vitamin E and selenium in anestrous
Role of vitamin E and selenium in anestrous and conception
Published: March 16, 2010
By: Dr Rajendran Duraisamy (National Institute of Animal Nutrition and Physiology)
Role of vitamin e and selenium in anestrous and conception
"Fertility is a luxurious event
For this, endometrium should be protected by antioxidant
Antioxidant can be given as nutrient
Vitamin E and Selenium can be given for this event
Let fed this nutrients in this moment "
With this quote let me start as competent
•1. Introduction
|
Anoestrus is one of the major causes of economic losses in both the large and small ruminants in India. It is a major problem in the tropics and subtropics, where inadequate nutrition, high ambient temperature, high parasite burdens and disease exacerbate the problem. Low body weight and poor body condition, compounded with lactation stress, can further extend the postpartum anoestrous period. Vandeplassche (1982). The causes of functional infertility included cystic and inactive ovaries with anoestrus, early embryonic mortality with repeat breeding, and prolonged gestation. Anoestrus often reflects a hormonal disturbance and accounted for 47.8% of the cases. Repeat breeding, where cows require three or more services to conceive, accounted for 11.5% of cases. Singh et al (1981) also found functional infertility to be more common than infertility due to infectious diseases (76 vs 24%). There is a very important interrelationship between nutrition and production performance of dairy cows and this interrelationship has far reaching effects on the physiological functioning of the reproductive system which is constantly under the influence of the endocrine system.
Anoestrus is a period of sexual quietude in which the animal fails to exhibit normal oestrus cycles and no manifestation of heat (Boyd, 1977). Its effects are greater than most dairy farmers realize. Not only does it lengthen the postpartum interval (a period from parturition to oestrus) but also substantially reduce the farmer''''s financial returns from milk or calf sales due to overall reduced number of pregnancies (calving), and the cost of managing prolonged postpartum anoestrus. It is essential to distinguish physiological from or pathological anoestrus. Cows are regarded as physiologically anoestrous before puberty, during pregnancy and for a few days (up to 60) following parturition, whereas, lack of oestrus at 60 days postpartum is termed pathological anoestrus (Lamming,1980) The potential problems that breeders may encounter include cows that gave birth to twin calves had retained placenta, have uterine infection or milk fever. Extensive efforts, world-wide, have been put into research to limit the occurrence of this disorder (anoestrus). However despite all these efforts postpartum infertility is still a significant problem in dairy herds. Nonetheless few advances have been made in reducing the postpartum interval through proper nutrition including supplementation of antioxidant namely vitamin E, Selenium and carotenoids.
A key factor influencing the productivity per cow is conception rate. The conception rate observed in Tamilnadu was just 34.53 per cent in cows and 25.52 per cent in buffaloes |
The main goal in a commercial dairy cattle operation is to optimize number of calf produced per cow as economically as possible. A key factor influencing the productivity per cow is conception rate, both at first service and during the remainder of the breeding season. The conception rate observed in Tamilnadu (Table 1) was just 34.53 per cent in cows and 25.52 per cent in buffaloes (Thirunavukkarasu, 2006). No data available regarding conception rate in cattle and buffaloes in India.
Nutrition, before and after calving, has an effect on pregnancy rates. Inadequate nutrition prior to calving, results in cows being thin at calving which delays the onset of estrual activity post-calving. Cows on an inadequate plane of nutrition after calving can easily delay the first service and overall conception rates reduced by 5-10 percent and even more dramatic effects, such as a 20-40 percent reduction in conception rates have been noted in research trials
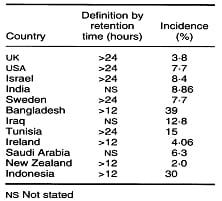
Table 1. Published estimates of the incidence of retained placenta indifferent countries. Laven and Peters (1996)
Soils in many of the important dairy regions of the world are Se-deficient. Dairy cattle consuming stored forages are often low in vitamin E unless supplemented., and vitamin E deficiencies are frequently observed during the periparturient period, leading to retain placenta and subsequently Anoestrus |
Calving Difficulty, It include retain placenta, a number of studies have shown that increased calving difficulty will not only increase the length of time required for the cows to cycle after calving, but decrease the percent of cows conceiving in the first post-partum estrus and often reduce overall conception rate during the breeding season. Two to 30% of cows retain their foetal membranes for 12 to 24 hours after a normal delivery. The afterbirth, or foetal membranes, is retained if the cotyledonary villi fail to detach from the caruncular crypts. Membranes retained for more than 2 or 3 days decompose in the uterus, leading to metritis. A number of reproductive diseases can have a major impact on the overall reproductive performance of a cow herd.
The two conditions mentioned above is related with few aspects of stress, immunity and health. Antioxidant can have the role to reduce factors affecting estrous cycle and conception rate.
Soils in many of the important dairy regions of the world are Se-deficient, and feedstuffs grown on these soils provide inadequate dietary Se. Dairy cattle consuming stored forages are often low in vitamin E unless supplemented, and vitamin E deficiencies are frequently observed during the periparturient period leading to retain placenta and subsequently anoestrus. Supplementation of vitamin E and Se is best result of improve overall health, increases conception rate and reduces the incidence of anestrous. The main aim of this essay is to explore the newer data regarding the role of vitamin E and selenium on anestrous and conception.
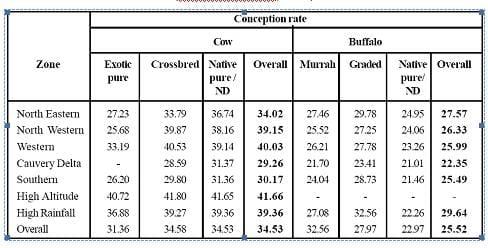
Table 2. Conception rates observed in different agro-climatic zones of Tamilnadu (Thirunavukkarasu, 2006)
•2. HEALTH VS. ANOESTRUS AND CONCEPTION RATE
Often one of the key factors reducing conception rates in a cow herd is health problems. Subclinical uterine infection is major cause of infertility in crossbred cattle and buffaloes in India. Subclinical uterine infection can be reduced or prevented by the increasing animal immune status either by vaccination or by increasing general immunity. There is evidence that deficiencies of E and Se can lead to impaired resistance to disease. A number of reproductive diseases can have a major impact on the overall reproductive performance of a cow herd casing anoestrus and reduce conception rate.
Many species of bacteria inhabit the vagina, uterus, and cervix of cows. Some are symbionts that become pathogenic when the animal is stressed; others are immediately pathogenic.
