Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs
Published: September 16, 2014
By: Veronika Nagl a, Bettina Woechtl b, Heidi Elisabeth Schwartz-Zimmermann a, Isabel Hennig-Pauka b, Wulf-Dieter Moll c, Gerhard Adam d, Franz Berthiller a
aChristian Doppler Laboratory for Mycotoxin Metabolism, Center for Analytical Chemistry, Department for Agrobiotechnology (IFA-Tulln), University of Natural Resources and Life Sciences, Vienna (BOKU), Konrad Lorenz Str. 20, Tulln 3430, Austria
bUniversity Clinic for Swine, Department for Farm Animals and Veterinary Public Health, University of Veterinary Medicine Vienna, Veterinärplatz 1, Vienna 1210, Austria
cBiomin Research Center, Technopark 1, Tulln 3430, Austria
dDepartment of Applied Genetics and Cell Biology, University of Natural Resources and Life Sciences, Vienna (BOKU), Konrad Lorenz Str. 24, Tulln 3430, Austria
Toxicology Letters 229 (2014) 190–197. Published by Elsevier Ireland Ltd. DOI: 10.1016/j.toxlet.2014.06.032
Article history:
Received 22 April 2014
Received in revised form 18 June 2014
Accepted 20 June 2014
Available online 23 June 2014
bUniversity Clinic for Swine, Department for Farm Animals and Veterinary Public Health, University of Veterinary Medicine Vienna, Veterinärplatz 1, Vienna 1210, Austria
cBiomin Research Center, Technopark 1, Tulln 3430, Austria
dDepartment of Applied Genetics and Cell Biology, University of Natural Resources and Life Sciences, Vienna (BOKU), Konrad Lorenz Str. 24, Tulln 3430, Austria
Toxicology Letters 229 (2014) 190–197. Published by Elsevier Ireland Ltd. DOI: 10.1016/j.toxlet.2014.06.032
Article history:
Received 22 April 2014
Received in revised form 18 June 2014
Accepted 20 June 2014
Available online 23 June 2014
Abstract
Plants can metabolize the Fusarium mycotoxin deoxynivalenol (DON) by forming the masked mycotoxin deoxynivalenol -3-ß-D-glucoside (D3G). D3G might be cleaved during digestion, thus increasing the total DON burden of an individual. Due to a lack of in vivo data, D3G has not been included in the various regulatory limits established for DON so far. The aim of our study was to contribute to the risk assessment of D3G by determination of its metabolism in pigs. Four piglets received water, D3G (116 μg/kg b.w.) and the equimolar amount of DON (75 μg/kg b.w.) by gavage on day 1, 5 and 9 of the experiment, respectively. Additionally, 15.5 μg D3G/kg b.w. were administered intravenously on day 13. Urine and feces were collected for 24 h and analyzed for DON, D3G, deoxynivalenol-3-glucuronide (DON-3-GlcA), deoxynivalenol-15-GlcA (DON-15-GlcA) and deepoxy-deoxynivalenol (DOM-1) by UHPLC–MS/MS. After oral application of DON and D3G, in total 84.8 ± 9.7% and 40.3 ± 8.5% of the given dose were detected in urine, respectively. The majority of orally administered D3G was excreted in form of DON, DON-15-GlcA, DOM-1 and DON-3-GlcA, while urinary D3G accounted for only 2.6 ± 1.4%. In feces, just trace amounts of metabolites were found. Intravenously administered D3G was almost exclusively excreted in unmetabolized form via urine. Data indicate that D3G is nearly completely hydrolyzed in the intestinal tract of pigs, while the toxin seems to be rather stable after systemic absorption. Compared to DON, the oral bioavailability of D3G and its metabolites seems to be reduced by a factor of up to 2, approximately.
Abbreviations
3-ADON, 3-acetyl-deoxynivalenol; 15-ADON, 15-acetyl-deoxynivalenol; b.w., body weight; D3G, deoxynivalenol-3-β-d-glucoside; DON, deoxynivalenol; DON-3-GlcA, deoxynivalenol-3-glucuronide; DON-15-GlcA, deoxynivalenol-15-glucuronide; DON-GlcA, deoxynivalenol-glucuronide; DOM-1, de-epoxy deoxynivalenol; DOM-1-GlcA, DOM-1-glucuronide; JECFA, Joint FAO/WHO Expert Committee on Food Additives; MeOH, methanol; LOD, limit of detection; LOQ, limit of quantification; MS, mass spectrometry; MS/MS, tandem mass spectrometry; PBS, phosphate buffered saline; RA, apparent recovery; RE, recovery of the extraction step; SSE, signal suppression/enhancement; UHPLC, ultra high performance liquid chromatography
Keywords: Conjugated mycotoxins; ADME; Swine; Gut
3-ADON, 3-acetyl-deoxynivalenol; 15-ADON, 15-acetyl-deoxynivalenol; b.w., body weight; D3G, deoxynivalenol-3-β-d-glucoside; DON, deoxynivalenol; DON-3-GlcA, deoxynivalenol-3-glucuronide; DON-15-GlcA, deoxynivalenol-15-glucuronide; DON-GlcA, deoxynivalenol-glucuronide; DOM-1, de-epoxy deoxynivalenol; DOM-1-GlcA, DOM-1-glucuronide; JECFA, Joint FAO/WHO Expert Committee on Food Additives; MeOH, methanol; LOD, limit of detection; LOQ, limit of quantification; MS, mass spectrometry; MS/MS, tandem mass spectrometry; PBS, phosphate buffered saline; RA, apparent recovery; RE, recovery of the extraction step; SSE, signal suppression/enhancement; UHPLC, ultra high performance liquid chromatography
Keywords: Conjugated mycotoxins; ADME; Swine; Gut
1. Introduction
Mycotoxins, secondary metabolites of different molds, are frequent contaminants of food and feed and cause various adverse health effects in plants, animals and humans. Deoxynivalenol (DON), mainly produced by Fusarium graminearum and Fusarium culmorum, is one of the economically most relevant mycotoxins worldwide (CAST, 2003). DON exhibits its biological activity by binding to the 60S subunit of eukaryotic ribosomes with subsequent inhibition of the protein biosynthesis (summarized by Arunachalam and Doohan, 2013). In animals, DON induces clinical symptoms like anorexia, emesis, weight loss or immunomodulation (reviewed by Pestka, 2007), while the mycotoxin is associated with outbreaks of gastroenteritis in humans (summarized by Pestka, 2010). As a consequence, regulatory limits in cereal-based food have been established for DON e.g., by the European Commission (European Commission, 2006).
