Antifungal Peptides and Proteins to Control Toxigenic Fungi and Mycotoxin Biosynthesis
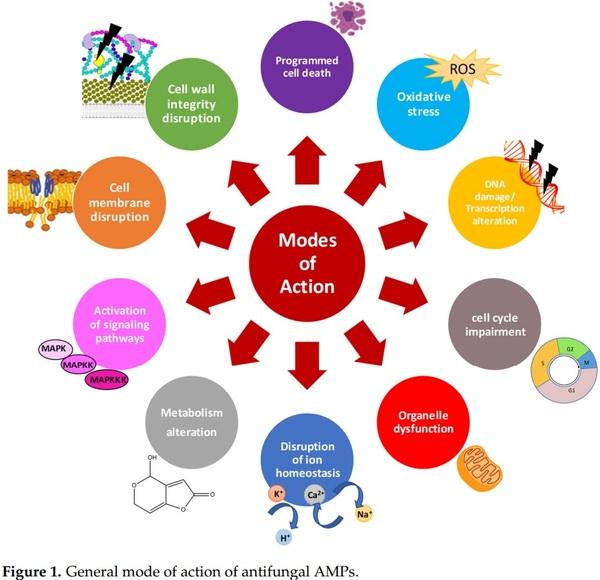
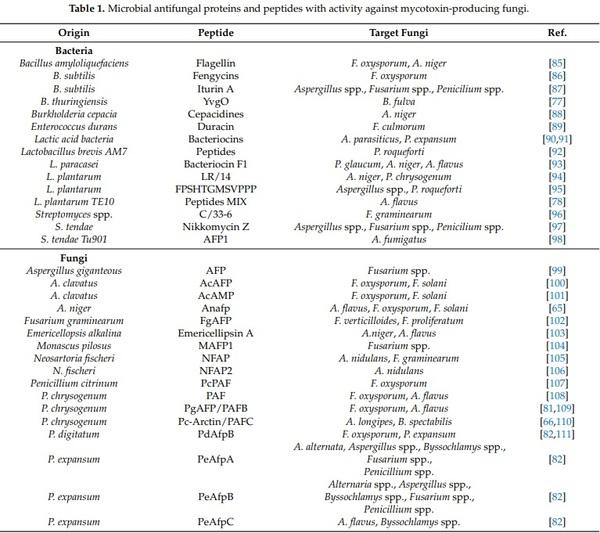
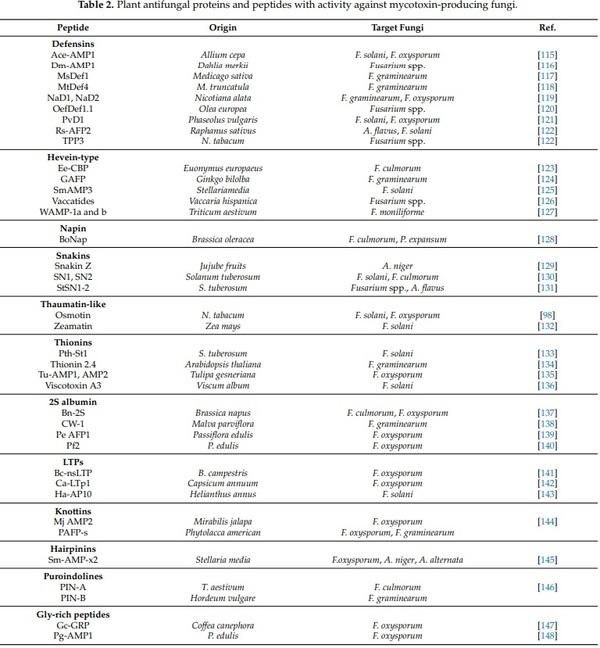
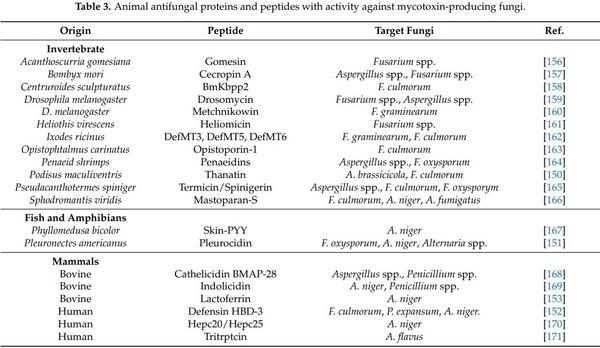


1. Liu, Y.; Yamdeu, J.H.G.; Gong, Y.Y.; Orfila, C. A Review of Postharvest Approaches to Reduce Fungal and Mycotoxin Contamination of Foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1521–1560. [CrossRef] [PubMed]
2. Park, D.L.; Njapau, H.; Boutrif, E. Minimizing Risks Posed by Mycotoxins Utilizing the HACCP Concept. Food Nutr. Agric. 1999,
8, 49–54.
3. Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide Contamination of Food-Crops with Mycotoxins:
Validity of the Widely Cited ‘FAO Estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [CrossRef] [PubMed]
4. Pitt, J.I.; Miller, J.D. A Concise History of Mycotoxin Research. J. Agric. Food Chem. 2017, 65, 7021–7033. [CrossRef]
5. Wu, F.; Mitchell, N.J. How Climate Change and Regulations Can Affect the Economics of Mycotoxins. World Mycotoxin J. 2016, 9,
653–663. [CrossRef]
6. Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, Toxicology, and Exposure Assessment. Food Chem.
Toxicol. 2013, 60, 218–237. [CrossRef]
7. European Commission. Commission Recommendation (2013/165/EU) of 27 March 2013 on the Presence of T-2 and HT-2 Toxin in Cereals and Cereal Products. Off. J. Eur. Union L 2013, 91, 12–15.
8. European Commission. Commission Regulation EC No. 1881/2006. Setting Maximum Levels for Certain Contaminants in
Foodstuffs (N◦ 1881/2006 of 19 December 2006). Off. J. Eur. Union 2006, 364, 5–24.
9. Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [CrossRef]
10. Smith, M.-C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in Vitro
Combined Toxicological Effects. Toxins 2016, 8, 94. [CrossRef]
11. Hua, Z.; Liu, R.; Chen, Y.; Liu, G.; Li, C.; Song, Y.; Cao, Z.; Li, W.; Li, W.; Lu, C.; et al. Contamination of Aflatoxins Induces Severe
Hepatotoxicity Through Multiple Mechanisms. Front. Pharmacol. 2021, 11, 2144. [CrossRef] [PubMed]
12. Squire, R.A. Ranking Animal Carcinogens: A Proposed Regulatory Approach. Science 1981, 214, 877–880. [CrossRef] [PubMed]
13. International Agency for Research on Cancer. IARC monographs on the evaluation of carcinogenic risk of chemicals to man.
IARC Monogr. Eval. Carcinog. Risk Chem. Man. 1972, 1, 1–184.
14. Bui-Klimke, T.R.; Wu, F. Ochratoxin A and Human Health Risk: A Review of the Evidence. Crit. Rev. Food Sci. Nutr. 2015, 55,
1860–1869. [CrossRef]
15. Koszegi, T.; Poór, M. Ochratoxin A: Molecular Interactions, Mechanisms of Toxicity and Prevention at the Molecular Level. Toxins
2016, 8, 111. [CrossRef] [PubMed]
16. Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and Its Metabolites: Effect on Human Health,
Metabolism and Neutralisation Methods. Toxicon 2019, 162, 46–56. [CrossRef] [PubMed]
17. Polak-Sliwi ´nska, M.; Paszczyk, B. Trichothecenes in Food and Feed, Relevance to Human and Animal Health and Methods of ´
Detection: A Systematic Review. Molecules 2021, 26, 454. [CrossRef] [PubMed]
18. Voss, K.A.; Riley, R.T. Fumonisin Toxicity and Mechanism of Action: Overview and Current Perspectives. Food Saf. 2013,
1, 2013006. [CrossRef]
19. Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [CrossRef]
20. Hahn, M. The Rising Threat of Fungicide Resistance in Plant Pathogenic Fungi: Botrytis as a Case Study. J. Chem. Biol. 2014, 7,
133–141. [CrossRef]
21. Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide Emergence of Resistance to Antifungal Drugs Challenges Human
Health and Food Security. Science 2018, 360, 739–742. [CrossRef]
22. Marín, P.; de Ory, A.; Cruz, A.; Magan, N.; González-Jaén, M.T. Potential Effects of Environmental Conditions on the Efficiency of the Antifungal Tebuconazole Controlling Fusarium verticillioides and Fusarium proliferatum Growth Rate and Fumonisin
Biosynthesis. Int. J. Food Microbiol. 2013, 165, 251–258. [CrossRef]
23. Mateo, E.M.; Valle-Algarra, F.M.; Mateo, R.; Jiménez, M.; Magan, N. Effect of Fenpropimorph, Prochloraz and Tebuconazole on
Growth and Production of T-2 and HT-2 Toxins by Fusarium langsethiae in Oat-Based Medium. Int. J. Food Microbiol. 2011, 151,
289–298. [CrossRef]
24. Hoppin, J.A.; Umbach, D.M.; Long, S.; London, S.J.; Henneberger, P.K.; Blair, A.; Alavanja, M.; Freeman, L.E.B.; Sandler, D.P.
