 250 mm) equipped with a guard column. Elution was at a flow rate of 1 mL/min at 20-22°C with 18 mM NaOH.
Mycotoxin quantification
Mycotoxins were analyzed by isocratic HPLC on an HP-1090 Series II HPLC (Hewlett-Packard Co.) using a UV diode array detector coupled to an HP-1046A fluorescence detector. A C18 Nucleosil Spherisorb ODS-2 column (4 × 150 mm) equipped with a guard column was used at a flow rate of 0.8 mL/min of mobile phase.
In vitro technique to estimate the complex forming capacity
For each in vitro test, 100 µg/mL of adsorbent were placed in tubes together with 2, 4, 6, 8, 10 or 20 µg/mL of ZEN dissolved in water and agitated at 200 rpm for 1.5 hrs at 37°C, before centrifugation at 5,000 × g.
The amount of bound toxin was calculated by subtracting the amount of free toxin found in the supernatant of the experimental tubes from the amount found in control tubes with no adsorbent.
DataFit 7.1 software (©Oakdale) was used to plot the experimental data, fit the regression curve and calculate the statistical data (Yiannikouris et al., 2003).
Structural investigations of yeast cell wall components
Molecular modeling was conducted on Silicon Graphics computers with Accelrys packages (Accelrys. Inc, CA, USA) and InsightII, Biopolymer, Analysis, Docking and Discover modules. A CFF91 force-field adapted to polysaccharide/ligand interaction studies was used in vacuum condition (e = 1).
Construction of the most highly probable conformations of ß-(1,3)-D-glucan chain was performed as described in previous work (Yiannikouris et al., 2004a). Dihedral angle [φ, ψ] values of a ß-(1,3) bond between two glucose molecules were explored through their rotation (from –180° to +180°) and their minimum energy conformations evaluated before further elongation to a 5,868 Da polymer.
The structure of ß-D-glucans was assessed using X-ray diffraction under several hydration levels (Yiannikouris et al., 2004a) in order to see their influence on the packing of yeast cell wall components (Nuessli et al., 2003).
Characterisation of the chemical interactions between the binder and mycotoxins (Yiannikouris et al., 2004a).
Fifty µg/mL of ZEN and soluble purified ß-D-glucans were added to 10 mL of a Milli-Quf+ water, shaken at 640 rpm for 1.5 hrs at 39°C, and then cooled in a bath of 2-propanol kept at -30°C before freeze-drying. The powder containing the [ß-D-glucans + ZEN] complex was solubilized in 500 µL of D RSD = 0.534 µg/mL; A ? 30%). Cell walls of wt292 and mnn9 strains with higher levels of ß-D-glucans were able to complex larger amounts of ZEN with higher affinity rates than the fks1 and sc1026 strains.
High chitin content in mnn9 and fks1 strains increased the insolubility of ß-D-glucan and decreased the flexibility of the overall structure, which consequently limited the accessibility of ZEN to the chemical sites of the ß-D-glucan. Thus, these strains exhibited a lower complex-forming capacity than expected from their ß-D-glucan content (Yiannikouris et al., 2004b).
The alkali-insoluble fraction had a greater affinity (up to 50%) than the alkali-soluble fraction (about 16%).
From the results obtained with individual fractions or with HMN-3 plotting (R² = 0.969; RSD = 0.296 µg/ mL), we confirmed that mnn9 and wt292 strains (A = 35.9 and 50.4% for the alkali-insoluble fractions, respectively) had the highest ZEN adsorption efficacy (Figure 2). Partial elimination of chitin during alkali extraction improved by 1.5 times the capacity of the alkali-insoluble fraction of ß-D-glucans of mnn9 strain to adsorb ZEN compared to wt292 (Yiannikouris et al., 2004c).
Pure ß-D-glucans were used to discriminate the role played by ß-(1,3)-D-glucan and ß-(1,6)-D-glucan, respectively. ß-(1,3)-D-glucan were responsible for the main complexing activity since they had greater affinity than ß-(1,6)-D-glucan, but were more sensitive to environmental conditions such an increase of pH.