In contrast, deoxynivalenol-3-β-D-glucoside (D3G) has not been included in these regulations so far. This masked mycotoxin is formed in plants by enzymatic conjugation of glucose to DON (Poppenberger et al., 2003), thus representing an important defense mechanism of plants against Fusarium related diseases (Lemmens et al., 2005). D3G can be found in different cereal crops (Berthiller et al., 2005; Lancova et al., 2008), as well as in animal feed and foodstuff (De Boevre et al., 2012; Malachova et al., 2011). In grains, the molar D3G/DON ratio varies strongly in dependence on factors like genotype or season, but typically reaches levels of around 20% (summarized by Berthiller et al., 2013). However, this ratio can further increase during food processing, resulting in D3G concentrations even exceeding those of DON (Varga et al., 2013).
Despite its frequent occurrence, toxicological data regarding D3G are still rare. Poppenberger et al. (2003) showed that D3G inhibits protein biosynthesis to a far lower extent than its parent toxin in wheat ribosomes. Nevertheless, there are major concerns that ingested D3G might be hydrolyzed in the intestinal tract of mammals, thus releasing its aglycon and increasing the total DON load of an individual (Joint FAO/WHO Expert Committee on Food Additives, JECFA, 2011). Although the toxin seems to be rather stable under conditions simulating early stages of the mammalian digestion (Berthiller et al., 2011; De Nijs et al., 2012), certain lactic acid bacteria were capable to cleave D3G in vitro (Berthiller et al., 2011). Furthermore, incubation of human fecal samples with D3G resulted in an extensive liberation of DON (Dall´Erta et al., 2013; Gratz et al., 2013). In 2012, the first in vivo reports on the fate of orally administered D3G in rodents became available (Nagl et al., 2012; Veršilovskis et al., 2012). Our group demonstrated that D3G is hydrolyzed in the digestive tract of rats. The liberated DON was further metabolized, but only poorly absorbed. We therefore concluded that D3G is of considerably lower toxicological relevance than DON in rats (Nagl et al., 2012). However, for risk analysis of D3G more data regarding the toxicity and metabolism of this masked mycotoxin are urgently needed (JECFA, 2011).
The situation is quite different for DON, whose toxic effects have been investigated for decades. Notably, major differences in the susceptibility to DON among animal species were determined, which are most likely related to species-dependent variations in the metabolism of DON (summarized by Pestka, 2007). In principle, ingested DON can be either de-epoxidated by anaerobic bacteria or conjugated to glucuronic acid by mammalian UDP- glucuronosyltransferases (UGTs). While the formation of de-epoxy deoxynivalenol (DOM-1) by ruminal or intestinal microbiota occurs prior to systemic absorption, glucuronidation of DON has so far only been described for liver microsomes (reviewed by Maresca, 2013). Both metabolites, DOM-1 as well as DONglucuronide (DON-GlcA) are less cytotoxic than their parent toxin (Sundstøl Eriksen et al., 2004; Wu et al., 2007). In recent years, highly sophisticated analytical methods elucidated the metabolic pattern of DON in more detail. For example, the species-specific formation of different DON-GlcA isomers, namely DON-3-glucuronide (DON-3-GlcA), DON-15-glucuronide (DON-15-GlcA), DON- 7-glucuronide and DON-8-glucuronide has been proposed (Maul et al., 2012; Uhlig et al., 2013; Warth et al., 2013). Furthermore, the occurrence of novel DON metabolites has been revealed in rats and chickens (Wan et al., 2014). Generally, pigs are considered to be the species most sensitive to DON exposure. This might be attributed to the species’ poor ability to detoxify DON to DOM-1 and the high absorption rate of unmetabolized DON (Pestka, 2007). As these characteristics are very similar to what is currently known about human DON metabolism (reviewed by Turner et al., 2012), swine can be regarded as most suitable animal model in this research field.
The aim of our study was to elucidate whether D3G has the potential to be reactivated in vivo by investigation of its metabolism in pigs. For this purpose, piglets were orally and intravenously dosed with D3G and their excreta subsequently analyzed for DON, D3G, DON-3-GlcA, DON-15-GlcA and DOM-1 by a validated LC–MS/MS based biomarker method. The implementation of an additional oral DON treatment allowed a comparison between absorption rates of D3G and its parent toxin. This study significantly extends the current knowledge about D3G metabolism in vivo and therefore facilitates an estimation of the potential health risk of this masked mycotoxin.
2. Materials and methods
2.1. Chemicals and reagents
Methanol (MeOH, LC grade) and glacial acetic acid (p.a.) were obtained from VWR International GmbH (Vienna, Austria), while ammonium acetate (MS grade) and β-glucuronidase (Escherichia coli, Type IX-A) were purchased from Sigma–Aldrich (Vienna, Austria). Reagents for preparation of phosphate buffered saline (PBS; sodium chloride, disodium hydrogen phosphate dihydrate, sodium dihydrogen phosphate monohydrate) were from Merck GmbH (Vienna, Austria). Water was purified using a Purelab Ultra system (ELGA LabWater, Celle, Germany).
DON and DOM-1 standards were purchased from Romer Labs GmbH (Tulln, Austria). D3G was either purified from DON treated wheat as previously described (Berthiller et al., 2005), or generated by in vitro enzymatic synthesis and preparative HPLC purification (manuscript in preparation). DON-3-GlcA was produced by chemical synthesis (Fruhmann et al., 2012). A combined multi standard stock solution, containing 50 μg/mL DON, D3G, DON-3- GlcA and DOM-1, was prepared in MeOH and stored at -20ºC. Further dilutions for spiking experiments and calibrants were prepared in MeOH/water (20/80, v/v).
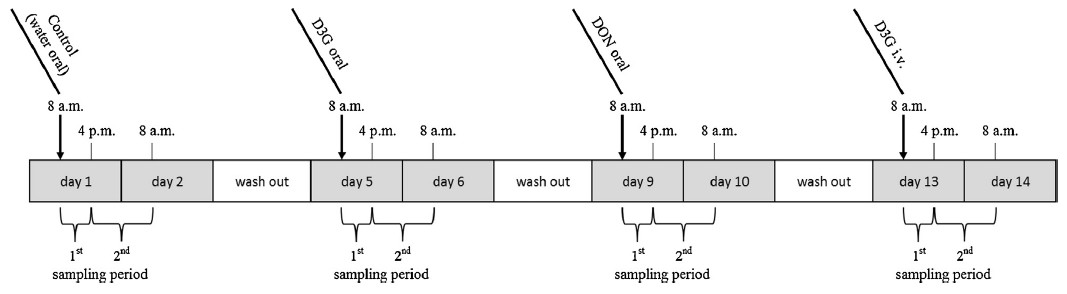
Fig. 1. Experimental design. After an acclimatization period of five days, four animals received water, D3G and DON by gavage on day 1, 5, and 9, respectively. On day 13, D3G was administered intravenously. After each of the treatments, animals were separated and urine and feces were collected in two different time periods (0–8 h, 8–24 h after administration).