Pesticides Are Associated with Allergic and Non-Allergic Wheeze among Male Farmers. Environ. Health Perspect. 2017, 125,
535–543. [CrossRef] [PubMed]
25. Juntarawijit, C.; Juntarawijit, Y. Association between Diabetes and Pesticides: A Case-Control Study among Thai Farmers. Environ.
Health Prev. Med. 2018, 23, 3. [CrossRef] [PubMed]
26. Piel, C.; Pouchieu, C.; Carles, C.; Béziat, B.; Boulanger, M.; Bureau, M.; Busson, A.; Grüber, A.; Lecluse, Y.; Migault, L.; et al.
Agricultural Exposures to Carbamate Herbicides and Fungicides and Central Nervous System Tumour Incidence in the Cohort
AGRICAN. Environ. Int. 2019, 130, 104876. [CrossRef]
27. Harris, C.A.; Tomerlin, J.R. The Regulation of Pesticides in Europe–Directive 91/414. J. Environ. Monit. 2002, 4, 28N–31N.
28. Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to Pesticides and the Associated Human Health Effects. Sci. Total Environ. 2017, 575,
525–535. [CrossRef]
29. Guynot, M.E.; Ramos, A.J.; Sanchis, V.; Marín, S. Study of Benzoate, Propionate, and Sorbate Salts as Mould Spoilage Inhibitors on Intermediate Moisture Bakery Products of Low PH (4.5-5.5). Int. J. Food Microbiol. 2005, 101, 161–168. [CrossRef]
30. Marín, S.; Magan, N.; Abellana, M.; Canela, R.; Ramos, A.J.; Sanchis, V. Selective Effect of Propionates and Water Activity on
Maize Mycoflora and Impact on Fumonisin B1 Accumulation. J. Stored Prod. Res. 2000, 36, 203–214. [CrossRef]
31. Hauser, C.; Thielmann, J.; Muranyi, P. Organic Acids: Usage and Potential in Antimicrobial Packaging. In Antimicrobial Food
Packaging; Barros-Velázquez, J., Ed.; Academic Press: San Diego, CA, USA, 2016; Chapter 46; pp. 563–580. ISBN 978-0-12-800723-5.
32. Schmidt-Heydt, M.; Baxter, E.; Geisen, R.; Magan, N. Physiological Relationship between Food Preservatives, Environmental
Factors, Ochratoxin and OtapksPV Gene Expression by Penicillium verrucosum. Int. J. Food Microbiol. 2007, 119, 277–283. [CrossRef]
[PubMed]
33. Leyva Salas, M.; Mounier, J.; Valence, F.; Coton, M.; Thierry, A.; Coton, E. Antifungal Microbial Agents for Food Biopreservation—
A Review. Microorganisms 2017, 5, 37. [CrossRef] [PubMed]
34. Fernández de Ullivarri, M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell.
Infect. Microbiol. 2020, 10, 105. [CrossRef] [PubMed]
35. Marx, F.; Binder, U.; Leiter, É.; Pócsi, I. The Penicillium chrysogenum Antifungal Protein PAF, a Promising Tool for the Development of New Antifungal Therapies and Fungal Cell Biology Studies. Cell. Mol. Life Sci. 2008, 65, 445–454. [CrossRef] [PubMed]
36. Meyer, V. A Small Protein That Fights Fungi: AFP as a New Promising Antifungal Agent of Biotechnological Value. Appl.
Microbiol. Biotechnol. 2008, 78, 17–28. [CrossRef] [PubMed]
37. Thery, T.; Lynch, K.M.; Arendt, E.K. Natural Antifungal Peptides/Proteins as Model for Novel Food Preservatives. Compr. Rev.
Food Sci. Food Saf. 2019, 18, 1327–1360. [CrossRef] [PubMed]
38. Rajasekaran, K.; Sayler, R.J.; Sickler, C.M.; Majumdar, R.; Jaynes, J.M.; Cary, J.W. Control of Aspergillus flavus Growth and Aflatoxin
Production in Transgenic Maize Kernels Expressing a Tachyplesin-Derived Synthetic Peptide, AGM182. Plant. Sci. 2018, 270,
150–156. [CrossRef] [PubMed]
39. Rajasekaran, K.; Cary, J.W.; Chlan, C.A.; Jaynes, J.M.; Bhatnagar, D. Strategies for Controlling Plant Diseases and Mycotoxin
Contamination Using Antimicrobial Synthetic Peptides. In ACS Symposium Series; Rajasekaran, K., Cary, J.W., Jaynes, J.M.,
Montesinos, E., Eds.; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, pp. 295–315. ISBN 978-0-8412-2748-4.
40. Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al.
Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 453. [CrossRef]
41. Sarkar, T.; Chetia, M.; Chatterjee, S. Antimicrobial Peptides and Proteins: From Nature’s Reservoir to the Laboratory and Beyond.
Front. Chem. 2021, 9, 432. [CrossRef]
42. Delgado, J.; Owens, R.A.; Doyle, S.; Asensio, M.A.; Núñez, F. Manuscript Title: Antifungal Proteins from Moulds: Analytical
Tools and Potential Application to Dry-Ripened Foods. Appl. Microbiol. Biotechnol. 2016, 100, 6991–7000. [CrossRef] [PubMed]
43. Shwaiki, L.N.; Lynch, K.M.; Arendt, E.K. Future of Antimicrobial Peptides Derived from Plants in Food Application—A Focus on
Synthetic Peptides. Trends Food Sci. Technol. 2021, 112, 312–324. [CrossRef]
44. Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a New Generation of Antimicrobials: Toxicity Aspects and Regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [CrossRef]
45. Hegedüs, N.; Marx, F. Antifungal Proteins: More than Antimicrobials? Fungal Biol. Rev. 2013, 26, 132–145. [CrossRef] [PubMed]
46. Paege, N.; Jung, S.; Schäpe, P.; Müller-Hagen, D.; Ouedraogo, J.-P.; Heiderich, C.; Jedamzick, J.; Nitsche, B.M.; van den Hondel,
C.A.; Ram, A.F.; et al. A Transcriptome Meta-Analysis Proposes Novel Biological Roles for the Antifungal Protein AnAFP in
Aspergillus niger. PLoS ONE 2016, 11, e0165755. [CrossRef]
47. Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute,
S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. Lancet Infect. Dis. 2020, 20, e216–e230.
[CrossRef]
48. Van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Properties and Mechanisms of Action of Naturally Occurring Antifungal
Peptides. Cell. Mol. Life Sci. 2013, 70, 3545–3570. [CrossRef]
49. Wang, G.; Li, X.; Wang, Z. APD3: The Antimicrobial Peptide Database as a Tool for Research and Education. Nucleic Acids Res.
2016, 44, D1087–D1093. [CrossRef] [PubMed]
50. Do Nascimento Dias, J.; de Souza Silva, C.; de Araújo, A.R.; Souza, J.M.T.; de Holanda Veloso Júnior, P.H.; Cabral, W.F.; da Glória da Silva, M.; Eaton, P.; de Souza de Almeida Leite, J.R.; Nicola, A.M.; et al. Mechanisms of Action of Antimicrobial Peptides
ToAP2 and NDBP-5.7 against Candida albicans Planktonic and Biofilm Cells. Sci. Rep. 2020, 10, 10327. [CrossRef]
51. Parvy, J.-P.; Yu, Y.; Dostalova, A.; Kondo, S.; Kurjan, A.; Bulet, P.; Lemaître, B.; Vidal, M.; Cordero, J.B. The Antimicrobial Peptide
Defensin Cooperates with Tumour Necrosis Factor to Drive Tumour Cell Death in Drosophila. eLife 2019, 8, e45061. [CrossRef]
52. Buda De Cesare, G.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial Peptides: A New Frontier in Antifungal Therapy. mBio
2020, 11, e02123-20. [CrossRef]
53. Struyfs, C.; Cools, T.L.; De Cremer, K.; Sampaio-Marques, B.; Ludovico, P.; Wasko, B.M.; Kaeberlein, M.; Cammue, B.P.A.;
Thevissen, K. The Antifungal Plant Defensin HsAFP1 Induces Autophagy, Vacuolar Dysfunction and Cell Cycle Impairment in
Yeast. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183255. [CrossRef] [PubMed]
54. Finking, R.; Marahiel, M.A. Biosynthesis of Nonribosomal Peptides. Annu. Rev. Microbiol. 2004, 58, 453–488. [CrossRef]
55. Wang, G. Post-Translational Modifications of Natural Antimicrobial Peptides and Strategies for Peptide Engineering. Curr.
Biotechnol. 2012, 1, 72–79. [CrossRef] [PubMed]
56. Brogden, K.A. Antimicrobial Peptides: Pore Formers or Metabolic Inhibitors in Bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250.
[CrossRef]
57. Marcos, J.F.; Gandía, M. Antimicrobial Peptides: To Membranes and Beyond. Expert Opin. Drug Discov. 2009, 4, 659–671.
[CrossRef] [PubMed]
58. Nicolas, P. Multifunctional Host Defense Peptides: Intracellular-Targeting Antimicrobial Peptides. FEBS J. 2009, 276, 6483–6496.