Namboothripad and Raja (1976), Eduvie et al (1984) and El-Azab et al (1988) isolated Staphylococcus aureus, Escherichia coli, Pseudomonas pyocyanea, Corynebacterium pyogenes, Proteus mirabilis, Streptococcus spp., Pasteurella multocida, Proteus vulgaris, Klebsiella spp. and several anaerobic microorganisms from the uteri of cows with a history of repeat breeding, retained placenta and metritis, as well as from the uteri of normal suckling cows. Mycobacterium tuberculosis was isolated by Mohanty et al (1980) from a Haryana heifer that was a chronic repeat breeder.
Subclinical uterine infection is major cause of infertility in crossbred cattle and buffaloes in India. There is evidence that deficiencies of E and Se can lead to impaired resistance to disease. |
Listeria monocytogenes may also cause abortion in cattle. When the organism infects a pregnant cow, it invades the foetal nervous system and forms necrotic foci on the liver, lungs and spleen (Watson, 1979), killing the foetus. Vandeplassche (1982) indicated that, although the organisms are easily eliminated from the uterus, they may persist in the mammary system. Antibodies to Listeria are short-lived, and immunity is thus only temporary and cows can be re-infected. Treatment is often futile, even with antibiotics. However if the animal are sound nutritional background and good immunity, the rate of infection is less.
Harrison and others (1984) found that supplementing cows with selenium significantly decreased the incidence of cystic ovarian disease from 47 to 19 per cent, but that the vitamin E status of the cows had no effect on the incidence ofcystic ovarian disease. In contrast, Jaskowski (1993) found that vitamin E supplementation significantly increased the beneficial effect of selenium supplementation on ovarian cysts. Erskine and others (1997) reported that 3000 iu of vitamin E given eight to 14 days before parturition significantly decreased the incidence of metritis, probably as a side effect of the reduction in retained fetal membranes which is usually associated with a significantly increased risk of uterine infection. However, Harrison and others (1984) found that supplementing cows with vitamin E in addition to selenium had no further beneficial effect on the incidence of metritis, despite a significant reduction in the incidence of retained fetal membranes.
•3. IMMUNE STATUS VS. ANOESTRUS AND CONCEPTION RATE
The immune system can be partitioned into two broad categories; specific (or acquired) and nonspecific. Specific or acquired immunity is the basis of vaccination programs. Specific immunity occurs when animals develop or acquire immunity to a specific pathogen once it is exposed to the pathogen. Antibodies specific to that pathogen are produced and the immune system "memorizes" the antigenic properties of the pathogen so that an immune response can be initiated quickly when the host is exposed to the pathogen again. Lymphocytes and macrophages are the cells primarily involved with specific immunity. The nonspecific immune system is designed to protect the body from all antigens. Vaccination will not influence nonspecific immunity and the nonspecific immune system does not have a "memory." Neutrophils are the cells most involved with nonspecific immunity.
When a pathogen invades the uterus or reproductive tract of a cow, a cascade of events occurs. First, neutrophils from the blood are drawn to the infection site. Neutrophils are the first line of defense after a pathogen invades the body. The function of neutrophil is to engulf (phagocytize) and then kill bacteria. After a neutrophil engulfs a bacterium, and chemical reaction called a respiratory burst occurs and this produces a high concentration of free radicals. These free radicals help kill the bacteria, but if they are not controlled, they also can damage and kill the neutrophil. The life span of neutrophils is short; each neutrophil can engulf 5 to 20 bacteria before the cell is killed. As part of the inflammatory response, macrophages also are drawn into the infection site. These cells can kill bacteria directly but more importantly they initiate the acquired immune response. Antibodies are produced against the bacteria and lymphocytes are drawn to the infection site. The combined efforts of neutrophils, macrophages, lymphocytes, and antibodies help to eliminate the invading pathogen and reduce the disease condition of reproductive tract and increase the occurrence of estrus and there by increases conception rate.Vitamin E and se containing glutathione peroxidase have sparing effects on the requirements for one another relative to intracellular killing of bacteria. The protection afforded cellular membranes by vitamin E may spare the requirement for glutathione peroxidase by reducing free radicals at the membrane, thereby preventing leakage of free radicals into the cytosol and maintaining intracellular killing capacity of the cell. Conversely, glutathione peroxidase activity in the cytosol may spare the requirement for vitamin E in the membranes.
Feeding approximately 1000 IU/day of supplemental vitamin E with adequate selenium - reduced the prevalence of retained fetal membranes 3000 iu of vitamin E given eight to 14 days before parturition significantly decreased the incidence of metritis |
Clinical studies have been conducted to evaluate the effect of supplemental vitamin E on prevalence of retained fetal membranes, intramammary infections, and clinical mastitis. Feeding approximately 1000 IU/day of supplemental vitamin E (usually all-ractocopheryl acetate) to dry cows when adequate selenium was supplemented reduced the prevalence of retained fetal membranes in some (Harrison et al., 1984; Miller et al., 1993). When vitamin E was injected (usually in combination with selenium) rather than fed, about half the time there was no effect for prevalence of retained fetal membranes and about half the time there was a positive response (Miller et al., 1993).
Two fairly common diseases, vibriosis and trichomoniasis, will cause lower conception rates. Cows will breed, but then return to heat fairly soon afterwards. Two other diseases that can impact conception rates are red-nose (IBR) and bovine viral diarrhea (BVD). A number of diseases will cause abortion to occur and leave the animals infertile. This would include diseases such as leptospirosis, hemopholaus somnus and brucellosis. Presence of immunity against these diseases reduces the incidence of anestrous and increases the conception rate. Feeding of vitamin E and Selenium increases non specific immunity and can reduce the occurrence of bacterial and viral diseases there by increase estrus cycle and conception rate.
