INTRODUCTION
Goats are an important source of meat and milk (Devendra and Burns, 1983). Recently, goats became an important aspect of animal production in Egypt. Increasing productivity of goats will contribute to improve the standard of living of the rural people. Ruminants have unique ability to utilize the fibrous material through anaerobic fermentation (Kibria et al., 1991), therefore, sunflower seed meal (SFM) can be efficiently used as a sole source of supplemental protein for ruminants (Lardy and Anderson, 2002). Cottonseed cakes (CSC) are being traditionally used as a feed for dairy animals. However, limited supply and seasonal availability of CSC result in high price. On the other hand, SFM is cheaper protein source and can be used as ruminants feed supplement (Yunus et al., 2004). To cover the various nutrients in goats, energy and protein, the feed concentration should be increased, as these dairy animals have a smaller rumen capacity. Oilseeds (sunflower) contribute to increase energy intake of goats and defatted oilseeds in the diet increase the protein intake (Antunac et al., 2001). Traditionally, the farmers have been using cottonseed cakes for feeding their livestock as a source of vegetable protein and its prolonged use can affect the fertility of these animals (Zahid et al., 2003). Earlier investigations (Ahmed et al., 2004) indicated that sunflower meal was equally good in performance, yet the cost of sunflower meal based rations was the lowest. Dietary lipids supplementation, as whole crude oilseeds, may also indeed change fatty acids composition of milk that is one of the most important factors influencing technological and nutritional quality of goat milk (Chilliard et al., 2003). Therefore, the present study was conducted on does to evaluate the effect of replacing concentrate feed mixture containing cotton seed cake by 15 or 20% of whole sunflower seeds on milk production and composition as well as economic efficiency.
MATERIALS AND METHODS
This research is the second part of a serial number of studies on Zaraibi goats (an Egyptian breed) fed a basal ration consisting of 25% concentrate feed mixture (CFM) beside 75% fresh clover (berseem, Trifolium alexandrinum) during winter feeding or 50% CFM and 50% berseem hay (BH) during summer feeding. Concentrate feed mixture consisted of cotton seed cake, linseed cake, yellow corn, wheat bran, molasses, calcium chloride and common salt. Goats of all groups were given the NRC feeding requirements (NRC, 2001) for production of 1-2 kg milk/head/day. The daily feed intake per doe composed of 1.250 kg CFM at 8.00 a.m. + 4 kg green berseem or 1.3 kg berseem hay twice daily (11 a.m. and 3 p.m.). At the late stage of pregnancy (last month of pregnancy, all experimental does (n=30) were divided into three experimental groups according to their age, weight and milk production. Goats in the 1st group (G1) served as a control group and were fed the basal ration (control). However, does in the 2nd (G2) and 3rd (G3) groups were fed the control ration, but 15 and 20% of CFM was replaced by whole sunflower seeds, respectively. The chemical composition and digestibility of the tested rations and their effects on the rumen liquor parameter were given before in the 1stpart (Abdelhamid et al., 2011) of this serial number of studies.
During the suckling period, all does of each experimental group were milked by hand every two weeks. Hand milking was carried out twice at the day of milking (6 a.m. and 5 p.m.). Milk yield was individually measured, recorded and milk samples were taken for chemical analysis. The total milk yield for a doe at the day of milking was considered to represent average daily milk yield during the previous two weeks. During the day of milking, kids were removed from their dams and allowed to suckle other goats. After the end of suckling period, machine milking was applied for all experimental groups up to the end of lactation. Milk yield was individually measured at each milking time using Tru-Test milk meter fixed on the milk line. Milk samples were collected every two weeks by means of such milk meter for chemical analysis. Each doe was dried up when her daily milk yield declined to 200 g for three successive days. The daily milk yield and duration of lactation were individually recorded for goats of each group.
Milk samples (100 ml each) were taken biweekly for chemical analysis. During suckling period, at the day of hand-milking, the morning milk from goats of each breed group cooled at 5°C, added to the evening milk, well mixed and representative samples were taken. During machine milking period, morning cooled milk samples were added to the evening ones, and the representative breed group samples were taken. Milk samples were frozen at -5°C till the time of chemical analysis. Milk samples were analyzed for fat, total protein, lactose and total solids using Milko-Scan (133B N. Foss Electric, Denmark). However, somatic cells count in milk was determined according to standard methodology stated by Marshall et al. (1993) using a Fossomatic cell counter (NS N. Foss Electric,Hillerød,Denmark).
Lipid extraction was conducted according to the methods of A.O.A.C. (2000) using chloroform / methanol (2:1, v/v). The association non-lipids were removed by washing lipid extract three times with CH3OH: H2O (1:1 v/v). The lipids in chloroform were dried over anhydrous sodium sulfate, and then the solvent was removed by heating at 60°C under vacuum. The lipid samples were saponified over night with ethnic KOH (20%) at room temperature. The fatty acids were freed from their potassium salts by acidification with hydrochloric acid (5 N), followed by extraction with ether or (pt.ether 40-60°C). The ether extract was washed three times with distilled water then dried over anhydrous sodium sulfate, and filtered off (Vogel, 1975). Diazomethane was prepped from methylamine hydrochloride as reported by Vogel (1975) as follows: Methalamine solution (100 ml) was placed in a stoppered 500 ml flask and concentrated hydrochloric acid (78 ml) and water were added to bring the total weight to 250 g. Urea (150 g) was introduced and the mixture was boiled gently under reflux for 200 min and vigorously for 15 min., the solution was cooled to room temperature, then sodium nitrite (55 g) were added at 0°C. A mixture of 300 g crushed ice and 50 g concentrated sulfuric acid was prepared in 1500 ml beaker surrounded by a bath of ice and salt. Cold methyl urea- nitrite solution was added slowly with mechanical stirring at such rate, that the temperature did not rise above 0oC. The crystalline nitrosomethyl urea was filtered at once then drained well and dried in vacuum desiccators. Aqueous potassium hydroxide solution (60 ml, 50% w/w) and ether (200 ml) were placed in 500 ml round bottomed flask. The mixture was cooled to 5°C, and then nitrosomethyl urea (20.6 g) and ether (80 ml) were added. The ethereal layer was separated using separating funnel and dried over pellets of potassium hydroxide for 2-3 h.
A set of standard fatty acids of 10:0, 11:0, 12:0, 13:0, 14:0, 15:0, 6:0, 18:0, 18:1, 18:2, 18:3, 20:0, 20:1 and 22:0 carbon atoms with a stated purity of 99% by gas liquid chromatography (GLC) was purchased from Nu-check prop. The purity of each fatty acid methyl ester was checked by GLC and gave one peak. The unsaponifiable phases were also fractionated by GLC on a coild glass column (2.8 m x 4 mm) packed with Diatomite (100-120 mech) and coated with 3% OV-17. The oven temperature was programmed at 10°C/min from 70°C, then isothermally at 270°C for 25 min. and nitrogen flow rate was 30 ml/min. Detector injector temperatures and hydrogen, air flow rates were generally 300°C, 280°C and 33 ml, 330 ml/min, respectively. Peak identification was performed by comparison the retention time of each compound with those of standard materials. The linear relationship between log retention times of the standard hydrocarbons and the number of carbon atoms of these compounds was used to characterize the unavailable authentic hydrocarbons. Peak area was measured by using a computing integrator (PU 4810, Philips).
