Introduction
Mycotoxins are secondary metabolites of low molecular weights produced by certain strains of filamentous fungi such as Aspergillus, Fusarium and Penicillium, which invade crops in the field and may grow on foods during storage under favorable conditions of temperature and humidity (Shamsudeen et al., 2013). The most common mycotoxins are aflatoxins, fumonisins, ochratoxin A, trichothecenes, zearalenone, and out of which aflatoxins (AF) commonly contaminate a wide variety of tropical and subtropical food/feedstuffs (Katole et al., 2013).
Organic (chelated) trace minerals are different from its inorganic counterpart as copper (Cu), ferrous (Fe), selenium (Se) and zinc (Zn) are held with metal binding agents (ligand) by high energy bond known as co-ordinate covalent bond (Leeson and Summers, 2002). Several studies report that the chelated minerals are more bioavailable to animals and poultry (Wedekind et al., 1992) than its inorganic counterpart and hence one can expect more counteracting effect of this form of minerals against aflatoxin (AF) than inorganic form. Dietary supplementation of any nutrient especially trace minerals that counteract aflatoxicosis cannot be recommended based on in vivo experiments alone, because the nutrients which counteract AF in vivo may influence the fungal growth and concomitant AF production in vitro in feed ingredient/ mixed feed (Shamsudeen et al., 2013). For example, the supplementation of Se was found to be effective against aflatoxicosis in poults (Burguera et al., 1983), but the effect of supplemental Se on AF production by the fungi in feed is not known. When dietary supplementation of any nutrient against aflatoxicosis increases its utilization by the contaminated fungi for AF production in feed, then its use is limited, even though the nutrient is effective against aflatoxicosis. Trace elements as inorganic form are an important factor that influences the fungal growth and AF production in feed ingredient (Jones et al., 1984) and in liquid media (Marsh et al., 1975). Trace metal elements such as Cu, Fe and Zn have a regulatory effect at the cellular and molecular level on fungal growth and concomitant mycotoxin synthesis. A nutrient which is effective against aflatoxicosis and at the same time, which is either unavailable to contaminated fungi of feed for AF production or toxic to fungi, is only useful for supplementation against aflatoxicosis. Even though several reports are available on the effect of inorganic trace metals on fungal growth and AF production in liquid media and in feed ingredients, literature regarding the effect of supplemental chelated minerals on fungal growth and AF production in liquid media and feed ingredient are scanty. The objective of present investigation is to study the effect of certain chelated and inorganic trace minerals on fungal growth and AF production in liquid media.
Materials and Methods
Preparation of basal medium
A synthetic medium as suggested by Reddy et al. (1971) for AF production was used without addition of trace minerals, as basal medium. The composition of basal medium is given in Table 1. All the required ingredients of basal medium were taken and pH of the medium was adjusted to 4.5. Ninety ml of the media were then transferred to individual flasks of similar type with same capacity and surface area. All the flasks were then autoclaved at 15 1bs pressure for 15 minutes and cooled.
Preparation of experimental media
The trace minerals were added to sterile distilled water to prepare trace mineral solution and it was prepared in such a way that one ml of solution contains calculated amount of minerals. Trace minerals which were not soluble in water, were weighed individually and added to basal medium. The trace mineral solutions were then added to basal media for preparation of different experimental media. Briefly, 10 ml of basal media were taken in test tubes, sterilized by autoclaving at 15 1b pressure for 15 minutes and cooled. Then, 1 ml of trace mineral solution was added to basal media under sterile condition and this mixed solution was again sterilized by cold sterilization by keeping the test tubes in maximum slanting position directly under UV light of laminar flow for 3 hours. Since we are not sure about the effect of autoclaving on metal-ligand bond of the chelated minerals, these minerals were not subjected to autoclaving. Ten ml of basal media with one ml of trace mineral solution in test tubes were then transferred to 90 ml basal media under sterile condition in laminar flow. For control flasks, 10 ml of basal media with one ml of sterile distilled water were added. Final quantity of basal media was 100 ml in all the flasks. All the flasks after addition of trace minerals were individually marked and incubated at room temperature for 2 days to check any bacterial/ fungal contamination.