2.2. Animals and study design
Male crossbred piglets (sow: Landrace x Large White, boar: Pietrain; 28 days old, 7.1 ±0.9 kg, non-castrated) were obtained from a commercial swine producer and allowed to acclimatize for five days. Animals were housed pairwise in metabolic cages and had free access to water. Feed was provided restrictively twice per day (5% of body weight/day) to ensure fast uptake of the diet. Before the start of the experiment, the diet was tested for its concentrations of DON, D3G and acetylated forms of DON (3- acetyl-deoxynivalenol, 3-ADON; 15-acetyl-deoxynivalenol, 15-ADON).
Using a repeated measures design, the piglets (n = 4) received water (negative control), D3G (116 μg/kg b.w.) and the equimolar dose (0.25 μmol/kg b.w.) of DON (75 μg/kg b.w.) per gavage on day 1, 5 and 9 of the experiment, respectively. In addition, D3G was administered intravenously (15.5 μg/kg b.w., 0.03 μmol/kg b.w.) on day 13 (Fig. 1). Stock solutions of D3G (1.2 μg/mL) and DON (1 mg/mL) were prepared by dissolving the solid standards in water (oral application) or physiological saline (0.9% NaCl, Fresenius Kabi, Graz, Austria; intravenous application). Volumes between 700 μL and 1000 μL were administered orally using polyvinyl chloride catheters (diameter 4.7 mm, length adapted to size of piglets, Medinorm, Spießen, Germany). Intravenous administration was performed by cannulation of the ear vein with sterile catheters (0.9 x 25 mm, VasoVet distributed by B. Braun, Melsungen, Germany). After each of the treatments, animals were separated and urine and feces of individual piglets were collected for 0–8 h and 8–24 h after dosing. Samples were frozen and stored at -20ºC until further analysis.
The animal experiment was approved by the Institutional Ethics Committee and the national authority according to §8ff of the Austrian Law for Animal Experiments, Tierversuchsgesetz – TVG (GZ 68.205/0236-II/3b/2012).
2.3. Sample preparation
For preparation of urine samples, a dilute-and-shoot approach was used. After centrifugation (10 min, 14,000 x g), 20 μL of urine samples were added to 180 μL of MeOH/water (20/80, v/v) and vortexed for 15 s. Thereof, 5 μL were injected into the LC–MS/MS system. In addition, some samples were enzymatically hydrolyzed to investigate the presence of DOM-1-glucuronide (DOM-1-GlcA). To this end, 180 μL of PBS (75 mM, pH 7.4) containing 9000 units of β-glucuronidase were mixed with 20 μL of centrifuged urine samples. After incubation for 18 h at 37 ºC, digested samples were clarified by centrifugation (10 min, 14,000 x g) for LC–MS/MS analysis.
Clean-up of feces samples was performed according to Nagl et al. (2012). In brief, 250 mg of freeze-dried and homogenized samples were extracted three times (40/40/20 min) with MeOH/ water (50/50, v/v, 3/2/2 mL). Following protein precipitation with cold MeOH, aliquots were evaporated and reconstituted in MeOH/ water (20/80, v/v, corresponding to a final sample to solvent ratio of 1:56). Afterwards, samples were centrifuged (10 min,14,000 x g) and 5 μL of the supernatant were injected into the LC–MS/MS system.
2.4. UHPLC–MS/MS parameters
Analyses were carried out on a 1290 Infinity series UHPLC system (Agilent Technologies, Waldbronn, Germany) coupled to a 4000 QTrap mass spectrometer (AB Sciex, Foster City, CA) equipped with a Turbo V electrospray ionization (ESI) source. Chromatographic separation was carried out on a Zorbax RRHD Eclipse XDB-C18 column (2.1 mm x 150 mm, 1.8 mm, Agilent Technologies, Waldbronn, Germany) equipped with a C18 security guard cartridge (Phenomenex, Aschaffenburg, Germany). Eluent A consisted of water and eluent B of MeOH/water (97/3, v/v), both containing 0.01% acetic acid and 5 mM ammonium acetate. After an initial period of 2 min at 5% B, the proportion of B was increased linearly to 100% (reached at 6.4 min), followed by a hold time of 2 min at 100% B. Afterwards, the column was re-equilibrated for 2 min at 5% B. The temperature of the column oven was set to 35 ºC, while 5 μL of sample volume were injected into a flow of 250 μL/min. Analytes eluted in the order of DON-3-GlcA (4.9 min), D3G (5.7 min), DON (5.9 min) and DOM-1 (6.3 min). Mass spectrometric detection was performed using negative ESI and selective reaction monitoring mode as described by Nagl et al. (2012).
2.5. Method validation and data evaluation
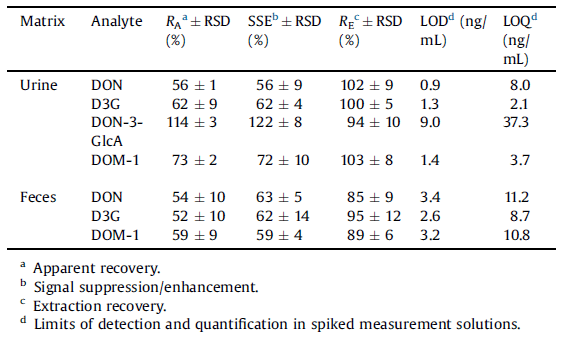
The determined method performance parameters include apparent recovery (RA), signal suppression/enhancement (SSE), recovery of the extraction step (RE), repeatability (RSD), as well as limits of detection (LODs) and quantification (LODs). Prior and after extraction, blank urine and feces samples (collected from piglets after negative control treatment) were spiked with standard compounds in triplicates at 5 and 4 spiking levels, respectively. The method for determination of DON, D3G, DON-3-GlcA and DOM-1 in urine was validated in a working range of 3–100 ng/mL in the measurement solution. For feces, method performance character- istics were determined for DON, DOM-1 and D3G in a working range between 30 and 300 ng/mL. Data evaluation was performed as stated in Nagl et al. (2012). Since oral DON and D3G administrations were incomplete in one of the piglets, data from the respective animal were not included in the calculations.