Also, the complexation abilities toward ZEN of some ß-D-glucans consisting of ß-(1,3)-D-glucan highly branched with ß-(1,6)-D-glucan were less sensitive to alkaline pH than ß-D-glucan made of pure chains of ß-(1,3)-D-glucan. We can conclude that branched ß-(1,6)-D-glucans stabilize the adsorption process of ZEN on ß-(1,3)-D-glucan.
Study of the interaction at macromolecular level
The case of ß-D-glucans
The alkali-insolubility of ß-D-glucan has a direct role in maintaining rigidity and integrity of the yeast cell wall. ß-D-glucan is thus defined as a complex 3D structure with alternating regions of random coil, single helices and triple helices (Kogan, 2000; Lipke and Ovalle, 1998).
In contrast, yeast cell walls with a high content of alkali-soluble ß-D-glucan are more flexible (Fleet, 1991; Kopecka et al., 1974).
Table 1.Models used to plot and evaluate ZEN adsorption on ß-D-glucans of yeast cell walls of S. cerevisiae according to Hill’s equation a.
To enlarge the imagen click here
a a = variable distinguishing the role played by alkali-insoluble compared with alkali-soluble ß-D-glucans; Insol = amount of alkali-insoluble ß-D-glucans (%); Sol = amount of alkali-soluble ß-D-glucans (%); KDtotal = association constant compared with amount of toxins initially added; Tbound = amount of bound toxin (ppm) ; Ttotal = amount of total toxin added (ppm); ![]() maximal amount of bound toxin (ppm).
maximal amount of bound toxin (ppm).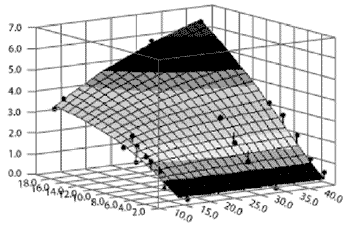
Figure 2.Graphic representation using HMN-3 of the relationship between [alkali-soluble + alkali-insoluble] ß-D-glucan content and the adsorption of ZEN.
X1 = amount of total ZEN added to the medium (µg/mL); X2 = [alkali-soluble + alkali-insoluble] ß-D-glucan content of the cell walls of the four strains tested (%); Z = amount of ZEN adsorbed by yeast cell walls (µg/mL).
A wide-angle X-ray diffraction study conducted on purified ß-D-glucan fractions indicated that they are spatially organised as a cluster of four triple-helix chains of ß-(1,3)-D-glucans (Chuah et al., 1983). These helical chains were 1.56 nm apart, with a fiber period of 0.60 nm, consequently defining six ß-D-glucopyranose units per turn of helix (Figure 3).
Such organisation was assessed by the comparison of pure molecules of ß-D-glucan such as paramylon, curdlan and laminarin, with reticular distances respectively equal to 1.36 (Chuah et al., 1983), 1.50 and 1.62 nm as obtained by X-ray diffraction measurements. The increase of the reticular distance between ß-D-glucan molecules involved an increase in the relaxation level of their triple helical conformation (Marchessault and Deslandes, 1979; Marchessault et al., 1977).
For example, the laminarin molecule that was used for further investigating the interaction with ZEN exhibited a highly relaxed triplehelix and/or single-helix organization relative to its very high reticular distance. These conformations and different degrees of relaxation might be found together in the highly complex structure of S. cerevisiae cell wall.
Molecular mechanics techiques confirmed that the most stable conformation of the ß-(1,3)-D-glucan single helical chain was found for a glycosidic linkage of [φ, ψ]=(-100°, 140°) with six ß-D-glucopyranose units per turn.
Two distinct low-energy populated conformations of ß-(1,6)-D-glucans were found for the dihedral angles [φ, ψ, ω] = (-87.7°, +179.9°, -72.6°) and [φ, ψ, ω] = (-85.5°,+ 179.7°, -177.9°), underlining the high flexibility of ß-(1,6)-D-glucan side chains, already reported in the literature (Kim et al., 2000; Manners et al., 1973; Stevens and Sathyanarayana, 1987).
Among the other conformations such as random coil, or densely packed triple helix that can be found in yeast cell wall, the single helix conformation elucidated for pure ß-D-glucan could be proposed as the structure that gave the highest number of potential sites for establishing an interaction with mycotoxins. In this respect, we investigated the ß-D-glucan single helix conformation for its complexing properties toward ZEN.