3.1IMMUNE RESPONSEON VITAMIN E AND SELENIUMSUPPLEMENTATION
Several studies have shown that vitamin E supplementation has a protective effect. Reddy et al. (1987) fed 32 Holstein calves up to 500 iu per day of supplementary vitamin E. They found that supplementation increased the blastogenic responses of both T cells and B cells, and that the anamnestic antibody response was highest in calves supplemented with 125 iu vitamin E per day. Hidiroglou et al. (1992) reported that the injection of 2700 iu of vitamin E every three weeks from birth for 12 weeks significantly increased the production of immunoglobulin (Ig) M; the production of IgG, and IgG2 was also increased, but not significantly. Hogan et al. (1992) reported that injecting 3000 iu of vitamin E subcutaneously, 10 and five days before expected calving, significantly increased the intracellular kill of bacteria by neutrophils at calving. However, neither the phagocytic index nor the percentage of neutrophils phagocytosing bacteria differed between the vitamin E-injected and placebo-injected cows. Politis and others (1995) supplemented eight dairy cows with 3000 iu of vitamin E per cow per day orally, beginning four weeks before and continuing until eight weeks after parturition, and with 5000 iu intramuscularly one week before the expected date of parturition. Although they found no evidence for an effect of the vitamin E supplementation on mammary macrophages, it significantly enhanced the chemotactic responsiveness of blood neutrophils from two to four weeks after parturition. Not all studies have observed a significant response of the immune system to vitamin E supplementation. Cipriano et al. (1982) found that the oral supplementation with 1000 iu of vitamin E of calves fed on skimmed milk had no significant effects on their IgG concentrations and lymphocyte stimulation indices. The lack of significance may have been because too few animals were used, because the differences they reported were large. Mudron et al. (1994) investigated the effect of vitamin E on leucocyte parameters in transported calves. The oral administration of 20 iu of vitamin E 24 hours before they were loaded had no significant effect on leucocyte migration and serum immunoglobulin concentration. Plasma cortisol concentration was reduced for 24 hours after the administration of vitamin E, but this reduction had no effect on the calves'''' cortisol response to the stress of transport. The mechanism by which vitamin E supplementation has immune stimulatory effects is not fully understood. One effect may be the reduction in corticosteroids observed by Reddy et al. (1987), because glucocorticoids are immunosuppressive compounds and inhibit lymphocyte proliferation directly. Another effect may be due to a change in arachidonic acid metabolism. Vitamin E supplementation has been shown to reduce the production of prostaglandin F2cx and prostaglandin E2 in chicks (Lawrence et al. 1985). Under conditions of stress, changes in the levels of these compounds can adversely affect the function of immune cells, particularly the activity and responsiveness of lymphocytes (Hadden 1987). It is most likely, however, that the protective effect of vitamin E is due to stabilisation of leucocyte membranes. Leucocyte membranes are more vulnerable to peroxidative damage than the membranes of body cells because they contain more free fatty acids (Kigoshi and Ito 1973). On the basis of serum enzyme levels, Reddy et al. (1987) reported that the cell membranes of unsupplemented calves were more prone to damage.
•4 OXIDATIVE DAMAGE OF ENDOMETRIUM ON ANOESTRUS AND CONCEPTION RATE
During parturition and after heavy milk yield causes stress to the animal. In stress condition, normal cell processes, environmental insults, and inflammatory responses produce compounds called reactive oxygen species or free radicals. Environmental insults include solar radiation, heavy milk yield , certain mycotoxins, nitrates, and a host of other toxic compounds. When a pathogen invades the uterus or reproductive tract of a cow, a cascade of events occurs. Respiratory burst occurs and this produces a high concentration of free radicals. The major free radicals found in biological systems are superoxide, hydrogen peroxide, hydroxyl radical, and fatty acid radicals. Hydrogen peroxide is found primarily in the cytosol of cells and fatty acid radicals are found primarily in cell membranes. Superoxide and hydroxyl radicals can be found in both cell components. Because free radicals are extremely toxic to cells, the body has developed a sophisticated antioxidant system (Table 1). Superoxide dismutase (an enzyme that contains copper and zinc) converts superoxide to hydrogen peroxide. Hydrogen peroxide is converted to water by the enzyme Glutathione peroxidase (GSHpx). Those two enzymes effectively control most free radicals within the cytosol. Superoxide and the hydroxyl radical can migrate into cell membrane where they attack fatty acids (especially unsaturated fatty acids) and produce fatty acid radicals (a process called initiation). Fatty acid radicals then react with other fatty acids producing a chain reaction. Vitamin E and to a lesser extent -carotene reacts with fatty acid radicals and stops the chain reaction.
Free radicals are highly reactive compounds because they are missing an electron. Free radicals can react with nucleic acids causing mutations, they can react with enzymes and render them inactive, and they can react with fatty acids in membranes causing membrane instability. Free radicals can eventually kill cells and damage tissues.
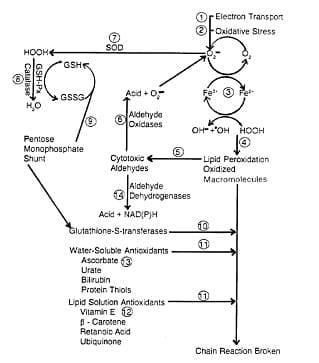
Figure 1. Systems for protection against reactive oxygen metabolites. (Miller et al., 1993)
1) Superoxide is generated during normal metabolism. 2) Exogenous contributors to oxidative stress include dietary imbalances, disease, environmental pollutants, and solar radiation 3) Superoxide reduces Fe3+, enabling it to enter into Fenton-type reactions. which produce hydroxyl radical. 4) The extremely reactive hydroxyl radical attacks macromolecules and initiates peroxidative chain reactions (22). 5) Cytotoxic aldehydes are end products of lipid peroxidation. 6) When tissues are disrupted, aldehyde dehydrogenases are converted to aldehyde oxidases, which generate superoxids. 7) Superoxide dismutases (Mn, Cu, and Zn) convert superoxide to peroxides. This conversion retards reduction of Fe3+ to Fez+, which catalyzes formation of .OH 8) Catalase Q and glutathione peroxidase (Se)convert peroxides to compounds that do not participate in Fenton-type reactions . Reduction of peroxides is accompanied by oxidation of reduced glutathione. 9) Reduced glutathione can be regenerated from glutathione disulfide (GSSG) by reducing equivalents from NADPH, which is generated by the pentose monophosphate shunt . 10) Glutathione S-transferases conjugate glutathione with peroxy radicals. This pathway may be more active when it is deficient in Se or vitamin E. The resulting destruction of glutathione increases consumption of reducing equivalents, thus competing with other metabolic pathways that depended on NADPH. 11) Chain-breaking antioxidants interrupt peroxicative chains initiated by reactive oxygen metabolites that escaped enzymatic degradation. 12) Vitamin E serves as a chain-breaking antioxidant by reacting directly with free radicals . Although vitamin E is consumed when free radicals are quenched . Reducing equivalents are conserved in comparison with glutathione S-transferases serving as chain breakers. 13) Vitamin C, in addition to regenerating vitamin E and possibly also glutathione, can act in its own ri&t as a water-soluble antioxidant. 14) Aldehyde dehydrogenases convert aldehydes to less toxic products.
Production of free radicals could represent a source of infertility because ovarian steroidogenic tissue (Carlson et al.,1993; Maas, et al.,1993), spermatozoa (Aitken, 1994) and preimplantation embryos are sensitive to free radical damage. In some studies, administration of beta carotene (Ascarelli, et al., 1985; Bindas, et al., 1984) or vitamin E and selenium (Arechiga, et al.1994; Segerson, et al. 1977) improved fertility of cattle.
Harrison et al. (1984) suggested that vitamin E and selenium act at the cellular level by regulating the generation of free radicals in the ovaries. Staats and others (1988) showed that vitamin E protected steroidogenic enzymes from oxidative degeneration, and Rapoport et al. (1998) found that the concentration of a-tocopherol in ovarian tissue was related to the animals'''' consumption of vitamin E during the period of maximal progesterone production. Other work has suggested that the generation of free radicals is a potential cause of abnormal embryonic development (Goto et al., 1992), and Barnes and Smith (1975) suggested that vitamin E promoted the release of follicle stimulating hormone (FSH), adrenocorticotrophic hormone (ACTH) and luteinising hormone (LH). There may also be a role for vitamin E in the protection of the pathway from arachidonic acid to prostaglandins, which are closely involved in the regulation of the reproductive system.