Table 1 Method performance parameters for the pig urine (n = 15) and feces (n = 12).
3. Results and discussion
3.1. Sample preparation and method performance
Interfering matrix components can represent a major challenge in analysis of DON and its metabolites in biological samples, especially in urine (Gambacorta et al., 2013; Warth et al., 2011). During method development, different sample clean-up techniques, including solid phase extraction, salting-out assisted liquid– liquid extraction and dilute-and-shoot (DAS), were tested. Since DAS comprised satisfactory apparent recoveries of analytes (56–114%) and simple sample preparation, this approach was chosen for final analysis of urine samples. Due to the dilution of samples by a factor of 10, overall limits of detection (LODs, S/N = 3) for DON, D3G, DON-3-GlcA and DOM-1 were 9, 13, 90 and 14 ng/ mL, respectively. Corresponding limits of quantification (LOQs, S/N = 10) of 80, 21, 373 and 37 ng/mL were obtained. In standard solutions, LODs and LOQs ranged between 0.5–3.3 and 1.5–9.4 ng/mL. Detailed results of method validation are provided in Table 1.
Fig. 2. SRM chromatogram (— m/z 471 -> 113, - - - m/z 471 -> 265) of DON-3-GlcA and DON-15-GlcA in a urine sample of a piglet orally dosed with DON. In enhanced product ion scans (inlays, collision energy = -40 eV) the fragmentation product of m/z 471 -> 441 is absent in the second DON-GlcA-isomer, suggesting the presence of DON-15-GlcA.
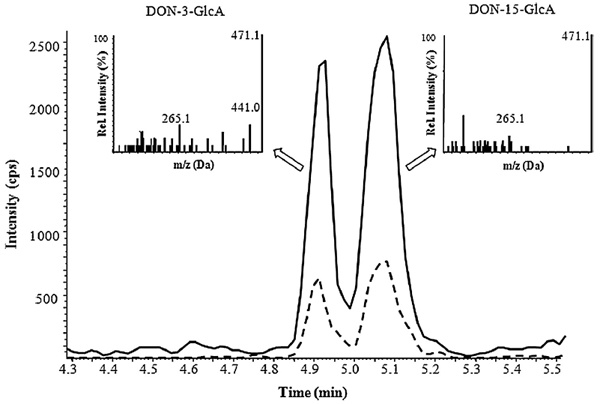
Besides DON-3-GlcA, a second DON-GlcA peak was detected in contaminated urine samples (Fig. 2). Further investigations of this peak by MS/MS experiments and enzymatic treatment according to Warth et al. (2012) suggested the presence of DON-15-GlcA. Most prominently, a fragment of m/z 441 (resulting from the loss of CH2O from C15) is observed in DON-3-GlcA, but not in DON-15- GlcA (see inlays of Fig. 2). Due to the lack of a suitable standard, quantification of DON-15-GlcA was performed using DON-3-GlcA calibrants. The comparatively higher MS response of DON-15-GlcA reported by Warth et al. (2012) was verified with our instrumental setup and taken into consideration for all further calculations (correction factor 1.88).
For feces, sample preparation consisted of a three step extraction, followed by protein precipitation. Reduced apparent recoveries (54–59%) were caused by matrix suppression rather than by insufficient extraction of analytes (Table 1). Dilutions of the samples by a factor of 56 resulted in LODs of 188, 146, 179 ng/g for DON, D3G and DOM-1, respectively, while corresponding LOQs of 626, 486 and 605 ng/g were determined.
To sum up, matrix interferences could be observed for both matrices and all analytes. Although their extent was reduced to a level acceptable for analysis of our samples, the importance of a validation for each analyte–matrix combination was once more highlighted.
3.2. Clinical symptoms
DON can cause severe adverse health effects in pigs, e.g., anorexia, emesis or weight loss (reviewed by Pestka, 2007). Due to animal welfare considerations as well as possible influences on the toxins’ oral bioavailability (in case of emesis), amounts of administered toxins were chosen on the basis of no observed adverse effect levels for oral (Forsyth et al., 1977), and intravenous DON application (Prelusky, 1993; Prelusky and Trenholm, 1993) in pigs, respectively. Independent of the treatment, neither retching nor emesis was observed in any of the animals. Feed intake was noticed for all piglets within 30 min after toxin administrations.
3.3. Detected amounts of excreted toxins in urine and feces
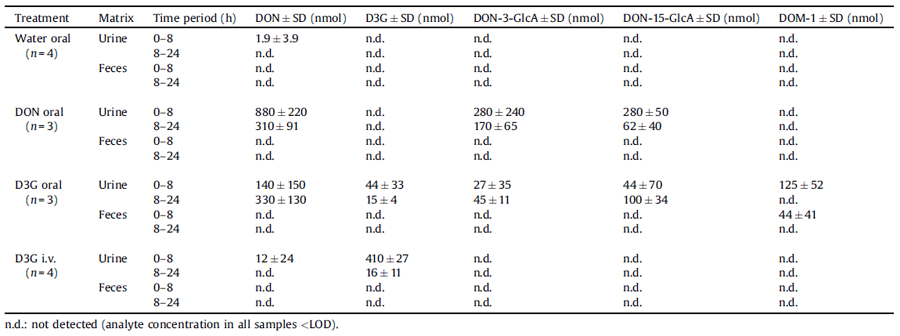
In the individual sampling periods, volumes of collected urine ranged between 22 and 838 mL per piglet, while amounts of freeze-dried feces varied between 2 and 22 g. In urine, concentrations of DON, D3G, DON-3-GlcA, DON-15-GlcA and DOM-1 were between 105 and 1420 ng/mL, 50.6 and 1100 ng/mL, 542 and 1030 ng/mL, 198 and 705 ng/mL and 84.8 and 501 ng/mL, respectively. Of all the analytes tested only DOM-1 was found in feces, with concentrations ranging between 914 and 1220 ng/g. To facilitate comparability between the excreted amounts of different toxins, results are expressed as molar amounts henceforth.
3.3.1. Negative control (water administration)
After oral application of water, traces of DON could be determined in urine samples of the first sampling period (Table 2). The provided feed was analyzed for DON, D3G, 3-ADON and 15- ADON before the start of the experiment and additional feed samples were taken on each of the treatment days. LC–MS/MS measurements revealed low concentrations of DON (67 ± 28 μg/ kg) in the diet, whereas none of the other analytes could be quantified (LOQs 20 μg/kg). Thus, the minimal DON contamination of the diet already led to detectable DON concentrations in urine. However, the daily amount of DON ingested via feed reflected only ≤2% of the toxin doses administered in the following treatments and was therefore not considered in further calculations.