Macromolecular in vitro experiments and NMR experiments (Yiannikouris et al., 2003, 2004b; 2004c; 2004d) established that environmental conditions such as pH and nature of the solvent can decrease the stability of the geometry of ß-D-glucans, and thus decrease their ability to complex ZEN.
Such results indicate that weak non-covalent bonds might be involved, the interactions between ß-D-glucans and ZEN becoming therefore more of an ‘adsorption’ type rather than a ‘binding’ type. 1H-NMR investigations showed that the interaction of the ZEN and ß-D-glucans strongly reduced the two hydroxyl peaks of the ZEN phenol moiety, which thus implicated hydrogen bonds.
Molecular mechanics showed that the ß-(1,3)-D-glucan chain favored a very stable intra-helical association with ZEN (Figures 4 and 5).
Two types of bonds were precisely identified in monitoring the association:
1) ZEN hydroxyl, ketone and lactone groups were involved in hydrogen bonds with ß-(1,3)-D-glucans;
2) a stabilizing van der Waals electronic interaction occurred due to the geometrical symmetry and to the proximity between ß-D-glucopyranose rings and ZEN phenol moiety.
Therefore, the ZEN molecule is totally trapped inside the ß-D-glucan structure (Yiannikouris et al., 2004a).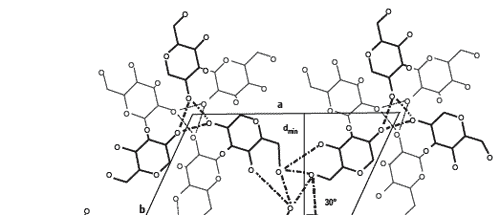
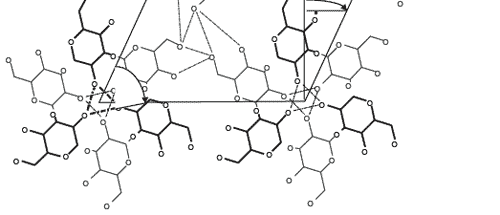
Figure 3.Scheme of the projection following a and b axes of the hexagonal crystal made of 4 x [3 dimer of ß-D-glucopyranose organized in triples helices] (Chuah et al., 1983; Pelosi, 2002). Glucopyranose residues in black are forward to the plane a, b and the residues in grey are backward to the plane a, b. Stabilisation of the helix involved intra and inter-helical hydrogen bonds. dmin = 1.36 nm ; a = b = 1.56 nm; fiber period c = 0.60 nm; ? = 120.
Differences between mycotoxins
Not all mycotoxins are bound in the same manner. By means of molecular modeling, we defined the first molecular indices of the stability of binding to ß-D-glucans for aflatoxin B1 (AFB1), patulin (PAT) and deoxynivalenol (DON). Both AFB1 and ZEN display geometric similarity to the helix of ß-(1,3)-D-glucans.
The aromatic ring and lactone groups of AFB1 form polar or electron bonds with the single helix of ß-D-glucans. The small size of PAT enables it to penetrate deep within the helix of ß-(1,3)-D-glucans.
Hydrogen bonds are formed between ß-D-glucans and lactone and hydroxyl groups from PAT. The oxygen atom of the side group can also form electron doublets with the hydroxyl groups of the ß-(1,3)-D-glucan helix.
No geometrical similarities were found between the DON molecule and the single helix of ß-D-glucan by comparison with the ZEN or AFB1 molecule, leading to a decrease of the contribution due to van der Waals.
However, DON was able to form at least two hydroxyl bounds with the ß(1,3)-D-glucan molecule.
These results enabled us, for the first time, to characterise the chemical mechanism of the interaction between mycotoxins and ß-D-glucans at the molecular level.
Although this work also demonstrates the difficulties involved in extending our conclusions to all mycotoxins, we can nonetheless identify certain chemical structures of interest with a high probability of interacting with ß-D-glucans.