4.1 OXIDATIVE STRESS, ANTIOXIDANTS AND THE PERIPARTURIENT DAIRY COW
The periparturient period is especially important for health of dairy cattle. A survey including 551 cows and 1305 lactations (Shanks et al., 1981)revealed that over one-half of total health costs resulting from mammary and reproductive problems occurred during the first 30 DIM. In addition to cost of treatment, udder edema (Dentine.
et al., 1983), retained placenta (Joosten et al., 1988), and mastitis (Smith et al., 1984) can reduce milk production, market value, and the productive life of the cow and can cause indirect costs that are difficult to quantify. Delayed first estrus, delayed first breeding, and repeated breeding resulting from failure to conceive or from early embryonic death increase days open and prolong calving intervals. Additional expenses include treatment, repeated breeding, and culling of cows for failure to conceive.
Table 3. Antioxidant functions of vitamin E and of mammalian cells.
Component (location in cell) | Nutrients Involved | Function |
Alpha-tocopherol (membranes) | Vitamin E | Breaks fatty acid peroxidation chain reactions |
Glutathione peroxidase (cytosol) | Selenium | An enzyme that converts hydrogen peroxide to water |
•5 ROLE OF VITAMIN E AND SELENIUM
5.1 VITAMIN E ACT AS ANANTIOXIDANT
Vitamin E is the most important lipid-soluble antioxidant and the biologically most active form is d,a-tocopherol (Putnam and Comben, 1987). Vitamin E is an integral component of all lipid membranes and serves to protecting lipid membranes from attack by ROS (Rice and Kennedy, 1988). Polyunsaturated fatty acids ( PUFA) of membranes are particularly vulnerable to attack by ROS, and ROS can initiate a chain reaction of lipid destruction that destroys the membrane of the cell. Vitamin E can quench peroxidation reactions in membranes and is probably the most important antioxidant located in cell membranes (Putnam and Comben, 1987).
Vitamin E and glutathione peroxidase function at two locations within the cell Glutathione peroxidase functions in the cytosol of the cell and vitamin E within lipid membranes. An important function of both systems is the protection of membrane PUFA. The PUFA are present in all cellular membranes, but their concentration varies considerably from tissue to tissue Membrane PUFA are extremely susceptible to attack from reactive oxygen species, and the higher the concentration of membrane PUFA, the more susceptible the cell and tissue to oxidant damage.
An important PUFA in cellular membranes is arachidonic acid ( AA) . Arachidonic acid can be metabolized to prostaglandins, thromboxanes, and prostacyclin by the enzyme complex cyclooxygenase and to the leukotrienes by the lipoxygenase enzyme complex. vitamin E may function to control peroxidation of AA or its unstable metabolites. The AA metabolites are important for PMN function and the amplification of the inflammatory response following pathogen invasion of tissues including the uterus, which is more prone for infection.
5.2 SELENIUM ACT AS AN ANTIOXIDANT
Beneficial health effects of selenium supplementation in dairy cows (Rayman, 2000; Malbe et al., 1995). It is generally understood that these benefits are achieved through the action of selenium as a component of selenoproteins.
The GSH-Px and thioredoxin reductase groups of selenoproteins catalyze reduction of peroxides that can damage cells and tissues. Being a part of these proteins, selenium has the capacity to affect oxidative processes in the system and is considered an antioxidant nutrient. These antioxidants protect cells from oxidative damage from free radicals and peroxides (Rotruck et al., 1973). The activity of these enzymes is directly related to the concentration of selenium in the diet. Furthermore, several studies have demonstrated that oxidative stress of the cow contributes to lowering immunity and increases the risk of retained fetal membranes as well as mastitis (Hemmingway, 1999) which increases post partum incidence of anestrous and lower conception rate. Higher activity of GSH-Px has also been reported in the erythrocytes of cows receiving supplemental selenium compared with non-supplemented individuals (Ortman and Pehrson, 1999). Selenium therefore has a positive antioxidant effect in cattle.
Table 4 Interpretation of selenium analyses in cattle

5.3 ANTI-PATHOGENIC ACTIVITIES OF SELENIUM AND VITAMIN E
Subclinical uterine infection is major cause of infertility in crossbred cattle and buffaloes in India. Subclinical uterine infection can be reduced or prevented by the increasing animal immune status either by vaccination or by increasing general immunity. There is evidence that deficiencies of E and Se can lead to impaired resistance to disease Vitamin E supplementation of diets increased intracellular kill of S. aureus and E. coli by bovine neutrophils but had no effect on phagocytic index (Hogan et al., 1990). The effects of vitamin E and Se supplementation on intracellular kill of bacteria by neutrophils were not additive. Supplementation with both vitamin E and Se did not result in greater intracellular kill of bacteria by neutrophils than did supplementation with either one of the nutrients alone. However, vitamin E and glutathione peroxidase have sparing effects on the requirements for one another relative to intracellular killing of bacteria. The protection afforded cellular membranes by vitamin E may spare the requirement for glutathione peroxidase by reducing free radicals at the membrane, thereby preventing leakage of free radicals into the cytosol and maintaining intracellular killing capacity of the cell. Conversely, glutathione peroxidase activity in the cytosol may spare the requirement for vitamin E in the membranes.
Clinical studies have been conducted to evaluate the effect of supplemental vitamin E on prevalence of retained fetal membranes, intramammary infections, and clinical mastitis. Feeding approximately 1000 IU/day of supplemental vitamin E (usually all-ractocopheryl acetate) to dry cows when adequate selenium was supplemented reduced the prevalence of retained fetal membranes in some (Harrison et al., 1984; Miller et al., 1993). When vitamin E was injected (usually in combination with selenium) rather than fed, about half the time there was no effect for prevalence of retained fetal membranes and about half the time there was a positive response (Miller et al., 1993).
The typical treatment was a single injection of approximately 700 IU of vitamin E and 50 mg of selenium given about 3 weeks before calving. Relative to the amount of vitamin E normally consumed, 700 IU of vitamin E over 21 days is trivial. Most likely, selenium, not vitamin E, was the nutrient responsible for the positive effect.
5.4 ACTION OF VITAMIN E AND SELENIUM ON NEUTROPHIL FUNCTION
Neutrophils are the first line of cellular defense against the aggression of invading pathogens. Thus, their recruitment to the site of pathogen attack will support defense mechanisms that favor reduction of the incidence of disease. The research on vitamin E and immunity in dairy cows has concentrated on neutrophil function. Vitamin E supplementation has consistently improved neutrophil function in dairy cows. The results from the two experiments that used fresh cows are noteworthy. The nonspecific immune system is depressed during the peripartum period and cows are extremely susceptible to uterine infections at this time. Both studies found that vitamin E supplementation eliminated the depression in neutrophil function associated with parturition and increases over all immune status.