3.3.2. Oral DON administration
Following oral administration of DON, we found DON, DON-3- GlcA and DON-15-GlcA in urine of piglets. The majority of analytes was excreted in the first sampling period (Table 2), thus confirming a fast elimination of the mycotoxin (Pestka, 2007). DON was the major urinary metabolite, representing 62.1 ±6.2% of the total analytes excreted in urine. The percentage of glucuronidated DON in pig urine (37.9 ± 6.2%) corresponds to values obtained in previous studies (Dänicke et al., 2005; Sundstøl Eriksen et al., 2003), in which DON-GlcA was deter- mined via indirect methods (enzymatic hydrolysis). Yet higher glucuronidation rates were observed in other species (Meky et al., 2003; Warth et al., 2012). A comparably low activity of porcine UDP-glucuronosyltransferases, as shown by Maul et al. (2012), may account for this phenomenon.
Table 2 Total amounts of recovered DON, D3G, DON-3-GlcA, DON-15-GlcA and DOM-1 in urine and feces of dosed piglets, respectively. Mean values ± standard deviations (SD) are listed for the indicated time periods after oral administration of water, DON (0.25 mmol/kg b.w.) and D3G (0.25 mmol/kg b.w.) as well as after intravenous D3G application (0.03 mmol/kg b.w.). Since oral toxin administrations were incomplete in one of the animals, data from the respective piglet were excluded.
In recentyears,special focushas beenputnotonlyon the extentof glucuronidation, but also on species-dependent variations regarding the formation of different DON-GlcA isomers. Lattanzio et al. (2011) were the first to postulate the presence of two DON-GlcA conjugates in rat urine, while Warth et al. (2012) successfully identified DON-3- GlcA and DON-15-GlcA in urine of humans. In addition, evidence for the formationoffurtherDON-GlcAshasbeen given(Mauletal., 2012; Nagl et al., 2012; Uhlig et al., 2013; Warth et al., 2013). To the best of our knowledge, the present study provides first in vivo data on the occurrence of different DON-GlcA isomers in pigs. Besides DON-3- GlcA, a second conjugate was detected in the urine of DON exposed piglets and was tentatively identified as DON-15-GlcA. While the latter was found tobe the predominantconjugate in humans (Warth et al., 2012, 2013), ratios of DON-15-GlcA to DON-3-GlcA varied between individual piglets (0.5–2.2). Further studies are needed to investigate this aspect in more detail.
The high susceptibility of pigs to DON is often related to the species’ poor ability to degrade DON to DOM-1 (Pestka 2007). In our experiment, DOM-1 could not be detected in urine or feces samples following DON indigestion. Moreover, none of the analytes included in our LC–MS/MS method seemed to be present in feces of piglets dosed with DON. While a complete lack of DOM- 1 formation in swine is reported in literature (Sundstøl Eriksen et al., 2002), the absence of any toxins in feces may be due to several reasons. First, dilution of feces samples resulted in higher LODs and LOQs than in urine, thus raising the possibility that minimal toxin amounts were not detectable. More importantly, DON is primarily eliminated via urine in pigs. Since excreted toxin amounts in feces were reported to account for only 0.1–3% of the administered dose (Goyarts and Dänicke, 2006; Dänicke and Brezina, 2013), our results are not in conflict with previous studies.
3.3.3. Oral D3G administration
In urine of piglets orally dosed with D3G, the masked mycotoxin itself, as well as DON, DON-3-GlcA, DON-15-GlcA and DOM-1 could be found. Amounts of urinary excreted D3G were low, especially in the second time period (15 ± 4 nmol). A limited bioavailability of D3G has already been shown for rats (Nagl et al., 2012). In a human volunteer consuming a diet naturally contaminated with D3G (7 μg D3G/day), the masked mycotoxin could not be detected in urine at all (Warth et al., 2013). A likely explanation for these findings is provided by De Nijs et al. (2012) who showed that D3G is far less absorbed by human intestinal Caco-2 cells compared to DON. Taking into consideration both the limited bioavailability of D3G and its lower biological activity on the ribosomal level (Poppenberger et al., 2003) data indicate so far that D3G is less toxic than its parent toxin in mammals.
Yet major concerns have been raised regarding a possible hydrolysis of D3G with subsequent release of its aglycon in the digestive system. Results of several in vitro studies have supported these suggestions (Berthiller et al., 2011; Dall´Erta et al., 2013; Gratz et al., 2013), while in vivo reports on the issue are rare and only available for rats (Nagl et al., 2012; Veršilovskis et al., 2012). In the present experiment, DON as well as DON-15-GlcA and DON-3-GlcA were found in urine of piglets after oral D3G administration, confirming that D3G is cleaved to DON, absorbed and metabolized to DON-GlcA in pigs. Since these findings correlate well with those observed in rats (Nagl et al., 2012), hydrolysis of ingested D3G seems not to be species-dependent, but potentially a common feature of mammals in general. Notably, urinary excreted amounts of DON and DON-GlcA increased in samples of the second sampling period, with DOM-1 solely being detectable 8–24 h after dosing (Table 1). Consistently, DOM-1 was also found in feces samples of this time period. In pigs, this metabolite is known to be mainly formed in the hindgut (Dänicke et al., 2004a). With respect to the seemingly low absorption of D3G in the small intestine (De Nijs et al., 2012), it is likely that relevant amounts of the masked mycotoxin reach distal parts of the gut, where they are hydrolyzed to DON, further metabolized to DOM-1 and absorbed. This scenario would explain the differences between the oral DON and D3G treatments regarding both the presence of DOM-1 in investigated matrices as well as the delayed urinary excretion of D3G metabolites. However, further specific study designs are necessary to verify this hypothesis.
3.3.4. Intravenous D3G administration
The vast majority of intravenously administered D3G was excreted in unmetabolized form via urine in the first sampling period (Table 2). In contrast, only traces of urinary DON could be detected, whereas none of the other analytes was found. Berthiller et al. (2011) have shown that human cytosolic b-glucosidase, an enzyme expressed in liver, kidney, spleen and gut (Berrin et al., 2002), had no activity with D3G. In agreement, our data strongly indicate that hydrolysis of D3G is marginal after systemic absorption (or even not occurring at all if taking into account that low levels of urinary DON were also detected after water administration). As a consequence, we suppose that in vivo cleavage of D3G predominantly occurs in the digestive tract.