[CrossRef]
59. Rautenbach, M.; Troskie, A.M.; Vosloo, J.A. Antifungal Peptides: To Be or Not to Be Membrane Active. Biochimie 2016, 130,
132–145. [CrossRef] [PubMed]
60. Nguyen, L.; Haney, E.; Vogel, H. The Expanding Scope of Antimicrobial Peptide Structures and Their Modes of Action. Trends
Biotechnol. 2011, 29, 464–472. [CrossRef]
61. Zasloff, M. Antimicrobial Peptides of Multicellular Organisms. Nature 2002, 415, 389–395. [CrossRef] [PubMed]
62. Guilhelmelli, F.; Vilela, N.; Albuquerque, P.; Derengowski, L.D.S.; Silva-Pereira, I.; Kyaw, C.M. Antibiotic Development Challenges:
The Various Mechanisms of Action of Antimicrobial Peptides and of Bacterial Resistance. Front. Microbiol. 2013, 4, 353. [CrossRef]
63. Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by
Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 302. [CrossRef] [PubMed]
64. Jiao, R.; Cai, Y.; He, P.; Munir, S.; Li, X.; Wu, Y.; Wang, J.; Xia, M.; He, P.; Wang, G.; et al. Bacillus amyloliquefaciens YN201732
Produces Lipopeptides With Promising Biocontrol Activity Against Fungal Pathogen Erysiphe Cichoracearum. Front. Cell. Infect.
Microbiol. 2021, 11, 387. [CrossRef]
65. Gun Lee, D.; Shin, S.Y.; Maeng, C.Y.; Jin, Z.Z.; Kim, K.L.; Hahm, K.S. Isolation and Characterization of a Novel Antifungal Peptide from Aspergillus niger. Biochem. Biophys. Res. Commun. 1999, 263, 646–651. [CrossRef]
66. Chen, Z.; Ao, J.; Yang, W.; Jiao, L.; Zheng, T.; Chen, X. Purification and Characterization of a Novel Antifungal Protein Secreted by
Penicillium chrysogenum from an Arctic Sediment. Appl. Microbiol. Biotechnol. 2013, 97, 10381–10390. [CrossRef]
67. Moreno, A.B.; Martínez Del Pozo, A.; San Segundo, B. Biotechnologically Relevant Enzymes and Proteins. Antifungal Mechanism of the Aspergillus giganteus AFP against the Rice Blast Fungus Magnaporthe Grisea. Appl. Microbiol. Biotechnol. 2006, 72, 883–895.
[CrossRef] [PubMed]
68. Silva, P.M.; Gonçalves, S.; Santos, N.C. Defensins: Antifungal Lessons from Eukaryotes. Front. Microbiol. 2014, 5, 97. [CrossRef]
69. Meyer, V.; Jung, S. Antifungal Peptides of the AFP Family Revisited: Are These Cannibal Toxins? Microorganisms 2018, 6, 50.
[CrossRef] [PubMed]
70. Batta, G.; Barna, T.; Gáspári, Z.; Sándor, S.; Kövér, K.E.; Binder, U.; Sarg, B.; Kaiserer, L.; Chhillar, A.K.; Eigentler, A.; et al.
Functional Aspects of the Solution Structure and Dynamics of PAF—A Highly-Stable Antifungal Protein from Penicillium chrysogenum. FEBS J. 2009, 276, 2875–2890. [CrossRef] [PubMed]
71. Campos-Olivas, R.; Bruix, M.; Santoro, J.; Lacadena, J.; Martinez del Pozo, A.; Gavilanes, J.G.; Rico, M. NMR Solution Structure of the Antifungal Protein from Aspergillus giganteus: Evidence for Cysteine Pairing Isomerism. Biochemistry 1995, 34, 3009–3021.
[CrossRef] [PubMed]
72. Yount, N.Y.; Yeaman, M.R. Multidimensional Signatures in Antimicrobial Peptides. Proc. Natl. Acad. Sci. USA 2004, 101,
7363–7368. [CrossRef]
73. Marcos, J.F.; Manzanares, P. Antimicrobial Peptides. In Antimicrobial Polymers; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp.
195–212. ISBN 978-0-470-59822-1.
74. Kerenga, B.K.; McKenna, J.A.; Harvey, P.J.; Quimbar, P.; Garcia-Ceron, D.; Lay, F.T.; Phan, T.K.; Veneer, P.K.; Vasa, S.; Parisi, K.; et al. Salt-Tolerant Antifungal and Antibacterial Activities of the Corn Defensin ZmD32. Front. Microbiol. 2019, 10, 795. [CrossRef]
75. López-García, B.; Harries, E.; Carmona, L.; Campos-Soriano, L.; López, J.J.; Manzanares, P.; Gandía, M.; Coca, M.; Marcos,
J.F. Concatemerization Increases the Inhibitory Activity of Short, Cell-Penetrating, Cationic and Tryptophan-Rich Antifungal
Peptides. Appl. Microbiol. Biotechnol. 2015, 99, 8011–8021. [CrossRef]
76. Marcos, J.F.; Muñoz, A.; Pérez-Payá, E.; Misra, S.; López-García, B. Identification and Rational Design of Novel Antimicrobial
Peptides for Plant Protection. Annu. Rev. Phytopathol. 2008, 46, 273–301. [CrossRef] [PubMed]
77. Manns, D.C.; Churey, J.J.; Worobo, R.W. Variable Efficacy of the Proteinaceous Antifungal YvgO in Select Fruit Juices and Teas as a Complement with UV Methods of Food Protection. J. Food Prot. 2015, 78, 1851–1860. [CrossRef] [PubMed]
78. Muhialdin, B.J.; Algboory, H.L.; Kadum, H.; Mohammed, N.K.; Saari, N.; Hassan, Z.; Meor Hussin, A.S. Antifungal Activity
Determination for the Peptides Generated by Lactobacillus plantarum TE10 against Aspergillus flavus in Maize Seeds. Food Control.
2020, 109, 106898. [CrossRef]
79. Garrigues, S.; Gandía, M.; Marcos, J.F. Occurrence and Function of Fungal Antifungal Proteins: A Case Study of the Citrus
Postharvest Pathogen Penicillium digitatum. Appl. Microbiol. Biotechnol. 2016, 100, 2243–2256. [CrossRef] [PubMed]
80. Leiter, É.; Gáll, T.; Csernoch, L.; Pócsi, I. Biofungicide Utilizations of Antifungal Proteins of Filamentous Ascomycetes: Current and Foreseeable Future Developments. BioControl 2017, 62, 125–138. [CrossRef]
81. Delgado, J.; Acosta, R.; Rodríguez-Martín, A.; Bermúdez, E.; Núñez, F.; Asensio, M.A. Growth Inhibition and Stability of PgAFP from Penicillium chrysogenum against Fungi Common on Dry-Ripened Meat Products. Int. J. Food Microbiol. 2015, 205, 23–29.
[CrossRef]
82. Martínez-Culebras, P.V.; Gandía, M.; Boronat, A.; Marcos, J.F.; Manzanares, P. Differential Susceptibility of Mycotoxin-Producing
Fungi to Distinct Antifungal Proteins (AFPs). Food Microbiol. 2021, 97, 103760. [CrossRef]
83. Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: New York, NY, USA, 2009; p. 519. ISBN 978-0-387-92206-5.
84. Delgado, J.; Ballester, A.-R.; Núñez, F.; González-Candelas, L. Evaluation of the Activity of the Antifungal PgAFP Protein and
Its Producer Mould against Penicillium spp. Postharvest Pathogens of Citrus and Pome Fruits. Food Microbiol. 2019, 84, 103266.
[CrossRef] [PubMed]
85. Zhao, P.; Quan, C.; Wang, Y.; Wang, J.; Fan, S. Bacillus amyloliquefaciens Q-426 as a Potential Biocontrol Agent against Fusarium oxysporum f. sp. spinaciae. J. Basic Microbiol. 2014, 54, 448–456. [CrossRef] [PubMed]
86. Vanittanakom, N.; Loeffler, W.; Koch, U.; Jung, G. Fengycin-a novel antifungal lipopeptide antibiotic produced by Bacillus subtilis
F-29-3. J. Antibiot. 1986, 39, 888–901. [CrossRef] [PubMed]
87. Klich, M.; Lax, A.; Bland, J. Inhibition of Some Mycotoxigenic Fungi by Iturin A, a Peptidolipid Produced by Bacillus Subtilis.
Mycopathologia 1991, 116, 77–80. [CrossRef]
88. Lim, Y.; Suh, J.W.; Kim, S.; Hyun, B.; Kim, C.; Lee, C.H. Cepacidine A, a Novel Antifungal Antibiotic Produced by Pseudomonas
Cepacia. II. Physico-Chemical Properties and Structure Elucidation. J. Antibiot. 1994, 47, 1406–1416. [CrossRef]
89. Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Baudy-Floc’h, M.; Le Blay, G.; Haertlé, T.; Chobert, J.-M. Antifungal Properties of
Durancins Isolated from Enterococcus durans A5-11 and of Its Synthetic Fragments. Lett. Appl. Microbiol. 2012, 56. [CrossRef]
[PubMed]
90. Daie Ghazvini, R.; Kouhsari, E.; Zibafar, E.; Hashemi, J.; Amini, A.; Niknejad, F. Antifungal Activity and Aflatoxin Degradation of
Bifidobacterium Bifidum and Lactobacillus Fermentum Against Toxigenic Aspergillus parasiticus. Open Microbiol. J. 2016, 10, 1–5.