Table 5. Research results on the effect of vitamin E supplementation on neutrophil function in dairy cows. | |||
Type of cow | Supplementation | Response | Ref. |
Lactating cow, 30 days in milk | 1000 IU/day of dietary vitamin E during the dry period and 500 IU/day during the first 30 days of lactation |
| Hogan et al., 1990 |
Fresh cow (<3 days in milk) | 3000 IU of vitamin E injected at 10 and 5 days before anticipated calving. |
| Hogan et al., 1992 |
Dairy cows from 4 wk pre to 5 wk postpartum | 3000 IU/d of dietary vitamin E from 4 wk pre to 8 wk postpartum + 3000 IU of vitamin E injected 1 wk prepartum |
| Politis et al., 1996 |
Fresh cows (<7 days in milk) | 3000 IU/d of dietary vitamin E from 4 wk pre to 8 wk postpartum + 3000 IU of vitamin E injected 1 wk prepartum |
| Politis et al., 1995 |
The positive effect of selenium supplementation on the health of dairy cows has necessitated the elucidation of the role of selenium in neutrophil function. Neutrophils release free radicals when they reach sites of infection to take part in the killing of pathogens. Oxygen free radicals such as superoxide anion, hydrogen peroxide and hydroxyl radical are generated in a process known as oxidative or respiratory burst. Although selenium deficiency has not been shown to affect neutrophil numbers in a range of species, it is clear that certain aspects of their function are affected.
In cattle, neutrophils collected from cows fed supplemental selenium diets had increased intracellular kill of bacteria, enhanced viability and reduced extracellular hydrogen peroxide concentration when compared with neutrophils from cows fed unsupplemented diets (Gyang et al., 1984; Hogan et al., 1990; Grasso et al., 1990). The role of selenium in these studies was associated with the activity of GSH-Px in the neutrophils. Under deficient conditions, a lack of/or decreased GSH-Px activity seems to reduce the ability to kill bacteria. On the other hand, increased GSH-Px activity in the presence of selenium enabled the continued production of radicals and bacterial clearance. Furthermore, Cebra et al., (2003) reported that neutrophils from postparturient dairy cows with higher blood concentrations of selenium appear to have greater potential to kill microbes. Considering the potential mechanisms of action of selenium, we hypothesize that selenium adequacy leads to production and increased abundance of certain selenoproteins (depending on the system and conditions) that increase the ability of neutrophils to kill pathogens. In addition, selenium adequacy will prolong the lifespan of neutrophils, thus enhancing their function. The antioxidant role of most selenoproteins already provides suitable conditions for neutrophil function. However, other unknown mechanisms may also be involved. Although several mammalian selenoproteins are known, their specific roles in cattle have not been investigated. Experiments are on-going or planned in various laboratory to test these hypotheses and characterize and determine the role of several selenoproteins in the bovine system.
•6 EFFECT OF SUPPLEMENTATION OF VITAMIN E AND SELENIUM ON REPRODUCTION
A single prepartum injection of vitamin E and selenium increased fertility (Arechiga et al. 1994). It was due to that vitamin E and selenium may have increased fertility by increasing uterine health due to the lower incidence of retained fetal membranes. Arechiga, et al., (1998) reported that service per conception was significantly reduced and conception rate was marginally higher in vitamin E and selenium supplemented cow. The report as depicted in table 5.
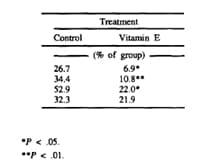
Table 6 Incidence of placental retention in dairy cows fed diets contain s.12 ppm of Se with or without 1000 IU of supplemental vitamin E during the last 40 d of gestation.
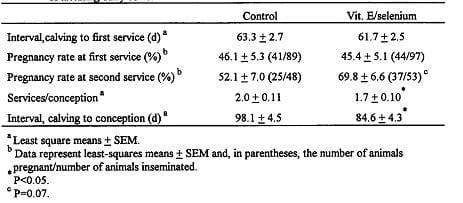
Table 7 Effect of postpartum administration of vitamin E and selenium on reproductive function Of lactating dairy cows. (Arechiga, et al., 1998)
Administration of vitamin E and/or selenium can also enhance neutrophil function (Either et al. 1994; Ndiweni and Finch 1996, Politis, et al. 1995; Politis, et al. 1996).Perhaps, increased neutrophil activity promotes removal of microorganisms and supports uterine tissue remodeling and involution. Prepartum treatment with selenium and vitamin E has been reported to hasten uterine involution in cows with metritis (Harrison et al. 1986). It is also possible that vitamin E and selenium affect events leading to fertilization since treatment with vitamin E and selenium increased fertilization rate in cattle (Segerson et al., 1981) and sheep (Segerson et al., 1977). This effect was ascribed to increased sperm transport (Segerson et al., 1980; Segerson et al., 1982) and may reflects actions of selenium on uterine motility (Segerson et al., 1980). Experiments in ewes indicate that selenium status can also affect embryonic survival (Hartley, 1963). It is not surprising that a single injection of vitamin E and selenium could cause a positive effect on reproductive function several weeks after the injection because administration of these molecules has long-term effects in cattle. Intramuscular injection of vitamin E caused elevated amounts in serum for at least 28 d (Charmley, et al. 1992), while injection of selenium increased concentrations of selenium in whole blood and serum for 28 d and increased whole blood-glutathione peroxidase activity for at least 84 d (Maas et al., 1993). Injections of vitamin E and selenium 3 and 1.5 wk before calving increased erythrocyte GSH peroxidase in dairy cows during the first 12 wk of lactation (Lacetera, et al., 1996).
Effects of selenium, vitamin E or their combination on fertility have been variable, with some studies reporting of an increase in fertility (Arechiga et al. 1994; Segerson et al., 1977; Segerson et al., 1981) and some reporting no effect (Stowe et al. 1988).Differences between studies in the amount of vitamin E and/or selenium administered, the period of administration, and nutritional status of the experimental animals with respect to vitamin E and selenium intake could explain some of these differential results.
Selenium deficiency adversely affects reproduction in both sexes and all species including humans (MacPherson, 1994). The problem is most extensively described in cattle and sheep; and while there have been reports of selenium deficiency as a direct cause of abortion, the related problems of increased disease susceptibility and retained placenta make it difficult to view infertility in the dam in terms of a single factor (Maas,1998). Selenium supply to particular tissues may also be important. Conrad (1985) reported that uterine involution was completed eight days sooner in cows supplemented with selenium and vitamin E. He also found that increasing whole blood selenium was associated with higher selenium in the ovary and that GSHPx activity in follicular fluid correlated with plasma GSH-Px. Significant GSH-Px activity was present in luteal tissue as well.