The histology study was conducted pre- and post-suckling as well as pre-milking during the milking period. Biopsied samples from the middle region of the mammary gland of three does of each group were taken using scalpel with disinfectant surface and about 0.25 g of the tissue was removed into a sterile glass, and after which 5 ml of salt solution + 5% formalin solution were added. A complete dehydration of the mammary tissue samples was accomplished by soaking mammary tissue in 60, 70, 80, 90 and 95% ethanol lasting two hours for each level, followed by two soakings in absolute ethanol (100%) for one hour each. Tissues were then cleared in xylol for not more than two hours, and were transferred to melted paraffin wax (m.p. 52 °C) for three hours in a thermostatically controlled oven. Mould brass was filled with fresh melted wax in which mammary tissues were embedded, oriented carefully and cooled rapidly in cold water. The wax blocks were then stored in refrigerator until sectioning. The mammary tissues were sectioned at 5-7 µm, mounted on glass slides smeared with albumin and glycerol (1: 1). Slides were placed on a hot plate at 45°C to expand the sections and left overnight in an oven at 37° C to dry completely. Sections were immersed in xylol for overnight and then transferred to descending grades of ethanol from 100% until 50%. Ehrlich's haematoxylin and aqueous eosin 1% were used for staining for three minutes and one minute, respectively. Slides were washed in running tap water for ten minutes after haematoxylin and two minutes after eosin. Section were taken up through ascending strengths of 50 to 100% ethanol, then cleared in xylol and mounted in Canada balsam.
Data obtained in this study were subjected to statistical analysis using General Linear Models Procedures (GLMP) adapted by SPSS for windows (2004) for user’s guide. Duncan test of SPSS programme was done to determine the degree of significance between the means at P<0.05.
RESULTS AND DISCUSSION
Milk production:
Milk yield:
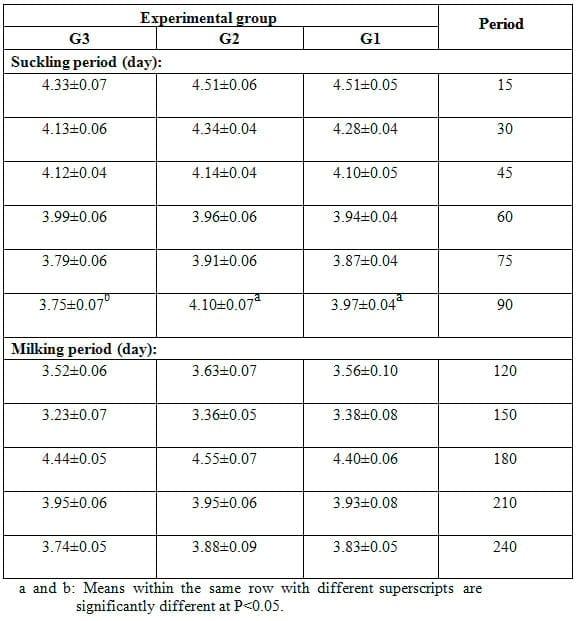
Average daily milk yield during the suckling period (biweekly) and milking period (monthly) as affected by dietary treatment is presented in Table 1. Average daily milk yield during the suckling period showed significant (P<0.05) differences as affected by dietary treatment, being the highest for does fed 20% SFS ration as compared to those in 15% SFS or the control group. The significant differences were observed only at 60, 75 and 90 d of the suckling period. At 90 d of the sucking period and during the whole milking period, average daily milk yield was significantly (P<0.05) higher for does in both treatment groups than that for the control does, but does in 20% SFS group showed significantly (P<0.05) higher yield than those in 15% SFS (Table 1). It is of interest to observe that does in all groups showed similar trend of change in milk yield throughout different lactation weeks, being the highest at 2 wk of lactation and the lowest at the end of lactation period (Table 1). Soliman et al. (1995) found that fat corrected milk yield was increased significantly (P<0.05) between week 1 and 3 of lactation, thereafter fat corrected milk decreased rapidly after three week till the end of lactation period. They suggested that these changes in the milk yield and composition during lactation period may be due to the level of prolactin hormone secretion, efficiency of the udder secretory cells and some other factors. Such trend was also found in Zaraibi goats and several breeds of goats as reported by many authors, with variable timings and levels of the production peak and variable rates of change in milk yield with lactation advance (Hassan et al., 2012). The reviewed studies on the quantitative milk production in lactating dairy goats indicated almost similar trend of changes in daily milk yield. Lactation starts high after kidding, daily increase in milk yield continues and peak is reached within many weeks. Then, the lactation curve declines almost linearly. Rate of decline increases with lactation advance up to the attainment of minimal milk yield values when goats are dried up (Mbayahaga et al.,1994).
Table 1: Effect of feeding the experimental rations on milk yield (mean ± SE) of Zaraibi goats during the suckling and milking periods.
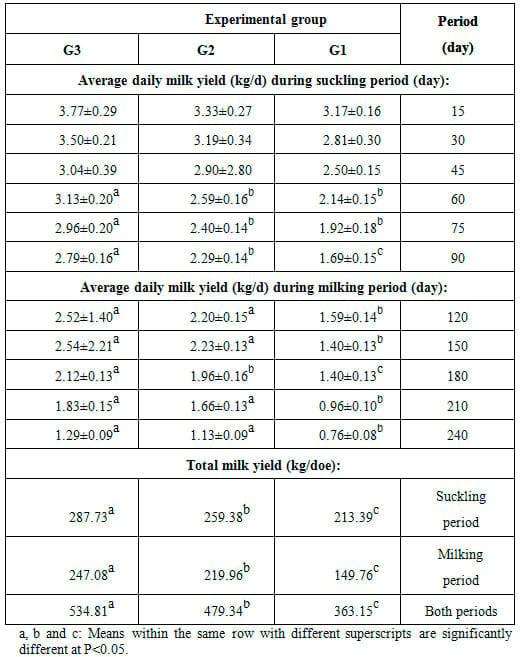
Based on total milk yield, does in 20% SFS group showed significantly (P<0.05) the highest milk yield during the suckling period, milking period or both periods. Does in 15% SFS group ranked the second concerning the corresponding values, but were significantly (P<0.05) lower than those fed 20% SFS. Such findings indicated marked increase in total milk yield of does as level of SFS substitution increased from 15 to 20%. The rate of increase in total milk yield was (21.55% and 10.92%), (46.75% and 12.32%) and (31.99% and 11.57%) in 15 and 20% SFS groups as compared to the control group during the suckling, milking and both periods, respectively (Table 1). Interestingly to observe that the noticed improvement in milk yield of does fed 20% SFS ration was associated with increasing digestibility coefficients of most nutrients and nutritive values of this ration as compared to other rations.