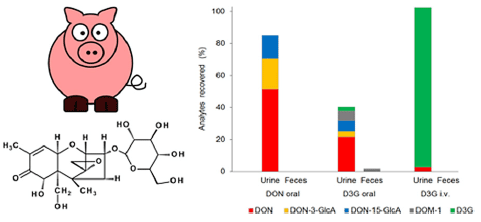
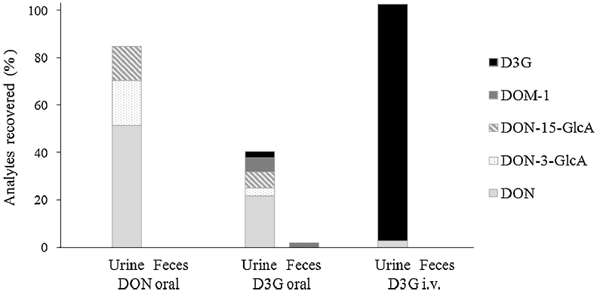
Fig. 3. Excretion of DON, D3G and their metabolites in urine and feces of toxin exposed piglets. Values are expressed as DON equivalent percentages of the administered dose. V. Nagl et al. / Toxicology Letters 229 (2014) 190–197 195
3.4. Total recoveries
The quantitative urinary recovery of DON allows an appropriate estimation of the toxin’s bioavailability in pigs (Goyarts and Dänicke, 2006). After oral administration of DON, 84.8 ± 9.7% of the administered dose were detected in urine within 24 h in total (Fig. 3). Unmetabolized DON accounted for 51.4 ± 6.3%, while DON- 3-GlcA and DON-15-GlcA reflected 19.0 ± 6.8% and 14.5 ±1.8% of the applied toxin dose, respectively. The high oral bioavailability of DON is in line with literature reports, where urinary DON recoveries between 44 and 82% were assessed after single oral/ intragastric DON administration (Dänicke et al., 2004b; Prelusky et al., 1988). Furthermore, urinary recoveries determined in our study are comparable to those observed in humans (summarized by Turner et al., 2012) underlining once more the similarities between those species in regard to DON metabolism.
Following oral D3G application, the overall amount of recovered analytes reflected 42.1 ±8.5% of the administered toxin dose. Thereof, urinary excreted metabolites represented 40.3 ± 10.0%, whereas elimination via feces accounted for only 1.8 ± 1.6%. Although the primary excretion route of D3G seems to differ between pigs and rats, the low oral bioavailability of the masked mycotoxin applies for both species: urinary D3G accounted for 2.6 ± 1.4% in our study, while amounts of 0.3 ± 0.1% were reported for rats (Nagl et al., 2012). DON, DON- 3-GlcA, DON-15-GlcA and DOM-1 in piglets’ urine reflected 21.6 ± 3.7%, 3.4 ± 2.0%, 6.8 ± 4.1% and 5.9 ± 2.9% of the administered dose, respectively. Despite the nearly complete hydrolysis and metabolization of D3G, the absorption of the formed metabolites seems to be limited. In comparison to DON treatment, the proportion of urinary excreted metabolites was reduced by a factor of 2, approximately. DON is predominantly absorbed in proximal parts of the small intestine in pigs (Dänicke et al., 2004a), whereas in vitro experiments indicate that D3G is hydrolyzed thereafter (Berthiller et al., 2011; Dall´Erta et al., 2013). Thus, the obtained in vivo data are not entirely surprising. Still, also nonabsorbed hydrolysis products of D3G could exhibit adverse health effects. For example, DON was shown to induce intestinal damage by alteration of tight junction protein expression and decrease of the barrier function (Pinton et al., 2009). Data dealing with possible effects of D3G and its degradation products on the gut health are lacking so far.
Yet another question of interest is the discrepancy in the overall recovered toxin amounts between oral DON and D3G treatment. One explanation might be an incomplete elimination of D3G by 24 h, as indicated by the considerable amounts of urinary excreted analytes 8–24 h after dosing. Thus, the short sampling period represents a limitation of our study. Moreover, we cannot ensure that the employed biomarker method includes all metabolites and degradation products which are actually formed of D3G in vivo. For example, novel sulfated DON metabolites have been identified in rats and chicken just recently (Wan et al., 2014). Furthermore, some intestinal bacteria may be able to use the phosphorelay uptake system and convert D3G into DON-3-phosphoglucoside, but are inefficient to cleave this metabolite into DON and glucosephosphate. Currently, no standard is available for DON-3- phosphoglucoside. Also the formation of other (fecal) metabolites cannot be excluded. However, to encounter this problem to some extent, we investigated the presence of DOM-1-glucuronide (DOM-1-GlcA) in urine samples. Enzymatic treatment resulted in an increase of DOM-1 concentrations in urine of piglets orally dosed with D3G by a factor of 3.5 on average, thus indicating the presence of DOM-1-glucuronide. Due to a lack of a suitable standard, this metabolite was not quantified in our study. In comparison, an extensive metabolization of the masked mycotoxin after its systemic absorption seems rather unlikely, since almost the complete amount of intravenously administered D3G was recovered in unmetabolized form in urine (99.9 ± 8.9% out of 102.4 ± 5.4% total recovery). In the future, the investigation and identification of unaccounted D3G metabolites, especially of those formed by gut microbiota, using high resolution mass spectrom- eters will be a major task.
In conclusion, we demonstrated that ingested D3G is nearly completely hydrolyzed in pigs, but only partially absorbed. Results indicate that the cleavage predominantly occurs in the digestive tract, while the toxin seems to be rather stable after systemic absorption. Compared to DON, the proportion of urinary excreted metabolites after oral D3G administration was reduced by a factor of 2, approximately. Based on these data we presume that D3G is less bioavailable than its parent toxin in pigs and therefore of lower toxicological relevance. However, the bioavailability of D3G in pigs may increase after chronic exposure via feed, as it has already been shown for DON (Goyarts and Dänicke, 2006). Clarification of this aspect will be also important regarding the risk assessment of D3G and its potential implementation in regulatory limits. Further- more, it should be emphasized that D3G may exhibit biological activity on its own. Therefore, future studies should for instance address possible emetic effects of the masked mycotoxin or its influence on intestinal gut health.
Conflict of interest
The authors declare that there are no conflicts of interest.
Transparency document
The Transparency document associated with this article can be found in the online version.