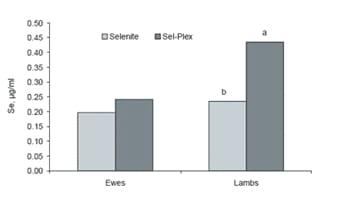
Figure 2. Effect of selenium supplement form on whole blood selenium content of ewes at lambing and newborn lambs (from Kincaid et al., 1999; abP#0.001).
Egg production is depressed by selenium deficiency in both chickens and turkeys; however the impact on hatchability is more severe in turkeys (Cantor and Scott, 1974; Cantor, 1997). While the role of selenium status in female fertility is more recognized than understood, recent work has provided much clearer insight into the well known association between selenium and male fertility through identification of the membrane-bound phospholipid hyeroxide GSH-Px in the testes.
6.1 EFFECT OF SUPPLEMENTATION OF VITAMIN E AND SELENIUM ON OVA FERTILIZATION
Fertility of ova was greater (P<.O05) in adequately-fed, selenium and vitamin E supplemented ewes (Segerson and Ganapathy, 1981) and beef cows (Segerson et al., 1977). Segerson et al. (1977) proposed that if Se/vitamin E supplementation is needed, one or both of the nutrients may influence uterine muscular function in a manner that increases uterine contractions. Ewes in the with vit E ans Se supplementation had a greater (P<.05) proportion of fertile ova than did ewes in the other groups. This observation and further data obtained at estrus which showed that ewes in the se- vitamin E supplemented group exhibited more (P<.05) total uterine contractions and more (P<.025) contractions migrating toward the oviduct than ewes in the non supplemented group. The finding that more uterine contractions migrated toward the oviduct at estrus than toward the cervix or in neither direction is in agreement with migration patterns previously reported. Previous studies with dairy cattle have demonstrated the efficacy of Se and Se plus vitamin E in decreasing the incidence of retained placenta, suggesting a possible effect of one or both of these nutrients on uterine muscular function. Julien et aI. (1976) indicated that Se was the more important of the two nutrients in reducing the incidence of retained placenta. Brown and Burk (1973) demonstrated that the rat uterus incorporates a quantity of the isotope, 7SSe, equaling the isotope incorporation of quadricep and cardiac muscles, suggesting that Se is involved in rat uterine function. Much previous work with musculature has clearly shown that Se is needed in young ruminants to prevent lesions and degenerative muscular dystrophy of cardiac and skeletal muscles (Underwood, 1977). Furthermore, characteristic abnormalities in the electrocardiograms of lambs afflicted with nutritional muscular dystrophy have been documented.
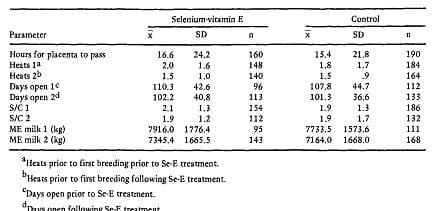
Table 8 Means and standard deviations for reproduction and lactation for the lactation prior to and following selenium-vitamin E treatment. (Gwazdauskas et al., 1979)
6.2Effect of supplementation of Vitamin E and Selenium on male reproduction and conception rate
Vitamin E deficiencies caused testicular degeneration in chickens, rats, hamsters, rabbits, guinea pigs, dogs, cats, pigs, and monkeys and resulted in a lower number of germ cells and a reduction in sperm production (cited from the Marin-Guzman et al., 1997). Brzezinska- Slebodzinska et al. (1995) suggested that dietary vitamin E may serve as an antioxidant in boar semen. Jones and Mann (1977) demonstrated that peroxidative structural damage to sperm occurred when unsaturated fatty acids were present in semen; this resulted in a decline in sperm motility, but the addition of tocopherol to the ejaculate did not protect the sperm from peroxide damage. Because Se and vitamin E can affect testicular and(or) spermatozoal development and subsequent sperm motility, the deficiency of either nutrient may affect different aspects of the male reproductive processes and possibly the fertilization of oocytes in the female. Diets inadequate in Se and vitamin E fed for prolonged periods will affect the reproductive efficiency of male animal. Testicular tissue has a high priority for Se retention and utilization compared to liver. When low-Se and vitamin E diets were fed, sperm motility declined and the percentage of abnormal sperm increased. Boars fed low-Se diets produce a higher percentage of abnormal sperm, mainly by disrupting tail morphology. Boars with a low Se status had fewer sperm reaching and penetrating the zona pellucida, and this could affect ovum fertilization rate. Although a diet low in vitamin E can also reduce the percentage of normal sperm, vitamin E seems important for boars, but it may function in a different manner than Se, perhaps through its antioxidant properties on the sperm.
In sperm, selenium is largely associated with the keratin-like material in the mitochondrial helix in the midpiece of spermatozoa and was previously referred to as the ''''mitochondrial capsule selenoprotein''''. Recently it has been determined that GSH-Px-PH is abundantly expressed in spermatogenic cells, but exists as an enzymatically-inactive structural protein in mature sperm where it contributes to formation of the mitochondrial capsule (Ursini et al., 1999). In spermatozoa, GSH-Px-PH therefore replaces ''''sperm capsule selenoprotein'''' as the link between selenium and fertility. Morphological defects of sperm in selenium-deficient males can be attributed to inadequate GSH-Px synthesis (Köhrle et al., 2000).
7. ACTION OF VITAMIN E AND SELENIUM ON OTHER NON-INFECTIOUS DISEASES
There has been much less work to investigate the effect of vitamin E on other diseases, and most of it has been peripheral to the main investigation. For instance, Miller and others (1993) demonstrated that supplementing primiparous cows with 1000 iu of vitamin E reduced the incidence of udder oedema when their diet contained at least 0-12 mg of selenium per kg DM, but not when their diet contained less than 0-07 mg of selenium per kg DM. Supplementing cows with vitamin E has been shown to have significant effects on the incidence of disease in some studies, but not in others. Some of the studies that have observed no effect have supplied vitamin E in insufficient amounts for too short a period, or at inappropriate times. Virtually all of the published results are based on North American data. The dairy industry in the USA and Canada is significantly different from that in the UK and most of Europe, and a large scale field study of the effect of vitamin E supplementation is required in Europe.
•8. OPTIMUM LEVEL OF VITAMIN E REQUIREMENT
The NRC (2001) established requirement for dietary vitamin E is 16 -27 IU/kg DMI for nonlactating and lactating cows depend on lactation, equivalent to consumption of 160 and 900 IU/d for nonlactating and lactating cows, respectively.