The present results of milk yield are in agreement with those obtained by Giaccone et al. (1995) on Derivata di Siria goats in Sicily. Also, Bekeová et al. (1997) reported the average of milk production as 1.98 kg per test-day in dairy goats in Slovenia. On the contrary, Ciappesoni et al. (2002) observed higher average milk yield (2.58 and 2.93 kg/day) for the white and the brown Czech breeds, respectively. Aguilera et al. (1990) found that average daily milk yield of goats ranged widely from 0.649 to 1.742 kg/d in the first lactation and from 0.222 to 1.989 kg/d in the second lactation. As affected by feeding diets supplemented with sunflower oilseeds in Zaraibi goats, El-Sanafawy (2008) found that daily milk yield for control, 5% sunflower and 10% sunflower groups gradually decreased from 1.83, 2.10 and 2.07 kg/day at the 1st lactation month to 1.43, 1.54 and 1.87kg/day at the end of the 4th month, respectively. Also, Hassan et al. (2012) revealed insignificant differences in milk yield of Zaraibi goats neither between treatment groups fed diets supplemented with calcium soap of fatty acids from soybean, cotton seed or palm oils and control group nor among treatment groups during suckling period or milking period, except for that during the first 30-day post-partum, milk yield of does fed soybean oil diet was significantly (P<0.05) higher than those of control and other treatment groups. Average daily milk yield ranged from 2.098-2.397 kg at 15 d of kidding to 1.090-1.347 kg at weaning (90 days post-kidding) and to 0.845- 0.883 kg at 120 d post-kidding. These results are similar to those reported by Otaru et al., (2011) on goats fed fat supplemented diets.
Milk composition:
Fat content:
Effect of feeding the experimental rations on fat content in milk of Zaraibi goats during the suckling and milking periods is shown in Table 2 which presents that the differences among the experimental groups in fat content of goat milk were not significant during the whole lactation period, except at the end of suckling period (90 d of lactation), fat content was significantly (P<0.05) lower in both treated groups than that in the control one and at 150 d of lactation (milking period), fat content was significantly (P<0.05) higher in does of G2 fed 15% SFS ration than that in those fed 20% SFS ration (G3) and control ration (G1).
Table 2: Effect of feeding the experimental rations on fat content (%, mean ± SE) in milk of Zaraibi goats during the suckling and milking periods.
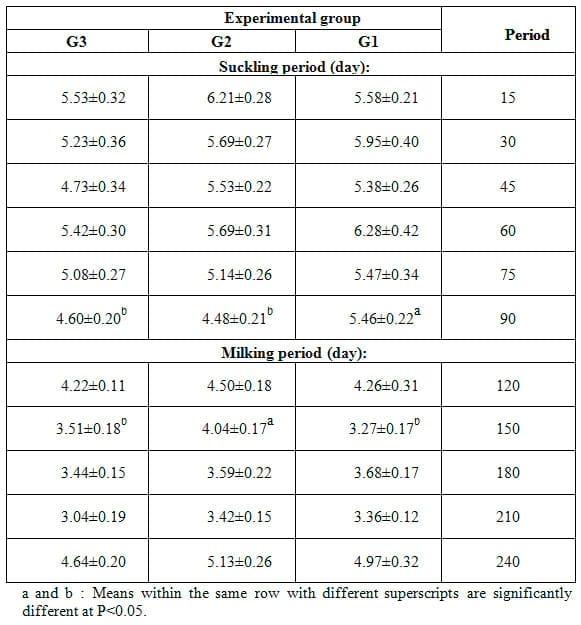
In accordance with the present results, Aguilera et al. (1990) found that milk composition remained rather constant during the mid- stage of lactation (weeks 9-18 of lactation), but the present results indicated slight reduction in fat content of all experimental groups during the same period. Also, Hassan et al. (2012) found that fat percent did not differ significantly by dietary supplementation of different types of plant oils. However, EL-Sanafawy (2008) found that diets contained 5 or 10% SFS increased fat content in milk of Zaraibi goats from 2.99- 4.07% in the control group to 3.51-4.14% in 5% group and 3.20-5.24% in 10% group. In local Barki goats and their crosses with Damascus, Eissa (1996) showed that the fat percentage in milk of Barki does decreased from 5.11% in the first week to 4.24% in the 8th week (suckling period) and increased to 5.67% at the end of milking period. They added that fat percentage in milk of Damascus decreased from 3.86% to 2.99% in the suckling period (the first eight weeks) and increased to 4.72% in the end of milking period. Sauvant et al. (1991) found that milk fat content was high after parturition and then decreased during the major part of lactation in the goat. This is related to a dilution effect due to the increase in milk volume until the lactation peak. Mukherjee et al. (1985) found negative significant correlations between milk yield and fat percentage in the Kuala Lumpur local goats at all stages of lactation. While, negative regression coefficients of fat on total milk yield were found in Jamnapari goats (Pal et al. 1996). In comparing milk fat of Zaraibi goats with others, lower values (1.8-3.7%) were recorded by Mashaly et al. (1984) in the Egyptian Baladi, and higher values (5-5.5%) were reported by Hadjipanayiotou and Photiou (1995) in Damascus goats in Cyprus. However, fat content of 2.8% was reported for Alpine goats in Southern Italy compared with 3.9-5.4% for the native breeds (Pizzillo et al., 1994).
Protein content:
Effect of feeding the experimental rations on protein content in milk of Zaraibi goats during the suckling and milking periods is presented in Table 3 which shows that the differences among the experimental groups in protein content of goat milk were not significant during the suckling period. Similar effect was recorded during the milking period, except at 120 d of lactation (milking period), protein content was significantly (P<0.05) higher in does of G3 fed 20% SFS ration than that in those fed 15% SFS ration (G2) and control ration (G1). Generally, protein content ranged between 2.12 and 3.0% during suckling period and between 2.44 and 3.03% during milking period in all groups. The obtained results are in agreement with those observed in milk of Zaraibi goats fed rations contained 5 or 10% SFS (El-Sanafawy, 2008) or fed different types of oils (Hassan et al., 2012). Also, results available on goats (Chilliard et al., 2003) showed that milk protein content had no marked changes in goats in responses to dietary fat supplementation. Therefore, the present results may indicate that inclusion of SFS in diets of goats had no effects on milk protein. In Zaraibi goats, milk protein content ranged from 2.3-3.2% (El- Gallad et al., 1988) and averaged 3.84% (Hassan et al., 1986).
Table 3: Effect of feeding the experimental rations on protein content (%, mean ± SE) in milk of Zaraibi goats during the suckling and milking periods.