All aflatoxins, citrinin, ergotamine, T-2 toxin, paspalitrems, slaframine and verrucarin probably form bonds with ß-D-glucans due to their ‘aflatoxin-like’, ‘deoxynivalenol-like’ or ‘zearalenone-like’ structures. For example, T-2 toxin has an affinity of about 33%, as determined with Mycosorb®, according to published data (Devegowda et al., 1998a; Devegowda et al., 1998b).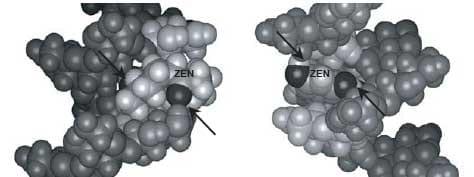
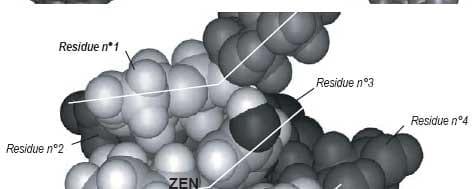
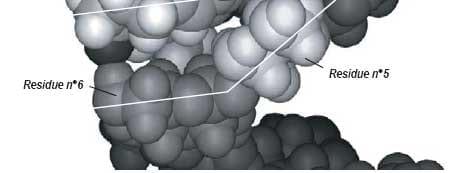
Figure 4. Computer-generated views with the Insight II program of the energy-minimized structure of the docking of the most favorable conformation of ZEN (B) into the single-helix of ß-(1,3)-D-glucan chain. Arrows indicate hydrogen bonds involved in the interaction. Lines highlight the steric complementarity between ZEN and ß-(1,3)-D-glucan geometry.
| Conclusions This work underlines the major role played by ß-D-glucan of yeast cell wall and its 3D complex and densely organised structure in the adsorption of ZEN. Interestingly, the geometrical, the electrostatic and the hydrophobic complementarities between a single helix of ß-D-glucan and ZEN have been characterized. As far as we know, this is the first study that elucidates how ß-D-glucan chains, and moreover Mycosorb®, have specific and quantitative complexing properties toward ZEN. The use of this organic material derived from yeast cell walls (Dawson et al., 2001; Devegowda et al., 1998b; Newman, 2000), could thus help to mitigate the harmful impact of mycotoxins on animals and overcome the reduction in bioavailability of some nutrients and their non-biodegradation in the environment. Finally, we describe an original method that can objectively distinguish between yeasts according to their ability to adsorb a mycotoxin. We also elucidated the mechanisms involved in the formation of complexes between ZEN and yeast cell walls, and we suggest molecular models describing the structures of the complexes formed and their stability, in the digestive tract in particular. The nature of the chemical bonds formed between ZEN and yeast cell wall components was determined. |
Acknowledgments
This work was co-supported (CIFRE No. 704/2000) by Alltech Inc., ANRT (Association Nationale de la Recherche Technique) and INRA.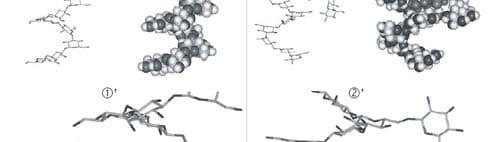
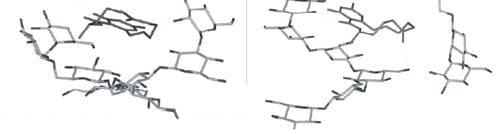
Figure 5. Computer-generated views with the Insight II program of the energy-minimized structure of the docking of the most favorable conformation of ZEN (B) into the single-helix of ß-(1,3)-D-glucan chain branched with ß-(1,6)-D-glucan side chain, in two conformations ( 1 and 2).
We especially thank G. Jeminet and I. Canet (UMRCNRS 6504, SEESIB, Université Blaise-Pascal Clermont II, Aubière, France), J. François (UMR-CNRS 5504, UR-INRA 792, INSA Toulouse, France), L. Poughon & C.-G. Dussap (CUST, Université Blaise- Pascal Clermont II Aubière, France), G. André (Institut Pasteur, Paris, France), A. Buléon and B. Pontoire (UR783 INRA, UPCM, Nantes, France), X. Cameleyre (Critt Bio-Industries, INSA, Toulouse, France), P. Galtier (INRA-UR66, Toulouse, France) for their helpful support and personal involvement in this research.
References
CAST. 2003. In: Mycotoxins: Risks in Plant, Animal, and Human Systems. Council for Agricultural Science and Technology (K. Niyo, ed.). Task Force Report 139, Ames, Iowa, USA, pp. 1-199.