Acknowledgements
The authors thank the Austrian Federal Ministry of Economy, Family and Youth, the National Foundation for Research, Technology and Development, BIOMIN Holding GmbH and Nestec Ltd. for funding the Christian Doppler Laboratory for Mycotoxin Metabolism. The financial support of the Lower Austrian Govern- ment (funding of used mass spectrometers) and the Vienna Science and Technology Fund (WWTF LS12-021; production of D3G) is greatly acknowledged. Furthermore, we owe sincere gratitude to Anja Joachim, Barbara Hinney and Lukas Schwarz from the University of Veterinary Medicine Vienna for provision of the animal housing facilities. In addition, we would like to thank Philipp Fruhmann, Hannes Mikula, Christian Hametner and Johannes Fröhlich from the Vienna University of Technology for providing the DON-3-GlcA reference standard. We also appreciate the valuable support of Thomas Mandl, Friedrich Reis and Klaus Teichmann in course of preparation of the experimental diet. Finally, we want to address special thanks to Alexandra Malachova for analyses of feed samples and Silvia Caha for her help during sample preparation.
References
Arunachalam, C., Doohan, F.M., 2013. Trichothecene toxicity in eukaryotes: cellular and molecular mechanisms in plants and animals. Toxicol. Lett. 217, 149–158.
Berrin, J.G., McLauchlan, W.R., Needs, P., Williamson, G., Puigserver, A., Kroon, P.A., Juge, N., 2002. Functional expression of human liver cytosolic beta-glucosidase in Pichia pastoris. Insights into its role in the metabolism of dietary glucosides. Eur. J. Biochem. 269, 249–258.
Berthiller, F., Dall'Asta, C., Schuhmacher, R., Lemmens, M., Adam, G., Krska, R., 2005. Masked mycotoxins: determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography–tandem mass spectrometry. J. Agric. Food Chem. 53, 3421–3425.
Berthiller, F., Krska, R., Domig, K.J., Kneifel, W., Juge, N., Schuhmacher, R., Adam, G., 2011. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 206, 264–267.
Berthiller, F., Crews, C., Dall'Asta, C., De Saeger, S., Haesaert, G., Karlovsky, P., Oswald, I.P., Seefelder, W., Speijers, G., Stroka, J., 2013. Masked mycotoxins: a review. Mol. Nutr. Food Res. 57, 165–186.
CAST (Council for Agricultural Science and Technology Task Force), 2003. Mycotoxins: Risks in Plant, Animal, and Human Systems. Council for Agricultural Science and Technology Task Force, Iowa, USA Report No. 139.
Dall´Erta, A., Cirlini, M., Dall'Asta, M., Del Rio, D., Galaverna, G., Dall’Asta, C., 2013. Masked mycotoxins are efficiently hydrolysed by the human colonic microbiota, releasing their toxic aglycones. Chem. Res. Toxicol. 26, 305–312.
Dänicke, S., Valenta, H., Döll, S., 2004a. On the toxicokinetics and the metabolism of deoxynivalenol (DON) in the pig. Arch. Anim. Nutr. 58, 169–180.
Dänicke, S., Valenta, H., Goyarts, T., Razzazi, E., Böhm, J., 2004b. On the effects of increasing deoxynivalenol (DON) concentrations in pig feed on growth performance and utilization of nutrients and on DON metabolism. J. Anim. Feed Sci. 13, 539–556.
Dänicke, S., Brüssow, K.-P., Valenta, H., Ueberschär, K.-H., Tiemann, U., Schollenberger, M., 2005. On the effects of graded levels of Fusarium toxin contaminated wheat in diets for gilts on feed intake, growth performance and metabolism of deoxynivalenol and zearalenone. Mol. Nutr. Food Res. 49, 932–943.
Dänicke, S., Brezina, U., 2013. Kinetics and metabolism of the Fusarium toxin deoxynivalenol in farm animals: consequences for diagnosis of exposure and intoxication and carry over. Food Chem. Toxicol. 60, 58–75.
De Boevre, M., Di Mavungu, J.D., Landschoot, S., Audenaert, K., Eeckhout, M., Maene, P., Haesaert, G., De Saeger, S., 2012. Natural occurrence of mycotoxins and their masked forms in food and feed products. World Mycotox J. 5, 207–219.
De Nijs, M., Van den Top, H.J., Portier, L., Oegema, G., Kramer, E., Van Egmond, H.P., Hoogenboom, L.A.P., 2012. Digestibility and absorption of deoxynivalenol-3- ß-glucoside in in vitro models. World Mycotox J. 5, 319–324.
European Commission, 2006. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=CON- SLEG:2006R1881:20120401:EN:PDF (last retrieved 10th February 2014).
Forsyth, D.M., Yoshizawa, T., Morooka, N., Tuite, J., 1977. Emetic and refusal activity of deoxynivalenol to swine. Appl. Environ. Microbiol. 34, 547–552.
Fruhmann, P., Warth, B., Hametner, C., Berthiller, F., Horkel, E., Adam, G., Sulyok, M., Krska, R., Fröhlich, J., 2012. Synthesis of deoxynivalenol-3-ß-D-O-glucuronide for its use as biomarker for dietary deoxynivalenol exposure. World Mycotox J. 5, 127–132.
Gambacorta, L., Solfrizzo, M., Visconti, A., Powers, S., Cossalter, A., Pinton, P., Oswald, I., 2013. Validation study on urinary biomarkers of exposure for aflatoxin B 1, ochratoxin A, fumonisin B 1, deoxynivalenol and zearalenone in piglets. World Mycotox J. 6, 299–306.
Goyarts, T., Dänicke, S., 2006. Bioavailability of the Fusarium toxin deoxynivalenol (DON) from naturally contaminated wheat for the pig. Toxicol. Lett. 163, 171–182.
Gratz, S.W., Duncan, G., Richardson, A.J., 2013. Human fecal microbiota metabolize deoxynivalenol and deoxynivalenol-3-glucoside and may be responsible for urinary de-epoxy deoxynivalenol. Appl. Environ. Microbiol. 79, 1821–1825.
JECFA, 2011. Evaluation of certain contaminants in food: Seventy-second report of the Joint FAO/WHO Expert Committee on Food Additives. WHO Technical Report Series 959. http://whqlibdoc.who.int/trs/WHO_TRS_959_eng.pdf (last retrieved 10th February 2014).
Lancova, K., Hajslova, J., Poustka, J., Krplova, A., Zachariasova, M., Dostálek, P., Sachambula, L., 2008. Transfer of Fusarium mycotoxins and ‘masked’ deoxynivalenol (deoxynivalenol-3-glucoside) from field barley through malt to beer. Food Addit. Contam. A 25, 732–744.