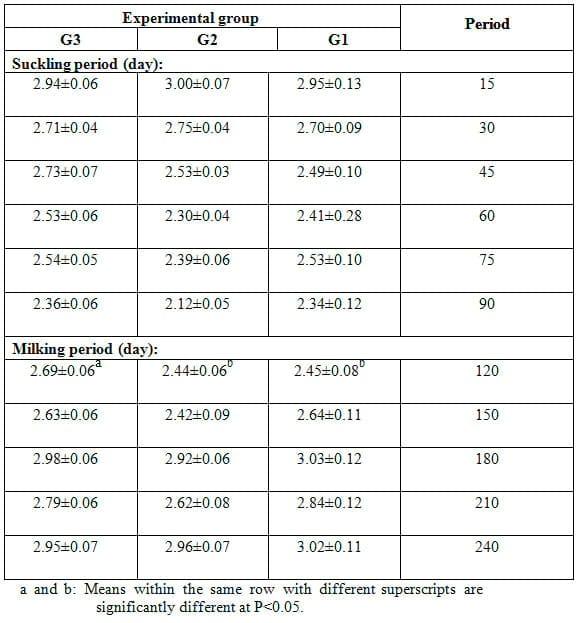
Contrary to the present results, EL-Sanafawy (2008) found marked reduction in protein content of Zaraibi goats with increasing lactation month from the 1st up to the 7th month. In Red Sokoto goats, Ehoche and Buvanendran (1983) reported that milk protein content was high in the first week of lactation, declined rapidly to minimum values between the fourth and sixth week, and then increased gradually up to the end of lactation with lower variations from week to week. In Damascus, local Barki goats and their crosses, Eissa (1996) found that milk protein content decreased from the first week until the fifth to sixth week, then increased till week 16. In Jamnapari goats, significant differences in casein content in milk were found between lactation stages (Pal et al., 1996). In Murciano-Granadina goats, Fernandez et al. (2004) reported that protein content was 3.54, 3.60, 3.45, 3.53 and 3.60 at 1st, 2nd, 3rd, 4th and 5th weeks of lactation, respectively,
Lactose content:
Effect of feeding the experimental rations on lactose content in milk of Zaraibi goats during the suckling and milking periods is shown in Table 4 which shows that the differences among the experimental groups in lactose content of goat milk were not significant during the suckling period, except at 90 d, lactose content was significantly (P<0.05) lower in does of G3 fed 20% SFS ration (G3) than that in those fed 15% SFS ration (G2) and control ration (G1). During the milking period, lactose content was not affected significantly by dietary treatment at all times of milking periods. Generally, lactose content ranged between 3.75 and 4.51% during suckling period and between 3.23 and 4.55% during milking period in all groups. As affected by lactation period, lactose content showed inconsistent trend of changes throughout the lactation period. However, the present findings are in agreement with Antunac et al. (2001), who observed higher content of lactose at the beginning of lactation than at middle of lactation period. Similar results were reported by El-Sanafawy (2008) and Zamfirescu (2009). However, Francisco et al. (2002) found that milk lactose decreased (P<0.05) at start of lactation period to reach lowest value at week 5, thereafter it tended to increase gradually until the end of lactation period.
Table 4: Effect of feeding the experimental rations on lactose content (%, mean ± SE) in milk of Zaraibi goats during the suckling and milking periods.
Total solids content:
Effect of feeding the experimental rations on total solids content in milk of Zaraibi goats during the suckling and milking periods is presented in Table 5 which shows significant (P<0.05) differences among the experimental groups in total solids (TS) content of goat milk only during the suckling period. Within the 1st month of suckling period, only inclusion of SFS at a level of 20% in ration of goats significantly (P<0.05) decreased TS content in milk as compared to the control. However, at the last 2 weeks of suckling period, inclusion of both SFS levels significantly (P<0.05) decreased TS content as compared to the control ration. On the other hand, TS content was not affected significantly by dietary treatment at all times of milking periods. Generally, TS content showed marked reduction in all groups in particular within the 1st intervals of suckling period.
Table 5: Effect of feeding the experimental rations on total solids content (%, mean ± SE) in milk of Zaraibi goats during the suckling and milking periods.
The general trend of change in the mean total solids of goat’s milk during lactation period is in full agreement with those reported by Pal et al. (1996). Wide variations in TS content were observed in different breeds of goats, being 10.7% in Alpine (Lu, 1993), 14.9% in Beetal goats (Verma and Chawla, 1984). However, no clear differences in total solids were found between Damascus and Barki goats and their crosses (Eissa, 1996).
Fatty acids composition in milk fat:
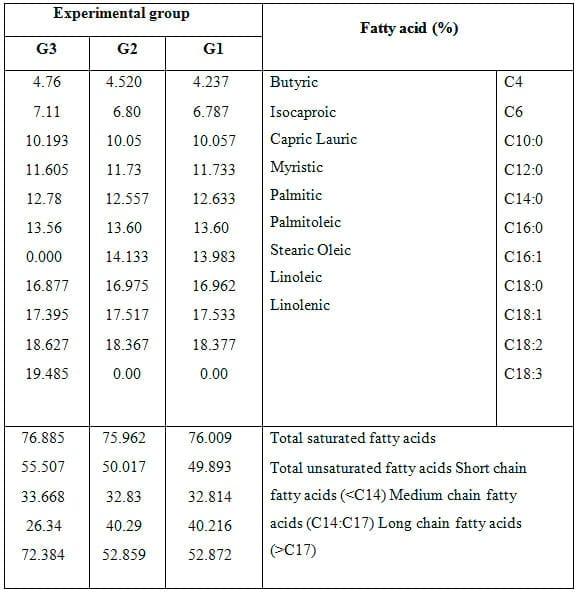
Effect of feeding experimental diets on separation condition of fatty acids (%) of milk is shown in Table 6 where the dietary treatment affected fatty acids composition in milk fat of goats. The pronounced differences in type of fatty acids among milk of different groups were in palmitoleic and linolenic fatty acids. Milk of goats fed 20% SFS ration was characterized by absence of palmitoleic acid and presence of linolenic acid. Also, feeding goats on 20% SFS ration increased total content of unsaturated fatty acids as well as long chain fatty acids in milk fat. Lipid composition is one of the most important components of the technological and nutritional quality of goat milk. Lipids are involved in cheese yield (per kilogram of milk) and firmness, as well as in the color and flavor of caprine dairy products (Delacroix-Buchet and Lamberet, 2000). Besides their quantitative contribution to the amount of dietary energy, the different fatty acids (short- and medium-chain, saturated, branched, mono- and polyunsaturated, cis and trans, conjugated) are potentially involved as positive or negative predisposing factors for the health of human consumers (Williams, 2000).
Furthermore, the peculiarities of goat milk lipolytic system (Chilliard, 1982) and medium-chain fatty acids (Ha and Lindsay, 1993) could greatly change the content in free fatty acids, playing a major role in the occurrence of the characteristic goat flavor. The response of milk fatty acid composition is nearly similar, in particular for major fatty acids, including conjugated linoleic acid (CLA) in milk of goats in all groups. Chilliard et al. (2003) observed that for goats fed diets supplemented with either protected or unprotected lipids, milk CLA content increases sharply after either vegetable oil supplementation or fresh grass feeding, but does not change markedly when goats receive whole untreated oilseeds. Important interactions are observed between the nature of forages and of oil supplements on trans- 10 and trans-11 C18:1 and CLA.
Griinari et al. (1998) reported that changes in the non-glucogenic ratio of VFA will depress acetate production and de novo synthesis of short chain fatty acids in the mammary gland and direct inhibition of milk fat synthesis can result from the production of partially hydrogenated fatty acids.Sauvant et al. (1991) reported that the reduction in fat mobilization decreased the availability of plasma NEFA, especially C18:0 and C18:1, for mammary lipid synthesis. These fatty acids did not differ in response to dietary treatment. Furthermore, many authors stated that the response to feeding different kinds of unprotected lipid supplements consisted mainly of an increase in the percentages of milk C18:0 and C18:1, at the expense of mainly C8 to C14 and C16:0 (Schmidely and Sauvant, 2001). This was probably due to the ruminal hydrogenation of polyunsaturated fatty acids into C18:0 and trans-C18:1, which are inhibitors of the de novo FA synthesis, mainly C8 to C16. The final response of milk C16:0 percentages depended on the level of dietary intake, that is, the C16:0 percentages in the lipid supplement which was studied. Therefore, the present similarity in fatty acid composition and slight increase in total unsaturated fatty acids in milk of goats fed 20% SFS ration may indicate the safe use of SFS up to 20% without adversely effects on hydrogenation of fatty acids in rumen and consequently marked changes in fatty acid composition in milk fat. Changes accompanied by C16:1 and C18:3 are mainly due to their proportions in SFS. A similar conclusion was reported by Bernard et al. (2005). However, saturated fatty acids of C16 and C18 had little effect on milk fatty acid profile (Harvatine and Allen 2006). In the present study, the higher proportions of C18:3 in milk of does fed 20% SFS ration are responsible for the lack of differences in C18:0 and C18:1. Chilliard and Ferly (2004) indicate that C18:3 is often hydrogenated into C18:0, while hydrogenation of C18:2 gives rise to different isomers of C18:1.