Catley, B.J. 1988. Isolation and analysis of cell walls. Chapter 8. In: Yeast: A Practical Approach (T. Campbell and J. H. Duffus, eds). IRL Press, Washington DC, USA, pp. 163-183.
Chuah, C.T., A. Sarko, Y. Deslandes and R.H. Marchessault. 1983. Packing analysis of carbohydrates and polysaccharides. Part 14. Triple-helical crystalline structure of curdlan and paramylon hydrates. Macromolecules 16:1375-1382.
Cordier, C., M. Gruselle, G. Jaouen, D.W. Hughes and M.J. McGlinchey. 1990. Structures of zearalenone and zearalanone in solution: a high-field NMR and molecular modelling study. Magn. Reson. Chem. 28:835-845.
Dallies, N., J. François and V. Paquet. 1998. Yeast functional analysis reports: a new method for quantitative determination of polysaccharides in the yeast cell wall. Application to the cell wall defective mutants of Saccharomyces cerevisiae. Yeast 14:1297- 1306.
Dawson, K.A., J. Evans and M. Kudupoje. 2001. Understanding the adsorption characteristics of yeast cell wall preparations associated with mycotoxin binding. In: Science and Technology in the Feed Industry, Proceedings of Alltech’s 17th Annual Symposium (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, UK, pp. 169-181.
Devegowda, G., M.V.L.N. Raju, N. Afzali and H.V.L.N. Swamy. 1998a. Mycotoxin picture world-wide: novel solutions for their counteraction. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 14th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 241-256.
Devegowda, G., M.V.L.N. Rayu and H.V.L.N. Swamy. 1998b. Mycotoxins: novel solutions for their counteraction. Feedstuffs 7:12-15.
Diaz, D.E., W.M. Hagler Jr, B.A. Hopkins and L.W. Whitlow. 2002. Aflatoxin binders I: in vitro binding assay for aflatoxin B1 by several potential sequestering agents. Mycopathologia 156:223-226.
Dvorska, J. 2003. Aurofusarin, a newly described Fusarium graminearum mycotoxin: oxidative stress and protective effect of Mycosorb®. In: Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s 19th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 93-106.
Dvorska, J.E. and P.F. Surai. 2001. Effects of T-2 toxin, zeolite and Mycosorb® on antioxidant systems of growing quail. Asian-Austr. J. Anim. Sci. 14:1752- 1757.
Dvorska, J.E., P.F. Surai, B.K. Speake and N.H.C. Sparks. 2003. Protective effect of modified glucomannans against aurofusarin-induced changes in quail egg and embryo. Comp. Biochem. Physiol. C 135: 337-343.
Fleet, G.H. 1991. Cell walls. Chapter 5. In: The Yeasts, 4. (A.H. Rose and J.S. Harrison, eds). Academic Press, London, UK, pp. 199-271.
Freimund, S., M. Sauter and P. Rys. 2003. Efficient adsorption of the mycotoxins zearalenone and T-2 toxin on a modified yeast glucan. J. Environ. Sci. Health B 38:243-255.
Galvano, F., A. Piva, A. Ritieni and G. Galvano. 2001. Dietary strategies to counteract the effects of mycotoxins: a review. J. Food Prot. 64:120-131.
Grant, P.G. and T.D. Phillips. 1998. Isothermal adsorption of aflatoxin B1 on HSCAS clay. J. Agric. Food Chem. 46:599-605.
Guerre, P., J-D. Bailly, G. Bernard and V. Burgat. 2000. Excrétion lactée des mycotoxines : quels risques pour le consommateur? Rev. Med. Vet. 151:7-22.
Huwig, A., S. Freimund, O. Käppeli and H. Dulter. 2001. Mycotoxin detoxification of animal feed by different adsorbents. Toxicol. Lett. 122:179-188.
Jemmali, M. 1990. Decontamination and detoxification of mycotoxins. J. Env. Pathol. Toxicol. Oncol. 10:154-159.
Karlovsky, P. 1999. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Nat. Toxins 7:1-23.
Kim, Y-T., E-H. Kim, C. Cheong, D.L. Williams, CW. Kim and S-T. Lim. 2000. Structural characterization of ß-(1,3)-, ß-(1,6)-linked glucans using NMR spectroscopy. Carbohydr. Res. 328:331- 341.