[CrossRef]
91. Luz, C.; Saladino, F.; Luciano, F.B.; Mañes, J.; Meca, G. In Vitro Antifungal Activity of Bioactive Peptides Produced by Lactobacillus plantarum against Aspergillus parasiticus and Penicillium expansum. LWT—Food Sci. Technol. 2017, 81, 128–135. [CrossRef]
92. Coda, R.; Rizzello, C.G.; Nigro, F.; De Angelis, M.; Arnault, P.; Gobbetti, M. Long-Term Fungal Inhibitory Activity of Water-Soluble
Extracts of Phaseolus Vulgaris Cv. Pinto and Sourdough Lactic Acid Bacteria during Bread Storage. Appl. Environ. Microbiol. 2008,
74, 7391–7398. [CrossRef]
93. Miao, J.; Guo, H.; Ou, Y.; Liu, G.; Fang, X.; Liao, Z.; Ke, C.; Chen, Y.; Zhao, L.; Cao, Y. Purification and Characterization of
Bacteriocin F1, a Novel Bacteriocin Produced by Lactobacillus paracasei subsp. Tolerans FX-6 from Tibetan Kefir, a Traditional
Fermented Milk from Tibet, China. Food Control. 2014, 42, 48–53. [CrossRef]
94. Gupta, R.; Srivastava, S. Antifungal Effect of Antimicrobial Peptides (AMPs LR14) Derived from Lactobacillus plantarum Strain
LR/14 and Their Applications in Prevention of Grain Spoilage. Food Microbiol. 2014, 42, 1–7. [CrossRef]
95. Muhialdin, B.J.; Hassan, Z.; Bakar, F.A.; Saari, N. Identification of Antifungal Peptides Produced by Lactobacillus plantarum IS10
Grown in the MRS Broth. Food Control. 2016, 59, 27–30. [CrossRef]
96. Fulgueira, C.L.; Amigot, S.L.; Magni, C. Growth Inhibition of Toxigenic Fungi by a Proteinaceous Compound from Streptomyces sp. C/33-6. Curr. Microbiol. 2004, 48, 135–139. [CrossRef]
97. Li, R.K.; Rinaldi, M.G. In Vitro Antifungal Activity of Nikkomycin Z in Combination with Fluconazole or Itraconazole. Antimicrob.
Agents Chemother. 1999, 43, 1401–1405. [CrossRef] [PubMed]
98. Freitas, C.; Nogueira, F.; Vasconcelos, I.; Oliveira, J.; Domont, G.; Ramos, M. Osmotin Purified from the Latex of Calotropis procera:
Biochemical Characterization, Biological Activity and Role in Plant Defense. Plant. Physiol. Biochem. PPB/Soc. Fr. De Physiol. Veg.
2011, 49, 738–743. [CrossRef]
99. Theis, T.; Marx, F.; Salvenmoser, W.; Stahl, U.; Meyer, V. New Insights into the Target Site and Mode of Action of the Antifungal
Protein of Aspergillus giganteus. Res. Microbiol. 2005, 156, 47–56. [CrossRef]
100. Skouri-Gargouri, H.; Gargouri, A. First Isolation of a Novel Thermostable Antifungal Peptide Secreted by Aspergillus clavatus.
Peptides 2008, 29, 1871–1877. [CrossRef]
101. Mohamed, H.; Jellouli, K.; Hmidet, N.; Balti, R.; Sellami-Kamoun, A. A Highly Thermostable Antimicrobial Peptide from
Aspergillus clavatus ES1: Biochemical and Molecular Characterization. J. Ind. Microbiol. Biotechnol. 2010, 37, 805–813. [CrossRef]
102. Patiño, B.; Vázquez, C.; Manning, J.M.; Roncero, M.I.G.; Córdoba-Cañero, D.; Di Pietro, A.; Martínez-del-Pozo, Á. Characterization of a Novel Cysteine-Rich Antifungal Protein from Fusarium graminearum with Activity against Maize Fungal Pathogens. Int. J.
Food Microbiol. 2018, 283, 45–51. [CrossRef] [PubMed]
103. Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’ko, A.B.;
Krasheninnikov, M.E.; Lyundup, A.V.; et al. A Novel Lipopeptaibol Emericellipsin A with Antimicrobial and Antitumor Activity
Produced by the Extremophilic Fungus Emericellopsis alkalina. Molecules 2018, 23, 2785. [CrossRef] [PubMed]
104. Tu, C.-Y.; Chen, Y.-P.; Yu, M.-C.; Hwang, I.-E.; Wu, D.-Y.; Liaw, L.-L. Characterization and Expression of the Antifungal Protein from Monascus pilosus and Its Distribution among Various Monascus Species. J. Biosci. Bioeng. 2016, 122, 27–33. [CrossRef]
[PubMed]
105. Kovács, L.; Virágh, M.; Takó, M.; Papp, T.; Vágvölgyi, C.; Galgóczy, L. Isolation and Characterization of Neosartorya fischeri
Antifungal Protein (NFAP). Peptides 2011, 32, 1724–1731. [CrossRef]
106. Tóth, L.; Kele, Z.; Borics, A.; Nagy, L.G.; Váradi, G.; Virágh, M.; Takó, M.; Vágvölgyi, C.; Galgóczy, L. NFAP2, a Novel
Cysteine-Rich Anti-Yeast Protein from Neosartorya fischeri NRRL 181: Isolation and Characterization. AMB Express 2016, 6, 75.
[CrossRef]
107. Wen, C.; Guo, W.; Chen, X. Purification and Identification of a Novel Antifungal Protein Secreted by Penicillium citrinum from the
Southwest Indian Ocean. J. Microbiol. Biotechnol. 2014, 24, 1337–1345. [CrossRef]
108. Kaiserer, L.; Oberparleiter, C.; Weiler-Görz, R.; Burgstaller, W.; Leiter, E.; Marx, F. Characterization of the Penicillium chrysogenum
Antifungal Protein PAF. Arch. Microbiol. 2003, 180, 204–210. [CrossRef] [PubMed]
109. Huber, A.; Hajdu, D.; Bratschun-Khan, D.; Gáspári, Z.; Varbanov, M.; Philippot, S.; Fizil, Á.; Czajlik, A.; Kele, Z.; Sonderegger,
C.; et al. New Antimicrobial Potential and Structural Properties of PAFB: A Cationic, Cysteine-Rich Protein from Penicillium chrysogenum Q176. Sci. Rep. 2018, 8, 1751. [CrossRef]
110. Holzknecht, J.; Kühbacher, A.; Papp, C.; Farkas, A.; Váradi, G.; Marcos, J.F.; Manzanares, P.; Tóth, G.K.; Galgóczy, L.; Marx, F. The
Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen
Candida albicans. JoF 2020, 6, 141. [CrossRef]
111. Garrigues, S.; Gandía, M.; Popa, C.; Borics, A.; Marx, F.; Coca, M.; Marcos, J.F.; Manzanares, P. Efficient Production and
Characterization of the Novel and Highly Active Antifungal Protein AfpB from Penicillium digitatum. Sci. Rep. 2017, 7, 14663.
[CrossRef]
112. Gandía, M.; Monge, A.; Garrigues, S.; Orozco, H.; Giner-Llorca, M.; Marcos, J.F.; Manzanares, P. Novel Insights in the Production,
Activity and Protective Effect of Penicillium expansum Antifungal Proteins. Int. J. Biol. Macromol. 2020, 164, 3922–3931. [CrossRef]
[PubMed]
113. Marcos López, J.F.; Gandía Gómez, M.; Garrigues, S.; Manzanares, P.; Coca, M. Antifungal Peptides and Proteins with Activity against
Fungi Causing Postharvest Decay; Taylor & Francis: New York, NY, USA, 2020; ISBN 978-1-315-20918-0.
114. Yan, J.; Yuan, S.-S.; Jiang, L.-L.; Ye, X.-J.; Ng, T.; Wu, Z.-J. Plant Antifungal Proteins and Their Applications in Agriculture. Appl.