Experimental design
In experiment I, Cu was supplemented and in experiment II Fe was supplemented to basal media. In experiment III and IV, Se and Zn were supplemented respectively. The details of supplementation in these experiments are given in Table 2. Each treatment was replicated by six observations and experimental design followed was Completely Randomized Design (CRD).
Inoculation of mould and incubation of media
The experimental media were inoculated with one ml of fungal spore suspension containing 106 -107 spores/ml of PBS under sterile condition. The flasks were then horizontally placed in the BOD incubator pre-maintained at 26° C±1°C to provide larger surface area for the mycelial growth and were incubated for 10 days.
Analysis of response criteria
After 10 days of incubation, the flasks were taken out from incubator and further processed for analysis of fungal biomass, AF production and pH change of media.
Fungal biomass and aflatoxin production
The fungal biomass as influenced by various treatments was estimated by the method as suggested by Cuero and Oullet (2005). The AF production in liquid media as influenced by various treatments was determined by Thin Layer Chromatography (TLC) as suggested by Tsai et al. (1984).
Results and Discussion
Effect of copper on fungal growth and aflatoxin production
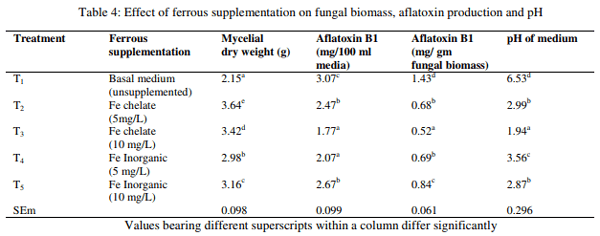
The fungal growth measured as mean mycelial dry weight, AFB1 production and change in pH of the media as influenced by chelated and inorganic Cu are presented in Table 3. The inorganic Cu at both levels and chelated Cu at lower level had higher fungal biomass, followed by higher level of chelated Cu than control (T1). Similarly, a significant effect (p<0.01) on AFB1 production was also noted. The basal media supplemented with 5 mg and 10 mg of inorganic Cu/L had higher total AFB1 production followed by basal media supplemented with 5 mg and 10 mg of chelated Cu/L than control. However, the media added with 5mg Cu/L in chelated and inorganic form did not differ significantly among each other. The result exerted non significant difference between treatment groups on AFB1 production per gm of fungal biomass. However, the pH of media supplemented with inorganic Cu was significantly (p<0.01) lower than media supplemented with 5 mg Cu/L or control without supplementation. The result of the experiment revealed that the chelated and inorganic form of Cu increased fungal growth. The AFB1 production in 100 ml media was also increased. However the production is higher in inorganic form than chelated form, but, the AFB1 per gm of fungal biomass was not increased. The results are in agreement with earlier findings of Cuero et al. (2003), Aziz and Moussa (1997), who also observed increased fungal growth and AF production due to Cu supplementation in liquid media. Increased AF production without increase in fungal growth was observed by Cuero and Ouellet (2005). On the contrary, Marsh et al. (1975) observed decreased AF production without influencing fungal biomass due to Cu at an added level of ≤ 25 mg/L. The lower AF production in 10 mg/L Cu chelate group than Cu inorganic groups might possibly be due to unavailability of the mineral ion as a result of chelation for AF production and higher in AF production in Cu chelate groups than control might possibly due to the trace of inorganic Cu in the Cu chelate product, since a traces of mineral is sufficient for AF production in liquid media.