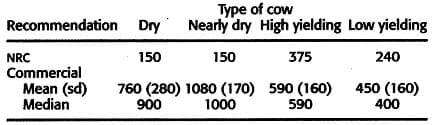
Table 9 NRC and commercial recommendation for Vitamin E intake (IU per day) dairy cow
To maintain these blood values, dry cows and heifers fed stored forages during the last 60 days of gestation require approximately 1.6 IU of supplemental vitamin E/kg of body weight (approximately 80 IU/ kg of DMI). An additional benefit on calf health may be observed by increasing vitamin E intake by cows and heifers in late gestation. Only minor amounts of vitamin E can pass the placenta (Van Saun et al., 1989); hence newborn calves rely on colostrum for vitamin E. Increased intake of vitamin E during the prepartum period significantly elevates vitamin E in colostrum. For lactating cows, the recommended amount of vitamin E supplemental) was changed to 0.8 IU/kg of body weight (approximately 20 IU/kg of DMI) when stored forages are fed. This recommendation is based on a reduction in retain placenta and other post partum infection, there by improve over all fertility of the animal. The difference between the recommendations for vitamin E for the two classes of cattle is mainly caused by expected differences in intake of vitamin E from basal feedstuffs and perhaps reduced absorption of vitamin E by cows fed conventional dry cow diets.
Based on typical feed intakes and average vitamin E concentrations in feedstuffs, the recom mended amount of total vitamin E (supplemental plus vitamin provided by feedstuffs) is approximately 2.6 IU/ kg of body weight during late gestation and for lactating dairy cows. Of that amount, the basal diet will provide on average about 1.8 IU/kg of body weight for lactating cows (ranges from about 0.8 for cows fed diets based on severely weathered hay to about 2.8 IU/kg of body weight for cows fed diets based on pasture) and about 1 IU/kg body weight (ranges from 0.5 to about 2.3 IU/kg of body weight) for dry cows.
Although several factors are known to influence vitamin E requirements, limited data make quantifying the necessary adjustments difficult. The amount of supplemental vitamin E fed may need to be changed during the following situations:
- When fresh forage is fed there should be less need for supplemental vitamin E. A diet based on fresh forage (ca. 50 percent of dietary DM) would require about 67 percent less supplemental vitamin E to meet the cows requirements compared with a diet that contained a similar amount of stored forage. Requirements for supplemental vitamin E is reduced 67 percent in the accompanying software when animals are fed pasture.
- The amount of supplemental vitamin E probably should be increased when low forage diets are fed (forages typically have more vitamin E than do concentrates). The requirements listed above were generated from studies using diets with 50 to 60 percent forage (lactating cows) and 60 to 80 percent forage (animals in late gestation).
- Cows in suboptimal selenium status probably require more vitamin E.
- Milk is not a major excretion route for alpha-tocopherol(0.4 to 0.6 g/ml) but colostrum contains high concentrations of -tocopherol (3 to 6 g/ml). Additional vitamin Emay be useful during colostral genesis.
- Intake of polyunsaturated fatty acids increases the vitamin E requirement of nonruminants. As methods for protecting fats from biohydrogenation in the rumenimprove, additional vitamin E may be required when protected unsaturated fats are fed.
- Additional vitamin E may be useful during periods of immunosuppression (peripartum period).
- Large amounts of supplemental vitamin E (1000IU/day) can reduce oxidative flavors in milk.
12 OPTIMUM LEVEL OF SELENIUM REQUIREMENT
In selenium-deficient locales, dietary intakes of .3 mg of selenium/kg of dry matter are necessary to have some degree of confidence that serum selenium concentrations will be adequate in dairy cattle. A reduction of the legal supplementation allowance to . 1 mg of supplemental selenium/kg of dry matter would have serious negative consequences. Reducing the allowable selenium supplementation level would result in many herds being inadequately supplemented in selenium deficient areas and presumably many more instances of clinical and subclinical selenium deficiency. Productivity losses would be significant in the national dairy herd, much of which is located in selenium-deficient areas.
The level of selenium supplementation in dairy cow diets varies from country to country and is mainly determined by the selenium content of feedstuffs, as influenced by soil selenium status. In Canada and the United States, both organic and inorganic sources of selenium can be supplemented at 0.3 mg/kg of DM. In the European Union, inorganic selenium is fed to cows at a recommended level of 0.5 mg/kg of DM (Ministry of Agriculture, Fisheries and Food, 2000) and was initially the only authorized form of selenium.
Since selenium was originally known to be toxic, it is imperative that care be taken not to reach toxic levels during supplementation. Rate of supplementation in most trials does not consider the selenium concentration in the other feed ingredients, implying that the final selenium intakes of animals is usually more than current recommended levels. However, little data exist to suggest toxic effects even when selenium was included at levels as high as 50 mg/d for 90 days or 100 mg/day for 28 days in diets of Holstein cows (Ellis et al., 1997).
Givens at al. (2004), as in most studies, observed increasing advantage of selenium inclusion levels on milk selenium concentration with the best results at a rate of 1.14 mg Se/kg as selenium yeast. These results demonstrate that current recommended levels of inclusion should be revisited in order to set the limits for favorable performance and to avoid toxicity. Since blood selenium concentration is a fairly good variable to measure individual or herd selenium status, Hogan et al. (1993) recommended its use in establishing selenium status. Their recommendation follows that blood selenium concentration should be at least 0.2 μg/ mL, but should not exceed 1 μg/mL.
On the other hand, Braun et al. (1991) considered blood selenium levels of 0.08 to 0.3 μg/mL as normal and 0.03 to 0.07 μg/mL as inadequate for cattle. To ensure that herd selenium intakes are within safe limits, it is imperative that intakes should not exceed the toxic threshold of selenium supplementation for dairy cows. Stating the selenium concentration on feed tags and routinely measuring concentration level in forages, hay and other feed materials may be one way of ensuring safe and favorable levels of inclusion and intakes.
10 OTHER UNEXPLORED EFFECTS OF SELENIUM IN DAIRY CATTLE
A further area where selenium seems to play a role, but not yet examined in dairy cattle, is in apoptosis. The ultimate goal of a cell is to perform its function to the best of its ability in the maintenance of tissue homeostasis. Apoptosis is the process of deliberate life relinquishment by a cell in an organism. It is also commonly referred to as ''''cell suicide.''''
In humans, an insufficient amount of apoptosis leads to uncontrolled cell proliferation, such as cancer whereas excessive apoptosis causes cell loss diseases such as ischemic damage (shortage of blood supply to an organism). Data in 1996 indicated that dietary supplementation of 200 μg of selenium in diets resulted in a decreased incidence of human prostate, lung and colorectal cancers (Clark et al., 1996).