Milk somatic cells count (SCC x103):
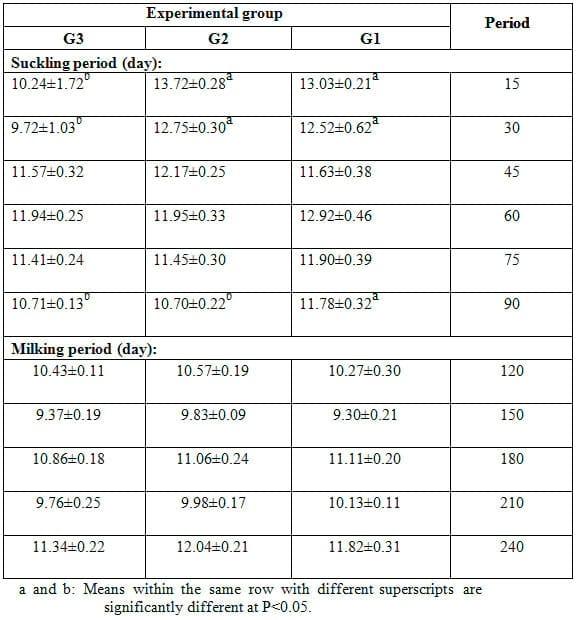
Means of somatic cells count (SSC) during different lactation stages (suckling and milking periods) in the milk of different groups are shown in Table 7. Results indicated that milk SCC was significantly (P<0.05) lower in milk of does fed 15% SFS ration (G2) than that of those fed 20% SFS ration (G3) and control one (G1) during different times of the suckling and milking periods. However, the differences in SSC between G3 and G1 were not significant, except at 15 and 120 days. Throughout the lactation period, SSC showed inconsistent trend of change in all experimental groups. It is of interest to note that values of SSC presented in this study for all groups are higher than that reported in milk of goats. This may reflect a higher incidence of infection; however, information on incidence of mastitis in this study was not available.
Table 6: Effect of feeding experimental diets on separation condition of fatty acid (%) of milk.
In this concern, Lerondelle and Poutrel (1984) determined SSC using a Coulter counter. They indicated that the SCC in uninfected halves of goat mammary glands ranged from 0.470 to 2.7 x106 cells/ml. Intramammary infection in one udder half of goats increased SCC in the corresponding uninfected half (Dulin et al., 1983), which could be an important factor in milk quality testing.
In accordance with the present results, Cerón-Muñoz et al. (2002) found that SCC decreased in the second month of lactation and increased thereafter up to the ninth month of lactation. Also, Farghaly (2002) showed that stage of lactation significantly affected milk SSC, since both the original and logarithm milk somatic cells were the highest shortly after calving, dropped to a minimum between 40 and 80 days postpartum and then steadily increased until the end of lactation. In addition, the changes in SSC in milk of goats may be associated with seasonal variation.
Table 7: Effect of feeding experimental diets on somatic cells count (x 103, mean ± SE) of milk during the suckling and milking period.
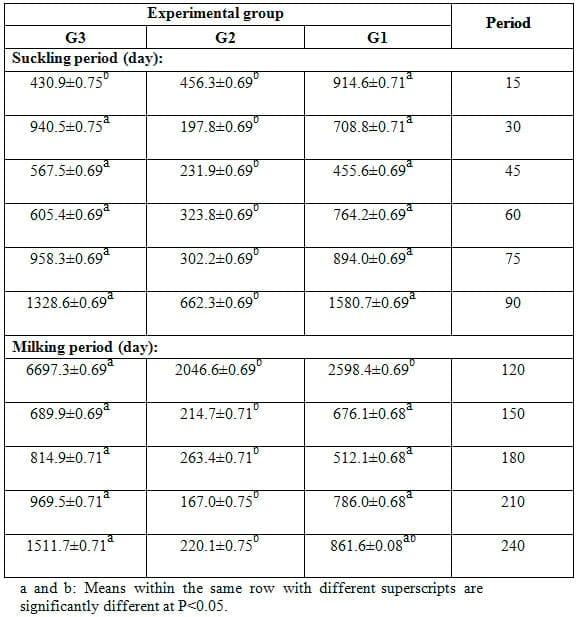
Hinckley (1991) observed the lowest SCC in milk of goats in April and the highest values in September-October. However, in another study with goat milk bulk tank samples, SCC did not appear to be affected by seasonal variation (Tirard-Collet et al., 1991). Seasonal variation may reflect the number of fresh to mid and late lactation does because goats breed seasonally. Similar results were obtained by Rodriguez et al. (2000), who reported that season had significant effect on milk somatic cells count. Zhang et al. (1994) found the highest SCC occurred during summer season. Dakic et al. (2006) stated that stress from winter cold and summer hot period had considerable influence on somatic cells count to increase in cow milk and therefore it causes considerable milk quality decrease. Furthermore, SSC in milk during winter season was higher than limit levels stated by Djabri et al. (2002) and Hamann (2002). The reasons for seasonal variations are unknown and only speculated to be the effects of housing, rain and temperature changes on infection status. Based on the foregoing results of milk production, increasing level of dietary inclusion of SFS to 20% showed pronounced improvement in milk yield as average daily milk yield during the suckling and milking periods. Both levels of SFS inclusion had no marked effect on approximate chemical composition and composition of fatty acids in milk fat. However, 15% SFS level showed considerable reduction in SCC in milk of goats.
Histological changes in mammary gland:
The histological examination of specimens of the mammary gland of Zaraibi goats showed normal architecture of the mammary gland in all experimental groups; however, some differences were observed as affected by dietary treatment or lactation period in terms of number and size of milk alveoli, number and volume of secretory cells, size as well as area of the mammary stroma (connective tissues).
During the suckling period:
During the suckling period, the histological examination pre- or post-milking revealed normality in the histological structure of the mammary gland of different experimental groups as affected by dietary treatment with marked differences, among experimental groups and even between pre- and post-milking within the same group, in volume, density, thickness and number of epithelium, and lumen diameter of milk alveoli as well as thickness of connective tissues (adipose tissues or stroma) as illustrated in Plates 1 and 2.
Plate (1): Sections in pre-suckling mammary gland of goats in different experimental groups.