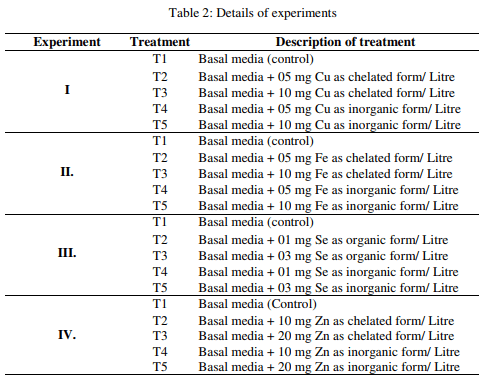
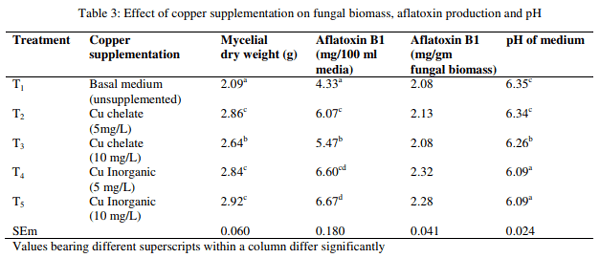
Effect of ferrous on fungal growth and aflatoxin production
The fungal growth measured as mean mycelial dry weight, AFB1 production and change in pH of the media as influenced by chelated and inorganic Fe are presented in Table 4. The result revealed that the mycelial dry weight was higher (p<0.01) in media with Fe (5 mg/L) in chelate form than basal media without Fe supplementation and the mycelial dry weight of other groups were found to be intermediary. The AFB1 production in 100 ml of media was lower (p<0.01) in media with Fe chelate at 10 mg/L or Fe inorganic at 5 mg/L than media without Fe supplementation, whereas the AFB1 production of other two groups was found to be intermediary. The AFB1 per gm fungal biomass and pH of media were significantly (p<0.01) lower in media with Fe chelate at 10 mg/L than media without Fe supplementation, while the AFB1 per gm fungal biomass and pH of media of other groups were found to be intermediary.
The result revealed that chelated and inorganic form of Fe increased fungal growth than control and it was higher in Fe chelate groups than Fe inorganic group. This is in accordance with earlier finding of Aziz and Moussa (1997) who also observed increased fungal growth due to Fe. But in contrary, Cuero et al. (2003) did not observe any change in fungal growth, while Cuero and Ouellet (2005) observed decreased fungal growth due to Fe supplementation. The AFB1 production in 100 ml media and AFB1 per gm of fungal biomass was decreased by both form of Fe supplementation and no difference could be observed between chelated and inorganic form of Fe. Similar observations were made by Tiwari et al. (1986), Marsh et al. (1975), Maggon et al. (1977), Shamsudeen et al. (2013) and Davis et al. (1967) who observed that the AF production was inhibited by Fe salts. On the contrary, Cuero and Ouellet, (2005), Cuero et al., (2003) and Aziz and Moussa (1997) observed an increase in AF due to Fe salts. The pH of media was reduced by Fe salts.
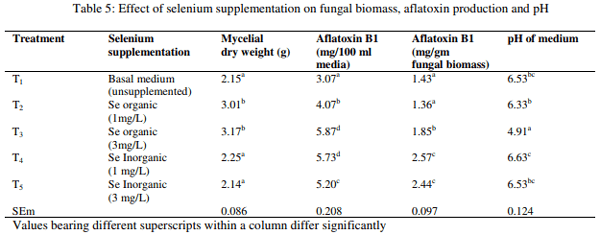
Effect of selenium on fungal growth and aflatoxin production
The fungal growth measured as mean mycelial dry weight, AFB1 production and change in pH of the media as influenced by organic and inorganic Se are presented in Table 5. The result revealed that mycelial dry weight was significantly (p<0.01) increased in basal media with organic Se at both levels than other groups, whereas the media amended with inorganic Se did not differ significantly with control. The AFB1 production in 100 ml of media was significantly (p<0.01) higher in media amended with organic Se at 3 mg/L and inorganic Se at 1 mg/L than the media without Se supplementation, whereas AFB1 production in 100 ml of media of other groups were found to be intermediary. However, the AFB1 production/gm fungal biomass was significantly (p<0.01) higher in media with inorganic Se at both levels followed by media with organic Se at 3 mg/L than other two groups. The pH of media was significantly (p<0.01) higher in media with inorganic Se or control than media with organic Se.
The fungal biomass measured as mycelial dry weight was significantly increased by organic Se than control or inorganic Se and the inorganic Se could not increase fungal growth. The increase in fungal growth might be due to yeast portion of organic Se (Se-yeast). The AF production in 100 ml media was increased by Se supplementation in both forms. However, the AF productions per gm of fungal biomass were increased significantly by inorganic Se than organic Se and the organic Se could not increase the AFB1 per gm of fungal biomass. The result suggests that organic Se might be toxic to the fungi for AF production, even though there was an increased in total AFB1 in media. The final pH of media did not change due to Se supplementation. The studies regarding effect of Se on AF production and fungal growth are lacking in literature.