Microbiol. Biotechnol. 2015, 99, 4961–4981. [CrossRef]
115. Wu, Y.; He, Y.; Ge, X. Functional Characterization of the Recombinant Antimicrobial Peptide Trx-Ace-AMP1 and Its Application on the Control of Tomato Early Blight Disease. Appl. Microbiol. Biotechnol. 2011, 90, 1303–1310. [CrossRef] [PubMed]
116. Sagaram, U.S.; El-Mounadi, K.; Buchko, G.W.; Berg, H.R.; Kaur, J.; Pandurangi, R.S.; Smith, T.J.; Shah, D.M. Structural and
Functional Studies of a Phosphatidic Acid-Binding Antifungal Plant Defensin MtDef4: Identification of an RGFRRR Motif
Governing Fungal Cell Entry. PLoS ONE 2013, 8, e82485. [CrossRef]
117. Cruz, L.P.; Ribeiro, S.F.F.; Carvalho, A.O.; Vasconcelos, I.M.; Rodrigues, R.; Cunha, M.D.; Gomes, V.M. Isolation and Partial
Characterization of a Novel Lipid Transfer Protein (LTP) and Antifungal Activity of Peptides from Chilli Pepper Seeds. Protein
Pept. Lett. 2010, 17, 311–318. [CrossRef]
118. Kaur, J.; Thokala, M.; Robert-Seilaniantz, A.; Zhao, P.; Peyret, H.; Berg, H.; Pandey, S.; Jones, J.; Shah, D. Subcellular Targeting of an Evolutionarily Conserved Plant Defensin MtDef4.2 Determines the Outcome of Plant-Pathogen Interaction in Transgenic
Arabidopsis. Mol. Plant. Pathol. 2012, 13, 1032–1046. [CrossRef] [PubMed]
119. Dracatos, P.M.; van der Weerden, N.L.; Carroll, K.T.; Johnson, E.D.; Plummer, K.M.; Anderson, M.A. Inhibition of Cereal Rust
Fungi by Both Class I and II Defensins Derived from the Flowers of Nicotiana alata. Mol. Plant. Pathol. 2013, 15, 67–79. [CrossRef]
[PubMed]
120. Li, H.; Velivelli, S.; Shah, D. Antifungal Potency and Modes of Action of a Novel Olive Tree Defensin Against Closely Related
Ascomycete Fungal Pathogens. Mol. Plant.-Microbe Interact. 2019, 32, 1649–1664. [CrossRef] [PubMed]
121. Games, P.D.; Dos Santos, I.S.; Mello, E.O.; Diz, M.S.S.; Carvalho, A.O.; de Souza-Filho, G.A.; Da Cunha, M.; Vasconcelos, I.M.;
Ferreira, B.D.S.; Gomes, V.M. Isolation, Characterization and Cloning of a CDNA Encoding a New Antifungal Defensin from
Phaseolus vulgaris L. Seeds. Peptides 2008, 29, 2090–2100. [CrossRef] [PubMed]
122. Baxter, A.A.; Richter, V.; Lay, F.T.; Poon, I.K.H.; Adda, C.G.; Veneer, P.K.; Phan, T.K.; Bleackley, M.R.; Anderson, M.A.; Kvansakul,
M.; et al. The Tomato Defensin TPP3 Binds Phosphatidylinositol (4,5)-Bisphosphate via a Conserved Dimeric Cationic Grip
Conformation to Mediate Cell Lysis. Mol. Cell. Biol. 2015, 35, 1964–1978. [CrossRef]
123. Van den Bergh, K.P.B.; Proost, P.; Van Damme, J.; Coosemans, J.; Van Damme, E.J.M.; Peumans, W.J. Five Disulfide Bridges
Stabilize a Hevein-Type Antimicrobial Peptide from the Bark of Spindle Tree (Euonymus europaeus L.). FEBS Lett. 2002, 530,
181–185. [CrossRef]
124. Wong, K.H.; Tan, W.L.; Kini, S.G.; Xiao, T.; Serra, A.; Sze, S.K.; Tam, J.P. Vaccatides: Antifungal Glutamine-Rich Hevein-Like
Peptides from Vaccaria hispanica. Front. Plant. Sci. 2017, 8, 1100. [CrossRef]
125. Rogozhin, E.; Slezina, M.; Slavokhotova, A.; Istomina, E.; Korostyleva, T.; Smirnov, A.; Grishin, E.; Egorov, T.; Odintsova, T. A
Novel Antifungal Peptide from Leaves of the Weed Stellaria media L. Biochimie 2015, 116, 125–132. [CrossRef]
126. Huang, R.-H.; Xiang, Y.; Liu, X.-Z.; Zhang, Y.; Hu, Z.; Wang, D.-C. Two Novel Antifungal Peptides Distinct with a Five-Disulfide
Motif from the Bark of Eucommia ulmoides Oliv. FEBS Lett 2002, 521, 87–90. [CrossRef]
127. Odintsova, T.I.; Vassilevski, A.A.; Slavokhotova, A.A.; Musolyamov, A.K.; Finkina, E.I.; Khadeeva, N.V.; Rogozhin, E.A.;
Korostyleva, T.V.; Pukhalsky, V.A.; Grishin, E.V.; et al. A Novel Antifungal Hevein-Type Peptide from Triticum kiharae Seeds with a Unique 10-Cysteine Motif. FEBS J. 2009, 276, 4266–4275. [CrossRef]
128. Thery, T.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Isolation, Characterisation and Application of a New Antifungal Protein from
Broccoli Seeds—New Food Preservative with Great Potential. Food Control. 2020, 117, 107356. [CrossRef]
129. Daneshmand, F.; Zare-Zardini, H.; Ebrahimi, L. Investigation of the Antimicrobial Activities of Snakin-Z, a New Cationic Peptide
Derived from Zizyphus jujuba Fruits. Nat. Prod. Res. 2013, 27, 2292–2296. [CrossRef] [PubMed]
130. Bártová, V.; Bárta, J.; Jarošová, M. Antifungal and Antimicrobial Proteins and Peptides of Potato (Solanum tuberosum L.) Tubers and Their Applications. Appl. Microbiol. Biotechnol. 2019, 103, 5533–5547. [CrossRef]
131. Berrocal-Lobo, M.; Segura, A.; Moreno, M.; López, G.; García-Olmedo, F.; Molina, A. Snakin-2, an Antimicrobial Peptide from
Potato Whose Gene Is Locally Induced by Wounding and Responds to Pathogen Infection. Plant. Physiol. 2002, 128, 951–961.
[CrossRef] [PubMed]
132. Abad, L.R.; D’Urzo, M.P.; Liu, D.; Narasimhan, M.L.; Reuveni, M.; Zhu, J.K.; Niu, X.; Singh, N.K.; Hasegawa, P.M.; Bressan, R.A.
Antifungal Activity of Tobacco Osmotin Has Specificity and Involves Plasma Membrane Permeabilization. Plant. Sci. 1996, 118,
11–23. [CrossRef]
133. Moreno, M.; Segura, A.; García-Olmedo, F. Pseudothionin-St1, a Potato Peptide Active against Potato Pathogens. Eur. J. Biochem.
1994, 223, 135–139. [CrossRef]
134. Asano, T.; Miwa, A.; Maeda, K.; Kimura, M.; Nishiuchi, T. The Secreted Antifungal Protein Thionin 2.4 in Arabidopsis thaliana
Suppresses the Toxicity of a Fungal Fruit Body Lectin from Fusarium graminearum. PLOS Pathog. 2013, 9, e1003581. [CrossRef]
[PubMed]
135. Fujimura, M.; Ideguchi, M.; Minami, Y.; Watanabe, K.; Tadera, K. Purification, Characterization, and Sequencing of Novel
Antimicrobial Peptides, Tu-AMP 1 and Tu-AMP 2, from Bulbs of Tulip (Tulipa esneriana L.). Biosci. Biotechnol. Biochem. 2004, 68,
571–577. [CrossRef]
136. Giudici, M.; Poveda, J.A.; Molina, M.L.; de la Canal, L.; González-Ros, J.M.; Pfüller, K.; Pfüller, U.; Villalaín, J. Antifungal Effects and Mechanism of Action of Viscotoxin A3. FEBS J. 2006, 273, 72–83. [CrossRef] [PubMed]
137. Ngai, P.H.K.; Ng, T.B. A Napin-like Polypeptide from Dwarf Chinese White Cabbage Seeds with Translation-Inhibitory, TrypsinInhibitory, and Antibacterial Activities. Peptides 2004, 25, 171–176. [CrossRef] [PubMed]
138. Wang, X.; Bunkers, G. Potent Heterologous Antifungal Proteins from Cheeseweed (Malva parviflora). Biochem. Biophys. Res.
Commun. 2001, 279, 669–673. [CrossRef] [PubMed]
139. Pelegrini, P.; Noronha, E.; Muniz, M.A.R.; Vasconcelos, I.; CHIARELLO, M.; Oliveira, J.T.A.; Franco, O. An Antifungal Peptide from Passion Fruit (Passiflora edulis) Seeds with Similarities to 2S Albumin Proteins. Biochim. Et Biophys. Acta 2006, 1764, 1141–1146.
[CrossRef]
140. Agizzio, A.P.; Carvalho, A.O.; Ribeiro, S.D.F.F.; Machado, O.L.T.; Alves, E.W.; Okorokov, L.A.; Samarão, S.S.; Bloch, C.; Prates,
M.V.; Gomes, V.M. A 2S Albumin-Homologous Protein from Passion Fruit Seeds Inhibits the Fungal Growth and Acidification of the Medium by Fusarium oxysporum. Arch. Biochem. Biophys. 2003, 416, 188–195. [CrossRef]
141. Lin, P.; Xia, L.; Wong, J.H.; Ng, T.B.; Ye, X.; Wang, S.; Xiangzhu, S. Lipid Transfer Proteins from Brassica campestris and Mung Bean
Surpass Mung Bean Chitinase in Exploitability. J. Pept. Sci. 2007, 13, 642–648. [CrossRef]
142. Diz, M.; de Oliveira Carvalho, A.; Ribeiro, S.; Cunha, M.; Beltramini, L.; Rodrigues, R.; Nascimento, V.; Machado, O.; Gomes, V.
Characterisation, Immunolocalisation and Antifungal Activity of a Lipid Transfer Protein from Chili Pepper (Capsicum annuum)
Seeds with Novel α-Amylase Inhibitory Properties. Physiol. Plant. 2011, 142, 233–246. [CrossRef] [PubMed]
143. Regente, M.; de la Canal, L. Purification, Characterization and Antifungal Properties of a Lipid-Transfer Protein from Sunflower (Helianthus annuus) Seeds. Physiol. Plant. 2000, 110, 158–163. [CrossRef]
144. Cammue, B.P.; De Bolle, M.F.; Terras, F.R.; Proost, P.; Van Damme, J.; Rees, S.B.; Vanderleyden, J.; Broekaert, W.F. Isolation and Characterization of a Novel Class of Plant Antimicrobial Peptides Form Mirabilis jalapa L. Seeds. J. Biol. Chem. 1992, 267,
2228–2233. [CrossRef]
145. Utkina, L.L.; Andreev, Y.A.; Rogozhin, E.A.; Korostyleva, T.V.; Slavokhotova, A.A.; Oparin, P.B.; Vassilevski, A.A.; Grishin,
E.V.; Egorov, T.A.; Odintsova, T.I. Genes Encoding 4-Cys Antimicrobial Peptides in Wheat Triticum kiharae Dorof. et Migush.:
Multimodular Structural Organization, Instraspecific Variability, Distribution and Role in Defence. FEBS J. 2013, 280, 3594–3608.