Plate (2): Sections in post-suckling mammary gland of goats in different experimental groups
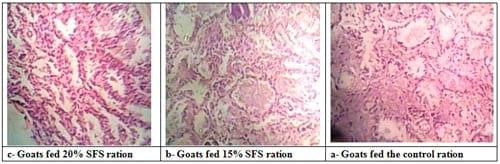
Pre-suckling, as a results of presence of milk in alveolar system within the alveolar lumen, milk alveoli were seen to be bigger, more compacted with wider lumen, and thinner and numerous epithelial cells in mammary gland of does fed 15 and 20% SFS than in the control does. Also, thickness of connective tissues and consequently the amount of adipose tissues or stroma was less in the mammary gland of does fed 15 and 20% SFS than in the control does (Plate 1). Such findings are in accordance with milk yield of goats in different experimental groups. During lactation, milk is synthesized within the alveolar compartment and its amount depends on the activity and number of the mammary epithelial cells (MEC). Cells number in the mammary gland is the function of the rates of cells proliferation and cells death. The mammary gland grows when the rate of proliferation exceeds the rate of death, and it regresses when the rate of death exceeds the rate of cells proliferation (Capuco et al., 2001). In addition, a close correlation between milk yield and the amount of secretory tissue has been reported in ruminants (Baldi et al., 2002). Nutrition affects mammary development during pregnancy and lactation in ruminants. During early lactation, mammary proliferation is proportionally greater in species that exhibit less negative energy balance (Stefanon et al., 2002). Deficiencies in the dietary supply of specific amino acids (histidine, methionine and lysine) in early and late lactation, markedly reduced milk yield with no clear evidence of corresponding changes in measurements of mammary cells number, activity or proliferation rate (Yeo et al., 2003). After suckling, the histological examination of the mammary gland revealed that milk alveoli in all experimental groups were smaller, denser with narrow lumen, and thicker epithelium than that observed pre-suckling. Moreover, thickness of connective tissues and consequently the amount of adipose tissues or stroma was higher in the mammary gland of does in all groups post- than pre-suckling. However, thickness of epithelium was the highest in the mammary gland of does fed 20% SFS (Plate 1). These observations are in association with removal of milk in alveolar system of goats in different experimental groups.
During the milking period:
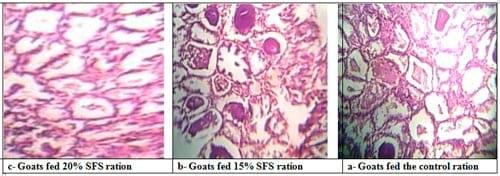
During the milking period, the histological examination pre-milking revealed also normality in the histological structure of the mammary gland of different experimental groups as affected by dietary treatment with marked differences between does fed 20% SFS diet and those fed 15% SFS and the control diets. Generally , the mammary gland of does fed 20% SFS diet showed milk alveoli with higher volume, density, thickness, number of epithelium, and lumen diameter of milk alveoli than those observed in the mammary glands of does fed 15% SFS and control diets (Plate 3).
Plate (3): Sections in pre-milking mammary gland of goats in different experimental groups.
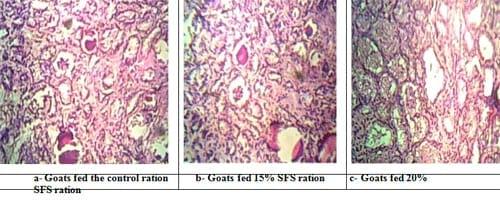
Since the activity of mammary cells did not decrease as lactation advanced in non pregnant animals, the main reason for milk decline during late lactation appear to be reduced MEC number. However, during late lactation, when animals were concomitantly lactating and pregnant, the secretory activity per cell also declines due to conflicting metabolic demands of gestation and lactation (Capuco et al., 2003). In accordance with the differences in the histological structure of the mammary gland of goats between suckling (early lactation period) and milking period (late lactation period), Capuco et al. (2001) observed that the increase in milk yield until lactation peak (from wk 3 to wk 8 in goats, and from wk 2 to wk 13 in cows) appeared to be due to increased synthetic capacity of the MEC (hypertrophy), rather than to an increase in number of secretory cells. The progressive decline in milk yield after peaking during suckling period was associated with a decrease in the total DNA content of the mammary parenchyma, representing a net fall in cells number (Capuco et al., 2001). Mammary gland in goats can grow during early lactation. This growth during early lactation is important; for instance, gestational mammary development can be reduced experimentally in mice with no deleterious effects on subsequent milk production (Knight and Wilde, 1987). In the same manner, this may occur in goats. However, during the late lactation (involution) the connective tissue increased concomitantly with the regression of the alveoli into the smallest size and reduced frequency of secretory cells (Plate 3). At the late lactation (suckling), the DNA frequency was lower than during milking (Plates 1 and 3), this case is in agreement with Capuco et al. (2001) and Hassan (2004), who reported, in cattle and buffalo, respectively, that the content of parenchymal DNA was the greatest at day 14 of lactation and declined to a low level at day 240. Also in sheep, the high content of DNA at the early stage of lactation indicated a high correlation between mammary cells number and milk yield. The highest amount of DNA at mid lactation indicates the greatest number of secretory cells with the high activity of lactation.
Economic efficiency of milk production:
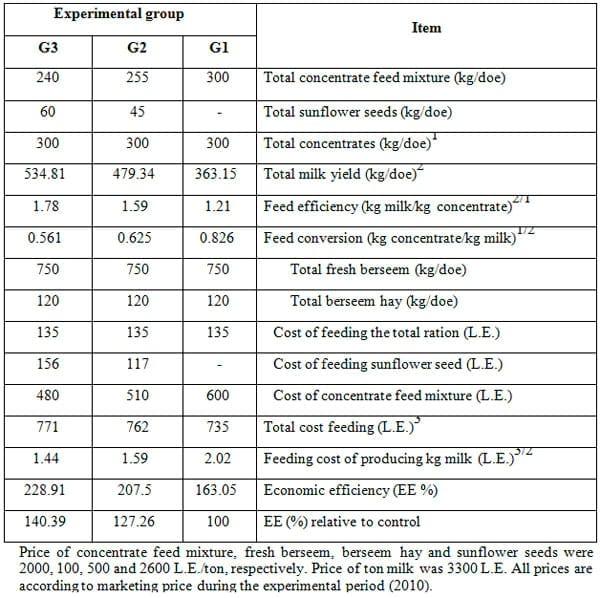
Data of economic evaluation of feeding the experimental rations on milk production during the whole lactation period (suckling and milking periods) are presented in Table 8. The present results revealed that milk / feed efficiency and feed conversion for concentrates was the best for does fed 20% SFS ration, followed by those fed 15% SFS ration, while those fed the control ration showed the poorest values. Cost of each kg milk was the lowest (1.44 L.E.) for does fed 20% SFS ration, the modest (1.59 L.E.) for those fed 15% SFS ration and the highest (2.02 L.E.) for the control does.
Such trend was reflected in the highest economic efficiency of milk production in does fed 20% SFS ration, followed by those fed 15% SFS and control rations (299.1, 207.5 and 150%, respectively. This was mainly attributed to feed efficiency and feed conversion to milk in each experimental group. Similar trends were obtained by El-Sanafawy (2008) on goats fed 5 and 10% SFS rations, which may indicate that increasing replacement of concentrate feed mixture containing cotton seed cake by sunflower seeds from 5% up to 10% increased economic efficiency of milk production.
Table 8: Economic efficiency of milk production in different experimental groups during the lactation period.