Kogan, G. 2000. Bioactive natural products (part D): ß-(1,3)- and ß-(1,6)-D-glucans of yeasts and fungi and their biological activity. In: Studies in Natural Products Chemistry, 23. Atta-ur-Rahman, ed. Elsevier Science BV, Karachi, Pakistan, pp. 107-151.
Kopecka, M., H.J. Phaff and G.H. Fleet. 1974. Demonstration of a fibrillar component in the cell wall of the yeast Saccharomyces cerevisiae and its chemical nature. J. Cell Biol. 62:66-76.
Krishnamachari, K.A., V.R. Bhat, V. Nagarajan, T.B. Tilak and P.G. Tulpule. 1977. The problem of aflatoxic human disease in parts of India-epidemiological and ecological aspects. Ann. Nutr. Aliment. 31:991-996.
Lee, I.-Y. 2002. Curdlan, p. 135-154. In E. Vandamme, S. De Baets, and A. Steinbüchel (ed.), Biopolymers, vol. 5. Wiley, Hoboken, N.J.
Lemke, S.L., S.E. Ottinger, K. Mayura, C.L. Ake, K. Pimpukdee, N. Wang and T.D. Phillips. 2001. Development of a multi-tiered approach to the in vitro prescreening of clay-based enterosorbents. Anim. Feed Sci. Technol. 93:17-29.
Lipke, P.N. and R. Ovalle. 1998. Cell wall architecture in yeast: new structure and new challenges. J. Bacteriol. 180:3735-3740.
Manners, D.J., A.J. Masson, J.C. Patterson, H. Björndal and B. Lindberg. 1973. The structure of a ß-(1,6)-Dglucan from yeast cell walls. Biochem. J. 135:31-36.
Marchessault, R.H., Y. Deslandes, K. Ogawa and P.R. Sundararajan. 1977. X-Ray diffraction data for ß (1,3)-D-glucan. Can. J. Chem. 55:300-303. Marchessault, R.H. and Y. Deslandes. 1979. Fine structure of ß-(1,3)-D-glucans: curdlan and paramylon. Carbohydr. Res. 75:231-242.
Newman, K. 2000. The biochemistry behind esterified glucomannans - titrating mycotoxins out of the diet. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 16th Annual Symposium (T. P. Lyons and K. A. Jacques, eds.). Nottingham University Press, UK, pp 369-382.
Nuessli, J., J.L. Putaux, P.L. Bail and A. Buleon. 2003. Crystal structure of amylose complexes with small ligands. Int. J. Biol. Macromol. 33:227-234.
Pasikatan, M.C. and F.E. Dowell. 2001. Sorting systems based on optical methods for detecting and removing seeds infested internally by insects or fungi: a review. Appl. Spectrosc. Rev. 36:399-416.
Patterson, R. and L.G. Young. 1993. Efficacy of hydrated sodium calcium aluminosilicate, screening and dilution in reducing the effects of mold contaminated corn in pigs. Can. J. Anim. Sci. 73:615- 624.
Pelosi, L. 2002. Caractérisation des ß-(1,3)-D-glucane synthases de Saprolegnia monoica et de leurs produits de synthèse, Université Joseph Fourier, Grenoble I. Pfohl-Leszkowicz, A. 2000. Risques mycotoxicologiques pour la santé des animaux et de l’Homme. Cah. Nutr. Diet. 35:389-401.
Pittet, A. 1998. Natural occurrence of mycotoxins in foods and feeds - an updated review. Rev. Med. Vet. 149:479.
Popolo, L., D. Gilardelli, P. Bonfante and M. Vai. 1997. Increase in chitin as an essential response to defects in assembly of cell wall polymers in the ggp1delta mutant of Saccharomyces cerevisiae. J. Bacteriol. 179:463- 469.
Ramos, A.J. and E. Hernandez. 1997. Prevention of aflatoxicosis in farm animals by means of hydrated sodium calcium aluminosilicate addition to feedstuffs: a review. Anim. Feed Sci. Technol. 65:197-206.
Schoental, R. 1984. Mycotoxins and the Bible. Perspect. Biol. Med. 28:117-120.
Schoental, R. 1991. Mycotoxins, porphyrias and the decline of the Etruscans. J. Appl. Toxicol. 11:453- 454.