Lattanzio, V.M.T., Solfrizzo, M., De Girolamo, A., Chulze, S.N., Torres, A.M., Visconti, A., 2011. LC–MS/MS characterization of the urinary excretion profile of the mycotoxin deoxynivalenol in human and rat. Food Addit. Contam. B 879, 707–715.
Lemmens, M., Scholz, U., Berthiller, F., Dall'Asta, C., Koutnik, A., Schuhmacher, R., Adam, G., Buerstmayr, H., Mesterházy Á, Krska, R., 2005. The ability to detoxify the mycotoxin deoxynivalenol colocalizes with a major quantitative trait locus for Fusarium head blight resistance in wheat. Mol. Plant Microbe Interact. 18, 1318–1324.
Nagl, V., Schwartz, H., Krska, R., Moll, W.D., Knasmüller, S., Ritzmann, M., Adam, G., Berthiller, F., 2012. Metabolism of the masked mycotoxin deoxynivalenol-3- glucoside in rats. Toxicol. Lett. 18, 367–373.
Malachova, A., Dzuman, Z., Veprikova, Z., Vaclavikova, M., Zachariasova, M., Hajslova, J., 2011. Deoxynivalenol, deoxynivalenol-3-glucoside, and enniatins: the major mycotoxins found in cereal-based products on the Czech market. J. Agric. Food Chem. 59, 12990–12997.
Maresca, M., 2013. From the gut to the brain: journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 5, 784–820.
Maul, R., Müller, C., Rieß, S., Koch, M., Methner, F.J., Nehls, I., 2012. Germination induces the glucosylation of the Fusarium mycotoxin deoxynivalenol in various grains. Food Chem. 131, 274–279.
Meky, F.A., Turner, P.C., Ashcroft, A.E., Miller, J.D., Qiao, Y.L., Roth, M.J., Wild, C.P., 2003. Development of a urinary biomarker of human exposure to deoxy- nivalenol. Food Chem. Toxicol. 41, 265–273.
Pestka, J.J., 2007. Deoxynivalenol: toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 137, 283–298.
Pestka, J.J., 2010. Deoxynivalenol: mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 84, 663–679.
Pinton, P., Nougayrède, J., Del Rio, J., Moreno, C., Marin, D.E., Ferrier, L., Bracarense, A., Kolf-Clauw, M., Oswald, I.P., 2009. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol. 237, 41–48.
Poppenberger, B., Berthiller, F., Lucyshyn, D., Sieberer, T., Schuhmacher, R., Krska, R., Kuchler, K., Glössl, J., Luschnig, C., Adam, G., 2003. Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 278, 47905–47914.
Prelusky, D.B., 1993. The effect of low level deoxynivalenol on neurotransmitter levels measured in pig cerebral spinal fluid. J. Environ. Sci. Health B 28, 731–761.
Prelusky, D.B., Trenholm, H.L., 1993. The efficacy of various classes of anti-emetics in preventing deoxynivalenol-induced vomiting in swine. Nat. Toxins 1, 296–302.
Prelusky, D., Hartin, K., Trenholm, H., Miller, J., 1988. Pharmacokinetic fate of 14C- labeled deoxynivalenol in swine. Fundam. Appl. Toxicol. 10, 276–286.
Sundstøl Eriksen, G., Pettersson, H., Johnsen, K., Lindberg, J.E., 2002. Transformation of trichothecenes in ileal digesta and faeces from pigs. Arch. Anim. Nutr. 56, 263–274.
Sundstøl Eriksen, G., Pettersson, H., Lindberg, J.E., 2003. Absorption, metabolism and excretion of 3-acetyl don in pigs. Arch. Anim. Nutr. 57, 335–345.
Sundstøl Eriksen, G., Pettersson, H., Lundh, T., 2004. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 42, 619–624.
Turner, P.C., Flannery, B., Isitt, C., Ali, M., Pestka, J., 2012. The role of biomarkers in evaluating human health concerns from fungal contaminants in food. Nutr. Res. Rev. 25, 162–179.
Uhlig, S., Ivanova, L., Faeste, C.K., 2013. Enzyme-assisted synthesis and structural characterization of the 3-, 8- and 15-glucuronides of deoxynivalenol. J. Agric. Food Chem. 61, 2006–2012.
Varga, E., Malachova, A., Schwartz, H., Krska, R., Berthiller, F., 2013. Survey of deoxynivalenol and its conjugates deoxynivalenol-3-glucoside and 3-acetyldeoxynivalenol in 374 beer samples. Food Addit. Contam. A 30, 137–146.
Veršilovskis, A., Geys, J., Huybrechts, B., Goossens, E., De Saeger, S., Callebaut, A., 2012. Simultaneous determination of masked forms of deoxynivalenol and zearalenone after oral dosing in rats by LC–MS/MS. World Mycotox J. 5, 303– 318.
Wan, D., Huang, L., Pan, Y., Wu, Q., Chen, D., Tao, Y., Wang, X., Liu, Z., Li, J., Wang, L., 2014. Metabolism, distribution and excretion of deoxynivalenol with combined techniques of radio-tracing, HPLC-IT-TOF/MS and online radiometric detection. J. Agric. Food Chem. 62, 288–296.
Warth, B., Sulyok, M., Berthiller, F., Schuhmacher, R., Fruhmann, P., Hametner, C., Adam, G., Fröhlich, J., Krska, R., 2011. Direct quantification of deoxynivalenol glucuronide in human urine as biomarker of exposure to the Fusarium mycotoxin deoxynivalenol. Anal. Bioanal. Chem. 401, 195–200.
Warth, B., Sulyok, M., Fruhmann, P., Berthiller, F., Schuhmacher, R., Hametner, C., Adam, G., Fröhlich, J., Krska, R., 2012. Assessment of human deoxynivalenol exposure using an LC–MS/MS based biomarker method. Toxicol. Lett. 211, 85– 90.
Warth, B., Sulyok, M., Berthiller, F., Schuhmacher, R., Krska, R., 2013. New insights into the human metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone. Toxicol. Lett. 220, 88–94.
Wu, X., Murphy, P., Cunnick, J., Hendrich, S., 2007. Synthesis and characterization of deoxynivalenol glucuronide: its comparative immunotoxicity with deoxyni- valenol. Food Chem. Toxicol. 45, 1846–1855.
Related topics
Authors:

Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post




You may be interested in