REFERENCES
Abdelhamid, A. M.; A. E. Abdel-Khalek, T. M. Ashmawy, F. M. Abou Ammou and H. A. El-Sanafawy (2011). Effect of dietary inclusion of whole sunflower seeds on feeding lactating Zaraibi goats:I.on digestibility coefficients, rumen function and live body weight digestibility coefficients, rumen function and live body weight. J. Animal and Poultry Production,MansouraUniversity, 2: 535-547
Aguilera, J. F.; C. Prieto and J. Fonolla (1990). Protein and energy metabolism of lactating Granadina goats. British Journal of Nutrition, 63: 165-175.
Ahmed,I.; K. Javed, K; R. H. Mirza; A. Sattar and F. Ahmad (2004). Effect of feeding sunflower meal as a substitute of cottonseed cake on growth and age of maturity in Holstein Friesian heifers.PakistanVet. J., 24 (2): 95-97.
Antunac, N. ; D. Samarsija ; J. L. Havranek ; V. Pavic and B. Mioc (2001). Effect of stage and number of lactation on chemical composition of goat milk. Czech.J.Animal Sci., 46 (12): 548-553.
AOAC (2000). Official methods of analysis, 14th ed. Association of Official Analytical Chemists.WashingtonDC.
Baldi, E.; M. Luconi; L. Bonaccorsi and G. Forti (2002). Signal transduction pathways in human spermatozoa. Journal of Reproductive Immunology, 53:121–131.
Bekeová E. ; J. Elecko; V. Hendrichovský ; J. Choma and; M. Krajnicakova (1987). The effect of beta-carotene on the changes in T4 and cholesterol concentrations in calving heifers before and after parturition. Vet. Med. (Praha), 32: 459 – 468.
Bernard, L. ; J. Rouel ; C. Leroux ; A. Ferlay ; Y. Faulconnier ; P. Legrand and Y. Chilliard (2005). Mammary lipid metabolism and milk fatty acid secretion in alpine goats fed vegetable lipids. J. Dairy Sci., 88:1478–1489.
Capuco, A. V.; D. L. Wood; R. Baldwin; K. Mcleod and M. J. Paape (2001). Mammary cell number, proliferation, and apoptosis during a bovine lactation. J. Dairy Sci., 84: 2177-2187.
Capuco, A. V. ; S. E. Ellis ; S. A. Hale ; E. Long ; R. A. Erdman ; X. Zhao and M. J. Paape (2003). Lactation persistency: Insights from mammary cell profileration studies. J. Anim. Sci., 81: (3): 18-31.
Cerón-Muñoz, M. ; H. Tonhati ; J. Duarte ; J. Oliveira ; M. Muñoz- Berrocal and H. Jurado-Gàmez (2002). Factors affecting somatic cell counts and their relations with milk and milk constituent yield in buffaloes. J. Dairy Sci., 85:2885-2889.
Chilliard, Y. (1982).Variations physiologiques des activites lipasique et de la lipolyse spontanee dans les laits de vache, de chevre et de femme: revue bibliographique. Lait 62:1–31 and 126–154.
Chilliard, Y. and A. Ferlay (2004). Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev., 44: 467–492.
Chilliard Y. ; A. Ferlay ; J. Rouel and G. A. Lamberet (2003.). Review of nutritional and physiological factorsaffecting goat milk lipid synthesis and lipolysis. J Dairy Sci., 86:1751–1770.
Ciappesoni C. G. ; M. Milerski and J. Pribyl (2002). Parámetros productivos de cabras de la raza checa marrón de pelo corto. In: XXVII Jornadas ientíficas y VI Jornadas Internacionales SEOC, 19–21 de septiembre Valencia (España): 819–825.
Dakic, A.; N. Pintic; F. Poljak; A. Novosel; D. Strucic; T. Jelen and V. Pintic (2006). Utjecaj godisnjeg doba na broj somatskih stanica u kravljem mlijeku isporucenom za trziste. Stocarstvo, 60: 35.
Delacroix-Buchet, A. and G. Lamberet (2000). Sensorial properties and typicity of goat dairy products. 7th Int. Conf. on Goats,Tours,France. 15–21 May 2000. Tome, 2:559–563.
Devendra, C. and M. Burns (1983). Goat Production in the Tropics. 2nd ed., CAB.,Edinburgh.
Djabri, B. ; N. Bareille, F. Beaudeau and H. Seegers (2002). Quarter milk somatic cell count in infected dairy cows: A meta-analysis. Vet. Res., 33: 335–357.
Dulin, A. M. ; M. J. Paape ; W. D. Schultze and B. T. Weinland (1983). Effect of parity, stage of lactation and intramammary infection on concentration of somatic cells and cytoplasmic particles in goat milk. J. Dairy Sci., 66:2426-2433.
Ehoche, O. W. and V. Buvanendran (1983). The yield and composition of milk and preweaning growth rate of Red Sokoto goats inNigeria. World Rev. Anim. Prod., 19(2):19-24.
Eissa, M. M. (1996). Improving productivity of desert Barki goats by crossing withDamascusgoats with special reference to milk production. Ph.D. Thesis, Fac. Agric., Alex. Univ.
El-Gallad, T. T. ; E. A. Gihad ; S. M. Allam and T. M. El-Badawy (1988). Effect of energy intake and roughage ratio on the lactation of Eyptian Nubian (Zaraibi) goats. J. Small Ruminant Res., 1:327-341.
El-Sanafawy, H. A. (2008). Performance of goats fed on rations containing whole sunflower seeds. M. Sc. Thesis, Fac. Agric., Kafer El-Sheikh Univ.
Farghaly, H. M. (2002). Comparison among different mathematical models of lactation curves to describe the somatic cell counts in Fleckvieh milk. Zagazig J. Agric. Res., 29: 211.
Fernandez, J. R. ; M. Rodriguez Osorio ; M. Ramos ; E. de la Torre ; G. Gil Extremera and M. R. Sanz Sampelayo (2004). Effect of rumen protected supplements of fish oil on intake, digestibility and nitrogen balance of growing goats. Anim. Sci., 79: 483–491.
Francisco, C. C. ; C. S. Chamberlian ; D. N. Waldner ; R. P. Wettemann and L. J. Spicer (2002). Propionibacteria fed to dairy cows: Effects on energy balance, plasma metabolites and hormones and reproduction. J. Dairy Sci., 85: 1738.
Giaccone, P. ; B. Portolano; A. Bonanno; M. L. Alicata and M. Todaro (1995). Aspetti quanti-qualitativi della produzione lattea nella popolazione caprina Derivata di Siria. Zoot. Nutr. Anim., 21: 97-109.
Griinari, J. M. ; D. A. Dwyer ; M. A. McGuire ; D.E. Bauman; D. L. Palmquist and K. V. V. Nurmela (1998). Trans-octadeconic acids and milk fat depression in lactating dairy cows. J. Dairy Sci., 81: 1251.
Ha, J. K. and R. C. Lindsay. (1993). Release of volatile branched-chain and other fatty acids from ruminant milk fats by various lipases. J. Dairy Sci., 76:677–690.
Hadjipanayiotou, M. and A. Photiou (1995). Effects of protein source and level on performance of lactatingDamascusgoats in negative energy balance. Small Ruminant Res.,15:257-263.