[CrossRef]
146. Charnet, P.; Molle, G.; Marion, D.; Rousset, M.; Lullien-Pellerin, V. Puroindolines Form Ion Channels in Biological Membranes.
Biophys. J. 2003, 84, 2416–2426. [CrossRef]
147. Zottich, U.; Da Cunha, M.; Carvalho, A.O.; Dias, G.B.; Casarin, N.; Vasconcelos, I.M.; Gomes, V.M. An Antifungal Peptide from Coffea canephora Seeds with Sequence Homology to Glycine-Rich Proteins Exerts Membrane Permeabilization and Nuclear
Localization in Fungi. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 3509–3516. [CrossRef]
148. Pelegrini, P.B.; Murad, A.M.; Silva, L.P.; dos Santos, R.C.P.; Costa, F.T.; Tagliari, P.D.; Bloch, C., Jr.; Noronha, E.F.; Miller, R.N.G.;
Franco, O.L. Identification of a Novel Storage Glycine-Rich Peptide from Guava (Psidium guajava) Seeds with Activity against
Gram-Negative Bacteria. Peptides 2008, 29, 1271–1279. [CrossRef] [PubMed]
149. López-Meza, J.; Ochoa-Zarzosa, A.; Aguilar, J.; Loeza-Lara, P. Antimicrobial Peptides: Diversity and Perspectives for Their
Biomedical Application. In Biomedical Engineering, Trends, Research and Technologies; IntechOpen: London, UK, 2011; ISBN
978-953-307-514-3.
150. Dash, R.; Bhattacharjya, S. Thanatin: An Emerging Host Defense Antimicrobial Peptide with Multiple Modes of Action. Int. J.
Mol. Sci. 2021, 22, 1522. [CrossRef]
151. Souza, A.L.A.; Díaz-Dellavalle, P.; Cabrera, A.; Larrañaga, P.; Dalla-Rizza, M.; De-Simone, S.G. Antimicrobial Activity of
Pleurocidin Is Retained in Plc-2, a C-Terminal 12-Amino Acid Fragment. Peptides 2013, 45, 78–84. [CrossRef] [PubMed]
152. Thery, T.; Tharappel, J.C.; Kraszewska, J.; Beckett, M.; Bond, U.; Arendt, E.K. Antifungal Activity of a Synthetic Human β-Defensin
3 and Potential Applications in Cereal-Based Products. Innov. Food Sci. Emerg. Technol. 2016, 38, 160–168. [CrossRef]
153. Fernandes, K.E.; Carter, D.A. The Antifungal Activity of Lactoferrin and Its Derived Peptides: Mechanisms of Action and Synergy with Drugs against Fungal Pathogens. Front. Microbiol. 2017, 8, 2. [CrossRef] [PubMed]
154. Muñoz, A.; Marcos, J.F. Activity and Mode of Action against Fungal Phytopathogens of Bovine Lactoferricin-Derived Peptides. J.
Appl. Microbiol. 2007, 101, 1199–1207. [CrossRef]
155. Niaz, B.; Saeed, F.; Ahmed, A.; Imran, M.; Maan, A.A.; Khan, M.K.I.; Tufail, T.; Anjum, F.M.; Hussain, S.; Suleria, H.A.R.
Lactoferrin (LF): A Natural Antimicrobial Protein. Int. J. Food Prop. 2019, 22, 1626–1641. [CrossRef]
156. Silva, P.I.; Daffre, S.; Bulet, P. Isolation and Characterization of Gomesin, an 18-Residue Cysteine-Rich Defense Peptide from the Spider Acanthoscurria gomesiana Hemocytes with Sequence Similarities to Horseshoe Crab Antimicrobial Peptides of the
Tachyplesin Family. J. Biol. Chem. 2000, 275, 33464–33470. [CrossRef]
157. De Lucca, A.J.; Bland, J.M.; Grimm, C.; Jacks, T.J.; Cary, J.W.; Jaynes, J.M.; Cleveland, T.E.; Walsh, T.J. Fungicidal Properties, Sterol
Binding, and Proteolytic Resistance of the Synthetic Peptide D4E1. Can. J. Microbiol. 1998, 44, 514–520. [CrossRef] [PubMed]
158. Zeng, X.-C.; Wang, S.; Nie, Y.; Zhang, L.; Luo, X. Characterization of BmKbpp, a Multifunctional Peptide from the Chinese
Scorpion Mesobuthus Martensii Karsch: Gaining Insight into a New Mechanism for the Functional Diversification of Scorpion
Venom Peptides. Peptides 2012, 33, 44–51. [CrossRef]
159. Zhang, Z.-T.; Zhu, S.-Y. Drosomycin, an Essential Component of Antifungal Defence in Drosophila. Insect. Mol. Biol. 2009, 18,
549–556. [CrossRef] [PubMed]
160. Atanasova-Penichon, V.; Legoahec, L.; Bernillon, S.; Deborde, C.; Maucourt, M.; Verdal-Bonnin, M.-N.; Pinson-Gadais, L.; Ponts,
N.; Moing, A.; Richard-Forget, F. Mycotoxin Biosynthesis and Central Metabolism Are Two Interlinked Pathways in Fusarium graminearum, as Demonstrated by the Extensive Metabolic Changes Induced by Caffeic Acid Exposure. Appl. Environ. Microbiol.
2018, 84. [CrossRef] [PubMed]
161. Lamberty, M.; Zachary, D.; Lanot, R.; Bordereau, C.; Robert, A.; Hoffmann, J.A.; Bulet, P. Insect Immunity. Constitutive Expression of a Cysteine-Rich Antifungal and a Linear Antibacterial Peptide in a Termite Insect. J. Biol. Chem. 2001, 276, 4085–4092. [CrossRef]
[PubMed]
162. Tonk, M.; Cabezas-Cruz, A.; Valdés, J.J.; Rego, R.O.M.; Grubhoffer, L.; Estrada-Peña, A.; Vilcinskas, A.; Kotsyfakis, M.; Rahnamaeian, M. Ixodes ricinus Defensins Attack Distantly-Related Pathogens. Dev. Comp. Immunol. 2015, 53, 358–365. [CrossRef]
163. Moerman, L.; Bosteels, S.; Noppe, W.; Willems, J.; Clynen, E.; Schoofs, L.; Thevissen, K.; Tytgat, J.; Van Eldere, J.; Van Der Walt,
J.; et al. Antibacterial and Antifungal Properties of Alpha-Helical, Cationic Peptides in the Venom of Scorpions from Southern
Africa. Eur. J. Biochem. 2002, 269, 4799–4810. [CrossRef]
164. Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial
Peptides in Marine Invertebrate Health and Disease. Phil. Trans. R. Soc. B 2016, 371, 20150300. [CrossRef] [PubMed]
165. Lamberty, M.; Caille, A.; Landon, C.; Tassin-Moindrot, S.; Hetru, C.; Bulet, P.; Vovelle, F. Solution Structures of the Antifungal
Heliomicin and a Selected Variant with Both Antibacterial and Antifungal Activities. Biochemistry 2001, 40, 11995–12003.
[CrossRef] [PubMed]
166. Zare-Zardini, H.; Taheri-Kafrani, A.; Ordooei, M.; Ebrahimi, L.; Tolueinia, B.; Soleimanizadeh, M. Identification and Biochemical
Characterization of a New Antibacterial and Antifungal Peptide Derived from the Insect Sphodromantis viridis. Biochemistry 2015,
80, 433–440. [CrossRef]
167. Vouldoukis, I.; Shai, Y.; Nicolas, P.; Mor, A. Broad Spectrum Antibiotic Activity of the Skin-PYY. FEBS Lett 1996, 380, 237–240.
[CrossRef]
168. Benincasa, M.; Scocchi, M.; Pacor, S.; Tossi, A.; Nobili, D.; Basaglia, G.; Busetti, M.; Gennaro, R. Fungicidal Activity of Five
Cathelicidin Peptides against Clinically Isolated Yeasts. J. Antimicrob. Chemother. 2006, 58, 950–959. [CrossRef]
169. Lee, D.G.; Kim, H.K.; Kim, S.A.; Park, Y.; Park, S.-C.; Jang, S.-H.; Hahm, K.-S. Fungicidal Effect of Indolicidin and Its Interaction with Phospholipid Membranes. Biochem. Biophys. Res. Commun. 2003, 305, 305–310. [CrossRef]
170. Park, C.H.; Valore, E.V.; Waring, A.J.; Ganz, T. Hepcidin, a Urinary Antimicrobial Peptide Synthesized in the Liver*. J. Biol. Chem.