Scott, P.M. 1998. Industrial and farm detoxification processes for mycotoxins. Rev. Med. Vet. 149:543- 548.
Smith, T.K., E.J. MacDonald and S. Haladi. 2001. Current concepts in feed-borne mycotoxins and the potential for dietary prevention of mycotoxicoses. In: Science and Technology in the Feed Industry, Proceedings of Alltech’s 17th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 183-189.
Smith, T.K., H.V.L.N. Swamy, S.L. Raymond and M. Zaytoun. 2002. Contemporary perspectives on Fusarium mycotoxicoses in livestock and poultry. In: Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s 18th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 373-378.
Stevens, E.S. and B.K. Sathyanarayana. 1987. Conformation of ß-(1,6)-D-glucan by vacuumultraviolet circular dichroism. Carbohydr. Res. 160:131-135.
Surai, P.F., J.E. Dvorska, N.H.C. Sparks and K.A. Jacques. 2002. Impact of mycotoxins on the body’s antioxidant defence. In: Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s 18th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 131-141.
Swamy, H.V.L.N., T.K. Smith, P.F. Cotter, H.J. Boermans and A.E. Sefton. 2002a. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on production and metabolism in broilers. Poult. Sci. 81:966-975.
Swamy, H.V.L.N., T.K. Smith, E.J. MacDonald, H.J. Boermans and E.J. Squires. 2002b. Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on swine performance, brain regional neurochemistry, and serum chemistry and the efficacy of a polymeric glucomannan mycotoxin adsorbent. J. Anim. Sci. 80:3257-3267.
Whitlow, L.W. and W.M.J. Hagler. 1999. An association of mycotoxins with production, health and reproduction in dairy cattle and guidelines for prevention and treatment. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 15th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 401-419.
Whitlow, L.W., D.E. Diaz, B.A. Hopkins and W.M. Hagler Jr. 2000. Mycotoxins and milk safety: the potential to block transfer to milk. In: Biotechnology in the Feed Industry, Proceedings of Alltech’s 16th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, UK, pp. 391-408.
Whitlow, L.W. and W.M. Hagler Jr. 2002. Mycotoxins in feeds. Feedstuffs 68-78.
Yiannikouris, A. and J-P. Jouany. 2002. Les mycotoxines dans les aliments des ruminants, leur devenir et leurs effets chez l’animal. INRA Prod. Anim. 15:3-16.
Yiannikouris, A., L. Poughon, X. Cameleyre, C.-G. Dussap, J. François, G. Bertin and J.-P. Jouany. 2003. A novel technique to evaluate interactions between Saccharomyces cerevisiae cell wall and mycotoxins: application to zearalenone. Biotechnol. Lett. 25:783- 789.
Yiannikouris, A., G. André, A. Buléon, G. Jeminet, I. Canet, J. François, G. Bertin and J-P. Jouany. 2004a. Comprehensive conformational study of key interactions involved in zearalenone complexation with ß-D-glucans. Biomacromolecules 5:2176-2185.
Yiannikouris, A., J. François, L. Poughon, C-G. Dussap, G. Bertin, G. Jeminet and J-P. Jouany. 2004b. Adsorption of zearalenone by ß-D-glucans in the Saccharomyces cerevisiae cell wall. J. Food Prot. 67:1195-1200.
Yiannikouris, A., J. François, L. Poughon, C-G. Dussap, G. Bertin, G. Jeminet and J-P. Jouany. 2004c. Alkaliextraction of ß-D-glucans from Saccharomyces cerevisiae cell wall and study of their adsorptive properties toward zearalenone. J. Agric. Food Chem. 52:3666-3673.
Yiannikouris, A., J. François, L. Poughon, C-G. Dussap, G. Jeminet, G. Bertin and J-P. Jouany. 2004d. Influence of pH on complexing of model ß-D-glucans with zearalenone. J. Food Prot. 67:2741–2746.
Authors: ALEXANDROS YIANNIKOURIS1, GÉRARD BERTIN1 and JEAN-PIERRE JOUANY2
1 Alltech France, Goussainville, France,
2 INRA, Unité de Recherches sur les Herbivores, Centre de Clermont-Theix, St. Genés, Champanelle, France





.jpg&w=3840&q=75)