Effect of zinc on fungal growth and aflatoxin production
The fungal growth measured as mycelial dry weight, AFB1 production and change in pH of the media as influenced by organic and inorganic Zn are presented in Table 6. The result revealed that mycelial dry weight was higher (p<0.01) in media with inorganic Zn at 10 mg/L than media without Zn supplementation and the mycelial dry weight of other groups were found to be intermediary. The total AFB1 production in 100 ml media was higher (p<0.01) in media with inorganic Zn at 20 mg/L than media without Zn supplementation and the total AFB1 production in 100 ml media of other groups were found to be intermediary. Similarly, the AFB1 production per gm of fungal biomass was higher (p<0.01) in media with inorganic Zn at 20 mg/L than media with chelated Zn at 20 mg/L, whereas the AFB1 production/gm of fungal biomass of other groups were found to be intermediary. The pH of media was significantly (p<0.01) higher in media with chelated or inorganic Zn at 10 mg/L than other groups.
The fungal growth measured as mycelial dry weight was increased by Zn supplementation to basal media. Similar results were observed by many authors (Cuero and Ouellet, 2005; Cuero et al., 2003; Aziz and Moussa, 1997; Shamsudeen et al., 2013). Metal ions have a regulatory effect at the cellular and molecular level on fungal growth and concomitant mycotoxin production (Cuero and Ouellet, 2005). However, there was only a little difference between Zn chelate and Zn inorganic suggesting both form might have been utilized by the fungi for fungal growth or the fungal growth might have been induced by traces of Zn in Zn chelate product. The AFB1 production in 100 ml liquid media and AFB1 per gm of fungal biomass was increased significantly by both levels of inorganic Zn.
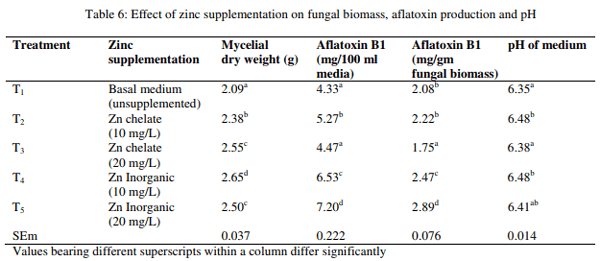
The increased AF due to inorganic Zn was observed by many authors (Cuero and Ouellet, 2005; Cuero et al., 2003; Tiwari et al., 1986). Detroy and Ciegler (1971) suggested that the function of mineral is important as metalloenzyme for enzymatic activity in the production of secondary metabolites. Alcohol dehydrogenase has been shown to be Zn dependent for both regulation of their gene expression and structural stability in yeast (Magonet et al., 1992; Eisen et al., 1988). Addition of Zn has a stimulatory effect on the fatty acid synthetase and in AFB1 production in A. Parasiticus (Reding and Harison, 1994). The AFB1 production was comparable to control at the concentration of 10 mg/L and decreased at the concentration of 20 mg/L, due to Zn chelate. The reduced AF production might possibly due to the unavailability of Zn ion for production of AF by the fungi due to the chelation of the mineral. It is suggested that the effect of mineral on fungal growth and AF production depends on type and concentration of the metal ions (Weinberg, 1982) as well as on the presence of ligand in the substrate (Hughes and Poole, 1989). The pH change of media did not differ between Zn chelate and Zn inorganic group.
Conclusion
It was concluded that there was a difference between chelated and inorganic trace minerals viz., copper, ferrous and zinc in their utilization by the fungi for aflatoxin production which indicates the fungi may not utilize the minerals in chelated form as efficiently as its inorganic counterpart, for aflatoxin production in the feed. The excess selenium either in organic or inorganic form was found to increase the aflatoxin production in liquid medium.
This article was originally published in Journal of Poultry Science and Technology | January-March, 2014 | Vol 2 | Issue 1 | Pages 11-17.