2001, 276, 7806–7810. [CrossRef]
171. De Lucca, A.J.; Walsh, T.J. Antifungal Peptides: Novel Therapeutic Compounds against Emerging Pathogens. Antimicrob. Agents
Chemother. 1999, 43, 1–11. [CrossRef] [PubMed]
172. Leannec-Rialland, V.; Cabezas-Cruz, A.; Atanasova, V.; Chereau, S.; Ponts, N.; Tonk, M.; Vilcinskas, A.; Ferrer, N.; Valdés, J.J.;
Richard-Forget, F. Tick Defensin γ-Core Reduces Fusarium graminearum Growth and Abrogates Mycotoxins Production with
High Efficiency. Sci. Rep. 2021, 11, 7962. [CrossRef]
173. López-García, B.; González-Candelas, L.; Pérez-Payá, E.; Marcos, J.F. Identification and Characterization of a Hexapeptide with
Activity against Phytopathogenic Fungi That Cause Postharvest Decay in Fruits. Mol. Plant. Microbe Interact. 2000, 13, 837–846.
[CrossRef] [PubMed]
174. López-García, B.; Pérez-Payá, E.; Marcos, J.F. Identification of Novel Hexapeptides Bioactive against Phytopathogenic Fungi through Screening of a Synthetic Peptide Combinatorial Library. Appl Environ. Microbiol. 2002, 68, 2453–2460. [CrossRef]
[PubMed]
175. Muñoz, A.; López-García, B.; Marcos, J.F. Studies on the Mode of Action of the Antifungal Hexapeptide PAF26. Antimicrob. Agents
Chemother. 2006, 50, 3847–3855. [CrossRef]
176. Muñoz, A.; Gandía, M.; Harries, E.; Carmona, L.; Read, N.D.; Marcos, J.F. Understanding the Mechanism of Action of CellPenetrating Antifungal Peptides Using the Rationally Designed Hexapeptide PAF26 as a Model. Fungal Biol. Rev. 2013, 26,
146–155. [CrossRef]
177. Jang, W.S.; Kim, H.K.; Lee, K.Y.; Kim, S.A.; Han, Y.S.; Lee, I.H. Antifungal Activity of Synthetic Peptide Derived from Halocidin,
Antimicrobial Peptide from the Tunicate, Halocynthia aurantium. FEBS Lett. 2006, 580, 1490–1496. [CrossRef] [PubMed]
178. Cary, J.W.; Rajasekaran, K.; Jaynes, J.M.; Cleveland, T.E. Transgenic Expression of a Gene Encoding a Synthetic Antimicrobial
Peptide Results in Inhibition of Fungal Growth in Vitro and in Planta. Plant. Sci. 2000, 154, 171–181. [CrossRef]
179. Fehlbaum, P.; Bulet, P.; Chernysh, S.; Briand, J.P.; Roussel, J.P.; Letellier, L.; Hetru, C.; Hoffmann, J.A. Structure-Activity Analysis of Thanatin, a 21-Residue Inducible Insect Defense Peptide with Sequence Homology to Frog Skin Antimicrobial Peptides. Proc.
Natl. Acad. Sci. USA 1996, 93, 1221–1225. [CrossRef]
180. Osusky, M.; Zhou, G.; Osuska, L.; Hancock, R.E.W.; Kay, W.; Misra, S. Transgenic Plants Expressing Cationic Peptide Chimeras
Exhibit Broad-Spectrum Resistance to Phytopathogens. Nat. Biotechnol. 2000, 18, 1162–1166. [CrossRef] [PubMed]
181. Badosa, E.; Ferre, R.; Francés, J.; Bardají, E.; Feliu, L.; Planas, M.; Montesinos, E. Sporicidal Activity of Synthetic Antifungal
Undecapeptides and Control of Penicillium Rot of Apples. Appl. Environ. Microbiol. 2009, 75, 5563–5569. [CrossRef]
182. Jiang, Z.; Kullberg, B.J.; van der Lee, H.; Vasil, A.I.; Hale, J.D.; Mant, C.T.; Hancock, R.E.W.; Vasil, M.L.; Netea, M.G.; Hodges,
R.S. Effects of Hydrophobicity on the Antifungal Activity of α-Helical Antimicrobial Peptides. Chem. Biol. Drug Des. 2008, 72,
483–495. [CrossRef] [PubMed]
183. Ramamourthy, G.; Na, H.; Seo, C.; Park, Y. Antifungal Activity of (KW)n or (RW)n Peptide against Fusarium solani and Fusarium oxysporum. Int. J. Mol. Sci. 2012, 13, 15042–15053. [CrossRef]
184. Thery, T.; O’Callaghan, Y.; O’Brien, N.; Arendt, E.K. Optimisation of the Antifungal Potency of the Amidated Peptide H-Orn-OrnTrp-Trp-NH2 against Food Contaminants. Int. J. Food Microbiol. 2018, 265, 40–48. [CrossRef] [PubMed]
185. Reed, J.D.; Edwards, D.L.; Gonzalez, C.F. Synthetic Peptide Combinatorial Libraries: A Method for the Identification of Bioactive
Peptides Against Phytopathogenic Fungi. MPMI 1997, 10, 537–549. [CrossRef]
186. Devi, M.S.; Sashidhar, R.B. Antiaflatoxigenic Effects of Selected Antifungal Peptides. Peptides 2019, 115, 15–26. [CrossRef]
[PubMed]
187. Yan, P.-S.; Song, Y.; Sakuno, E.; Nakajima, H.; Nakagawa, H.; Yabe, K. Cyclo(l-Leucyl-l-Prolyl) Produced by Achromobacter
Xylosoxidans Inhibits Aflatoxin Production by Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 7466–7473. [CrossRef]
188. Jermnak, U.; Chinaphuti, A.; Poapolathep, A.; Kawai, R.; Nagasawa, H.; Sakuda, S. Prevention of Aflatoxin Contamination by a
Soil Bacterium of Stenotrophomonas sp. That Produces Aflatoxin Production Inhibitors. Microbiology 2013, 159, 902–912. [CrossRef]
189. Iimura, K.; Furukawa, T.; Yamamoto, T.; Negishi, L.; Suzuki, M.; Sakuda, S. The Mode of Action of Cyclo(l-Ala-l-Pro) in Inhibiting
Aflatoxin Production of Aspergillus flavus. Toxins 2017, 9, 219. [CrossRef]
190. Shehata, M.G.; Badr, A.N.; Sohaimy, S.A.E. Novel Antifungal Bacteriocin from Lactobacillus paracasei KC39 with AntiMycotoxigenic Properties. Biosci. Res. 2018, 15, 4171–4183.
191. Afsharmanesh, H.; Ahmadzadeh, M.; Javan-Nikkhah, M.; Behboudi, K. Improvement in Biocontrol Activity of Bacillus Subtilis
UTB1 against Aspergillus flavus Using Gamma-Irradiation. Crop. Prot. 2014, 60, 83–92. [CrossRef]
192. Farzaneh, M.; Shi, Z.-Q.; Ahmadzadeh, M.; Hu, L.-B.; Ghassempour, A. Inhibition of the Aspergillus flavus Growth and Aflatoxin
B1 Contamination on Pistachio Nut by Fengycin and Surfactin-Producing Bacillus Subtilis UTBSP1. Plant. Pathol. J. 2016, 32,
209–215. [CrossRef] [PubMed]
193. González Pereyra, M.L.; Martínez, M.P.; Petroselli, G.; Erra Balsells, R.; Cavaglieri, L.R. Antifungal and Aflatoxin-Reducing
Activity of Extracellular Compounds Produced by Soil Bacillus Strains with Potential Application in Agriculture. Food Control.
2018, 85, 392–399. [CrossRef]
194. Liu, Y.; Teng, K.; Wang, T.; Dong, E.; Zhang, M.; Tao, Y.; Zhong, J. Antimicrobial Bacillus velezensis HC6: Production of Three
Kinds of Lipopeptides and Biocontrol Potential in Maize. J. Appl. Microbiol. 2019, 128, 242–254. [CrossRef]
195. Veras, F.F.; Correa, A.P.F.; Welke, J.E.; Brandelli, A. Inhibition of Mycotoxin-Producing Fungi by Bacillus Strains Isolated from Fish
Intestines. Int. J. Food Microbiol. 2016, 238, 23–32. [CrossRef] [PubMed]
196. Jiang, C.; Li, Z.; Shi, Y.; Guo, D.; Pang, B.; Chen, X.; Shao, D.; Liu, Y.; Shi, J. Bacillus subtilis Inhibits Aspergillus Carbonarius by
Producing Iturin A, Which Disturbs the Transport, Energy Metabolism, and Osmotic Pressure of Fungal Cells as Revealed by
Transcriptomics Analysis. Int. J. Food Microbiol. 2020, 330, 108783. [CrossRef] [PubMed]
197. Deleu, M.; Paquot, M.; Nylander, T. Fengycin Interaction with Lipid Monolayers at the Air–Aqueous Interface—Implications for the Effect of Fengycin on Biological Membranes. J. Colloid Interface Sci. 2005, 283, 358–365. [CrossRef] [PubMed]
198. Han, Q.; Wu, F.; Wang, X.; Qi, H.; Shi, L.; Ren, A.; Liu, Q.; Zhao, M.; Tang, C. The Bacterial Lipopeptide Iturins Induce
Verticillium dahliae Cell Death by Affecting Fungal Signalling Pathways and Mediate Plant Defence Responses Involved in
Pathogen-Associated Molecular Pattern-Triggered Immunity. Environ. Microbiol. 2015, 17, 1166–1188. [CrossRef]
199. Wang, Y.; Wang, L.; Wu, F.; Liu, F.; Wang, Q.; Zhang, X.; Selvaraj, J.N.; Zhao, Y.; Xing, F.; Yin, W.-B.; et al. A Consensus Ochratoxin
A Biosynthetic Pathway: Insights from the Genome Sequence of Aspergillus ochraceus and a Comparative Genomic Analysis. Appl.