Hamann, J. (2002). Milk quality and udder health in relation to modern milking. In: Recent developments and perspectives in bovine medicine. 22nd World Buiatrics Congress, Hannover;Germany, pp: 334-345.
Harvatine, K. J. and M. S. Allen (2006). Effects of fatty acid supplements on ruminal and total tract nutrient digestion in lactating dairy cows. J. Dairy Sci., 89:1092-1103.
Hassan Laila R. (2004). Histochemical assessment of mammary gland capacity pertaining to milk production in buffaloes. Egyptian J. Anim. Prod., 41 (Suppl. Issue, Nov.): 309-320.
Hassan, G. A. ; F. D. El-Nouty ; M. A. Samak and M. H. Salem (1986). Relationship between milk production and some blood constituents in Egyptian Baladi goats. Beitrage zür Tropischen Landwirtschaft und Veterinarmedizin., 24(2): 213-219.
Hassan, T. M. ; M. Ibrahim, M. ; K. Itman andI.Abdel-Hai (2012). Productive and reproductive performance of Zaraibi goats fed different types of protected fat. The 13th Scientific Conference for Animal Nutrition, Sharm El-Sheikh, 14-17 Feb.(under publishing).
Hinckley, L. S. (1991). Proposed somatic cell count revision.United Caprine News, June, 36.
Kibria, S. S.; M. R. Islam and C. K. Shah (1991). Development of straw based ration for feeding ruminants. Asian-Aust. J. Anim. Sci., 4: 235-240.
Knight, C. H. and C J. Wilde (1987) Mammary growth during lactation: implications for increasing milk yield. J. Dairy Sci., 70: 1991- 2000.
Lardy, G. P. and V. Anderson (2002). Canola and sunflower meal in beef cattle diets. Vet. Clinics of North America: Food Animal Practice, 18: 327-338.
Lerondelle, C. and B. Poutrel (1984). Characteristics of non-clinical mammary infections of goats. Annales de Recherches Vétérinaires, 15:105-112.
Lu, C. D. (1993). Implication of feeding isoenergetic diets containing animals fat on milk composition of Alpine does during early lactation. J. Dairy Sci., 76:1137-1147.
Marshall, R.T. ; J. E. Edmonson and B. Steevens (1993). Using thecaliforniamastitis test.UniversityofMissouriExtension. October 1993. Available at http:// extension.missouri. Edu / explore / agguides / dairy / g03653.htm.
Mashaly, R.I.; S. A. El-Deeb ; F. D. El-Nouty ; G. A. Hassan and M. H. Salem (1984). Changes in milk yield and in milk chemical and physical properties during lactation period in Egyptian Baladi goats. Egyptian J. Dairy Sci., 12(2):123-134.
Mbayahaga, J. ; S. N. M. Mandiki; J. L. Bister; R. Paquay; L. Bangirinama and R. Branckaert (1994). Production and composition of the milk of localBurundigoats and growth of suckled kids. Revue d’Elevage et de Medecine Veterinaire des Pays Tropicaux, 47 (4): 405-410.
Mukherjee, T. K.; A. R. Samudram and S. Sivaraj (1985). Milk composition of local and F1 (local female X improved German Fawn male) goats. Malaysian Applied Biology, 14(2): 100-103.
NRC (2001). Nutrient Requirements of Dairy Goats. 7th Rev. ed. National Academy Press, Washington, DC.
Pal, U. K.; V. K. Saxena; M. K. Agnihotri and R. Roy (1996). Effect of season, parity and stage of lactation on the composition of Jamnapari goat’s milk. International J. Anim. Sci., 11(1): 245- 248.
Pizzillo, M. ; E. Cogliandrio; R. Rubino and V. Fedele (1994). Productivity and qualitative characteristics of milk from the principal goat breeds reared inSouthern Italy. “Progressi scientifici e tecnologici in tema di patologia e di allevamento degli ovini e dei cvaprini”. Societa Italiana di Potologia e di Allevamento degli Ovini e dei Caprini. Atti XI Congresso Nazionale, Perugia, Italy, 1-4 June 1994. Perugia, Italy ; Facolta di Agraria, Universita degli Studi di Perugia 431-434 (ABAs, 64:3724).
Rodriguez, S. D. ; M. A. Garcia Ortiz ; G. Hernandez Salgado; N. A. Santos Cerda ; R. Aboytes Torre and G. J. Canto Alarcon (2000). Anaplasmamarginale inactivated vaccine: dose titration against a homologous challenge. Comp Immunol. Microbiol. Infect. Dis., 23: 239-252.
Sauvant, D.; Y. Chilliard and P. Morand-Fehr (1991). Etiological aspects of nutritional and metabolic disorders of goats in Goat Nutrition. P. Morand-Fehr, ed. Pudoc, Wageningen (NLD), EAAP Publ. No. 46:124–142.
Schmidely, P. and D. Sauvant (2001). Fat content yield and composition of milk in small ruminants: effects of concentrate level and addition of fat. INRA-Productions-Animals, 14(5): 337.
Soliman, A. M.; M. H. KHalil; S. M. Zahed and E. A. Afifi (1995). Effectiveness and comparison of the three methords of cow evaluation to improve milk traits of Fleckvieh cattle.Egypt. J. Anim. Prod., 32: 151.
SPSS for windows (2004). Statistical Package for Social Science (SPSS). Release 13.0. The Apasche software foundation for windows.
Stefanon, B.; M. Colitti; G. Gabai; C. H. Knight and C. J. Wilde (2002). Mammary apoptosis and lactation persistency in dairy animals. J. Dairy Res., 69:37-52.
Tirard-Collet, P.; J. A. Zee; L. Carmichael and R. Simard (1991). A study of the microbiological quality of goat milk inQuebec. J. Food Prod., 54: 263-266.
Verma, N. K. and D.S. Chawla(1984). Variation in milk composition in dairy goats. Indian J. Anim. Sci., 54(6):539-543.
Vogel, A. J. (1975). A Textbook of Practical Original Chemistry.3rd ed. P: 969-971, English Language Book Society and Longman Group Ltd.London.
Williams, C. M. (2000). Dietary fatty acids and Human health. Ann.
Zootech., 49: 165-180.
Yeo, J. M.; C. H. Knight and D. G. Chamberlain (2003). Effects of changes in dietary amino acid balance on milk yield and mammary function in dairy cows. J. Dairy Sci., 86 (4): 1436- 1444.
Yunus, A. W.; A. G. Khan; Z. Alam; J. I. Sultan and M. Riaz (2004). Effect of substituting cottonseed meal with sunflower meal in rations for growing buffalo calves. Asian- Aust. J. Anim. Sci., 17: 659-662.
Zahid,I.A.; L. A. Lohdi; N. Ahmed; N. U. Rehman and M. S. Akhtar (2003). Effects of cotton seed cake (gossypol) on live weight of teddy male goats. Pakistan Vet. J., 23: 27-30.
Zamfirescu, S. (2009). NoutaNi în cresterea caprelor, Ed. EX PONTO, ConstanNa.
Zhang, W. C.; J. C. M. Dekkers; G. Banos and E. B. Burnside (1994). Adjustment factors and genetic evaluation of somatic cell score and relationships with other traits of Canadian Holsteins. J. Dairy Sci., 77: 659–665.







.jpg&w=3840&q=75)