Environ. Microbiol. 2018, 84, e01009-18. [CrossRef]
200. Chen, Y.; Kong, Q.; Liang, Y. Three Newly Identified Peptides from Bacillus megaterium Strongly Inhibit the Growth and Aflatoxin
B1 Production of Aspergillus flavus. Food Control. 2019, 95, 41–49. [CrossRef]
201. Gong, Q.; Zhang, C.; Lu, F.; Zhao, H.; Bie, X.; Lu, Z. Identification of Bacillomycin D from Bacillus Subtilis FmbJ and Its Inhibition
Effects against Aspergillus flavus. Food Control. 2014, 36, 8–14. [CrossRef]
202. Barakat, H.; Spielvogel, A.; Hassan, M.; El-Desouky, A.; El-Mansy, H.; Rath, F.; Meyer, V.; Stahl, U. The Antifungal Protein AFP from Aspergillus giganteus Prevents Secondary Growth of Different Fusarium Species on Barley. Appl. Microbiol. Biotechnol. 2010,
87, 617–624. [CrossRef] [PubMed]
203. Fodil, S.; Delgado, J.; Varvaro, L.; Yaseen, T.; Rodríguez, A. Effect of Potassium Sorbate (E-202) and the Antifungal PgAFP Protein on Aspergillus carbonarius Growth and Ochratoxin A Production in Raisin Simulating Media. J. Sci. Food Agric. 2018, 98, 5785–5794.
[CrossRef] [PubMed]
204. Da Cruz Cabral, L.; Delgado, J.; Patriarca, A.; Rodríguez, A. Differential Response to Synthetic and Natural Antifungals by
Alternaria tenuissima in Wheat Simulating Media: Growth, Mycotoxin Production and Expression of a Gene Related to Cell Wall
Integrity. Int. J. Food Microbiol. 2019, 292, 48–55. [CrossRef]
205. Li, J.; Zhi, Q.-Q.; Zhang, J.; Yuan, X.-Y.; Jia, L.-H.; Wan, Y.-L.; Liu, Q.-Y.; Shi, J.-R.; He, Z.-M. Synthetic Antimicrobial Agents Inhibit
Aflatoxin Production. Braz. J. Microbiol. 2021, 52, 821–835. [CrossRef]
206. Da Cruz Cabral, L.; Rodríguez, A.; Delgado, J.; Patriarca, A. Understanding the Effect of Postharvest Tomato Temperatures on
Two Toxigenic Alternaria spp. Strains: Growth, Mycotoxins and Cell-wall Integrity-related Gene Expression. J. Sci. Food Agric.
2019, 99, 6689–6695. [CrossRef]
207. Delgado, J.; Rodríguez, A.; García, A.; Núñez, F.; Asensio, M.A. Inhibitory Effect of PgAFP and Protective Cultures on Aspergillus parasiticus Growth and Aflatoxins Production on Dry-Fermented Sausage and Cheese. Microorganisms 2018, 6, 69. [CrossRef]
208. Da Cruz Cabral, L.; Rodríguez, A.; Andrade, M.J.; Patriarca, A.; Delgado, J. Effect of Debaryomyces hansenii and the Antifungal
PgAFP Protein on Alternaria spp. Growth, Toxin Production, and RHO1 Gene Expression in a Tomato-Based Medium. Food
Microbiol. 2021, 97, 103741. [CrossRef] [PubMed]
209. Baro, J.I.; Gil-Serna, J.; del Pozo, A.M.; Alvarez, B.P. Analysis of Fusarium graminearum Antifungal Protein and Latrodectin-I
Effect over Growth and Toxigenesis of Aspergillus Fungi with Agrofood Impact. 2021. Available online: https://sciforum.net/ paper/view/9700 (accessed on 2 June 2021). [CrossRef]
210. Mello, E.O.; Ribeiro, S.F.F.; Carvalho, A.O.; Santos, I.S.; Da Cunha, M.; Santa-Catarina, C.; Gomes, V.M. Antifungal Activity of
PvD1 Defensin Involves Plasma Membrane Permeabilization, Inhibition of Medium Acidification, and Induction of ROS in Fungi
Cells. Curr. Microbiol. 2011, 62, 1209–1217. [CrossRef] [PubMed]
211. Jayashree, T.; Subramanyam, C. Oxidative Stress as a Prerequisite for Aflatoxin Production by Aspergillus parasiticus. Free Radic.
Biol. Med. 2000, 29, 981–985. [CrossRef]
212. Merhej, J.; Richard-Forget, F.; Barreau, C. Regulation of Trichothecene Biosynthesis in Fusarium: Recent Advances and New
Insights. Appl. Microbiol. Biotechnol. 2011, 91, 519–528. [CrossRef] [PubMed]
213. Reverberi, M.; Gazzetti, K.; Punelli, F.; Scarpari, M.; Zjalic, S.; Ricelli, A.; Fabbri, A.A.; Fanelli, C. Aoyap1 Regulates OTA Synthesis by Controlling Cell Redox Balance in Aspergillus ochraceus. Appl. Microbiol. Biotechnol. 2012, 95, 1293–1304. [CrossRef]
214. Hoff, B.; Kamerewerd, J.; Sigl, C.; Mitterbauer, R.; Zadra, I.; Kürnsteiner, H.; Kück, U. Two Components of a Velvet-Like Complex
Control Hyphal Morphogenesis, Conidiophore Development, and Penicillin biosynthesis in Penicillium Chrysogenum. Eukaryot
Cell 2010, 9, 1236–1250. [CrossRef]
215. Hegedus, N.; Leiter, E.; Kovács, B.; Tomori, V.; Kwon, N.-J.; Emri, T.; Marx, F.; Batta, G.; Csernoch, L.; Haas, H.; et al. The Small
Molecular Mass Antifungal Protein of Penicillium chrysogenum—A Mechanism of Action Oriented Review. J. Basic Microbiol. 2011,
51, 561–571. [CrossRef]
216. Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between Secondary Metabolism and Fungal Development. Microbiol.
Mol. Biol. Rev. 2002, 66, 447–459. [CrossRef]
217. Meyer, V.; Wedde, M.; Stahl, U. Transcriptional Regulation of the Antifungal Protein in Aspergillus giganteus. Mol. Genet. Genom.
MGG 2002, 266, 747–757. [CrossRef]
218. Manju Devi, S.; Raj, N.; Sashidhar, R.B. Efficacy of Short-Synthetic Antifungal Peptides on Pathogenic Aspergillus flavus. Pestic.
Biochem. Physiol. 2021, 174, 104810. [CrossRef]
219. Bolouri Moghaddam, M.R.; Groß, T.; Becker, A.; Vilcinskas, A.; Rahnamaeian, M. The Selective Antifungal Activity of Drosophila melanogaster Metchnikowin Reflects the Species-Dependent Inhibition of Succinate–Coenzyme Q Reductase. Sci. Rep. 2017,
7, 8192. [CrossRef]
220. Parachin, N.S.; Mulder, K.C.; Viana, A.A.B.; Dias, S.C.; Franco, O.L. Expression Systems for Heterologous Production of
Antimicrobial Peptides. Peptides 2012, 38, 446–456. [CrossRef] [PubMed]
221. Ingham, A.B.; Moore, R.J. Recombinant Production of Antimicrobial Peptides in Heterologous Microbial Systems. Biotechnol.
Appl. Biochem. 2007, 47, 1–9. [CrossRef] [PubMed]
222. Bleackley, M.R.; Dawson, C.S.; Payne, J.A.E.; Harvey, P.J.; Rosengren, K.J.; Quimbar, P.; Garcia-Ceron, D.; Lowe, R.; Bulone, V.; van der Weerden, N.L.; et al. The Interaction with Fungal Cell Wall Polysaccharides Determines the Salt Tolerance of Antifungal
Plant Defensins. The Cell Surface 2019, 5, 100026. [CrossRef]
223. Thery, T.; Arendt, E.K. Antifungal Activity of Synthetic Cowpea Defensin Cp-Thionin II and Its Application in Dough. Food
Microbiol. 2018, 73, 111–121. [CrossRef]
224. Palou, L.; Ali, A.; Fallik, E.; Romanazzi, G. GRAS, Plant- and Animal-Derived Compounds as Alternatives to Conventional
Fungicides for the Control of Postharvest Diseases of Fresh Horticultural Produce. Postharvest Biol. Technol. 2016, 122, 41–52.
[CrossRef]
225. Da Silva Malheiros, P.; Daroit, D.J.; Brandelli, A. Food Applications of Liposome-Encapsulated Antimicrobial Peptides. Trends
Food Sci. Technol. 2010, 21, 284–292. [CrossRef]
226. Luz, C.; Calpe, J.; Saladino, F.; Luciano, F.B.; Fernandez-Franzón, M.; Mañes, J.; Meca, G. Antimicrobial Packaging Based on
ε-polylysine Bioactive Film for the Control of Mycotoxigenic Fungi in Vitro and in Bread. J. Food Process. Preserv. 2018, 42.
[CrossRef]






.jpg&w=3840&q=75)