The precise role of selenium in modulating cancers is much debated. Some authors are of the opinion that small molecules of selenium metabolites selectively promote apoptosis in transformed prostate epithelium (Jiang et al., 2004; Ip et al., 2000) while others believe that selenium supplementation prevents DNA damage that could lead to cell transformation by increasing levels of antioxidant selenoproteins (Diwadkar- Navsariwala and Diamond, 2004; Lu and Jiang, 2005).
A recent study with transgenic mouse models showed that selenoprotein deficient mice exhibited accelerated development of lesions associated with prostate cancer progression thus suggesting that selenium may function in cancer prevention by modulating the levels of selenoproteins (Diwadkar- Navsariwala et al., 2006). Another finding demonstrates that selenoprotein expression and selenium metabolism are regulated at multiple levels in prostate cells (Rebsch et al., 2006).
Selenium may therefore play a positive role in mediating useful apoptosis necessary for normal functioning of cells. Selenium''''s role in this regard, is however, yet to be established in cattle.
11.CONCLUSION
The periparturient and early lactation periods are critical for the health of dairy cows. Udder edema, milk fever, retained placenta, mastitis, anoestrus and suboptimal reproduction reduce profits for dairy producers. Oxidative stress may contribute to all of these disorders. Antioxidant requirements of high producing dairy cows may be higher than generally recognized, and intakes of antioxidants needed to control ROM balance effectively may exceed amounts supplied by average feeds. For this reason, supplementation with all known nutrients required for antioxidant defense in adequate and balanced amounts would be beneficial. The primary effect of vitamin E supplementation appears to involve the activity and effectiveness of the immune system. Some studies have observed an effect on the lymphocyte system, others have observed an increase in the activity of neutrophils. In several studies there has been a reduction in the incidence of retained fetal membranes after vitamin E supplementation. There is some evidence for a reduction in cystic ovarian disease and metritis.
In the studies in which vitamin E supplementation had a beneficial effect on fertility, the vitamin E intake of the cows during the dry period has been increased to about 3000 iu per cow per day. Recent research shows that vitamin E and selenium had beneficial effect on reducing the incidence of anestrous, reduces number of services per conception, reduces calving interval, increases conception rate and over all fertility of the cattle.
However, more experiment is needed to study on reproductive disorder with supplemental vitamin E and Selenium for treatment and for further reproductive performance of the animal and to identify optimal amounts of each nutrient Vitamin E and selenium supplementation on the health and some aspects of the fertility of lactating dairy cows. These effects have been observed not only in cows which are deficient in vitamin E or selenium, but also in studies in which supplemented animals were compared with control cows fed diets containing up to three times the current recommendations for vitamin E intake.
"One calf per annum is dream of farmer For this, I should come estrus proper" "Feed me antioxidant to protect my endometrium Vitamin E and Se can be given optimum" "This will increase the conception rate This quote for my interest" |
12 REFERENCE
Aitken RJ. A free radical theory of male infertility. Reprod Fertil Dev 1994;6:19-23.
Arechiga CF, Ortiz O, Hansen PJ. 1994. Effect of prepartum injection of vitamin E and selenium on postpartum reproductive function of dairy cattle. Theriogenology 41:1251-1258.
Arechiga, C. F., S. Vazquez-Flores, O. Ortiz, J. Hernandez-Ceron, A. Porras, L. R. McDowell, and P. J. Hansen. 1998. Effect of injection of beta-carotene or vitaminE and selenium on fertility of lactating dairy cows. Theriogenology 50:65-76.
Ascarelli I, Edelman Z, Rosenberg M, Folman Y. Effect of dietary carotene on fertility of high yielding dairy cows. Anim Prod 1985;40:195-207.
Atroshi. E, J. Tyopponen, S. Sankari, R. Kangasniemi. and J. Parantainen. 1986. Possible roles of vitamin E and glutathione metabolism in bovine mastitis. Int. 1. Vitam. Nut. Res. 57:37.
Atroshi. F., J. Parantainen. S. Sankari. and T. Osterman. 1986. Prostaglandins and glutathione peroxidase in bovine mastitis. Res. Vet. Sci. 40:361.
BARNES, M. M. C. & SMITH, A. J. (1975) The effects of vitamin E deficiency on some enzymes of steroid hormone biosynthesis. International Journal of Vitamin Nutrition Research 45, 396-403
Bindas EM, Gwazdauskas FC, McGilliard ML, Polan CE. 1984. Progesterone responses to human chorionicgonadotropinin dairycattlesupplementedwith β-carotene. J. Dairy Sci;67:2978- 2985.
Boyd, H., 1977. Anoestrus in cattle. Vet. Rec. 100, 150-153.
Brown, D. G. and R. F. Burk. 1973. Selenium retention in tissues and sperm of rats fed a Torula yeast diet. J. Nutr. 102:102.
Brzezinska-Slebodzinska, E., A. B. Slebodzinski, B. Pietras, and G. Wieczorek. 1995. Antioxidant effect of vitamin E and glutathione on lipid peroxidation in boar semen plasma. Biol. Trace Elem. Res. 47:69-74.
Cantor, A.H. 1997. The role of selenium in poultry nutrition. In: Biotechnology in the Feed Industry, Proceedings of the 13th Annual Symposium. (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, Nottingham, UK.
Cantor, A.H. and M.L. Scott. 1974. The effect of selenium in the hen''''s diet on egg production, hatchabihty, performance of progeny and selenium concentrations in eggs. Poultry Sci. 53:1870.
Carlson JC, Wu XM, Sawada M. Oxygen radicals and the control of ovarian corpus luteum function. Free Pad Biol Med 1993;14:79-84.
Jones, R., and T. Mann. 1977. Toxicity of exogenous fatty acid peroxides towards spermatozoa. J. Reprod. Fertil. 50:255-260.
Charmley E, Hidiroglou N, Ochoa L, McDowell LR, Hidiroglou M. 1992. Plasma and hepatic a- tocopherol in cattle following oral or intramuscular supplementation. J Dairy Sci;75:804-810
CIPRIANO, J. E., MORRILL, J. L. & ANDERSON, N. V. (1982) Effect of dietary vitamin E on immune responses of calves. Journal of Dairy Science 65,2357- 2365
Conrad. H.R. 1985. The role of selenium and vitamin E in bovin reproduction. In: Selenium Responsive Diseases in Food Animals. Proc. Symposium, Western States Veterinary Conference, Las Vegas, NV.
Dentine. M. R., and B. T. McDaniel. 1983. Variation of edema scores from herd-year, age, calving month, and sire. J. Dairy Sci. 66:2391.
Either SD, Morrill JL, Blecha F. Vitamin concentration and function ofleukocytes fi''''om dairy calves supplemented with vitamin A, vitamin E, and 13-carotene in vitro. J Dairy Sci 1994; 77:560-565.
El-Azab M A, Kakoma I, Brodie B O. McKenna D J and Gustafusson K. 1988. Bacteriology of the postpartum bovine uterus with retained placenta and induced metritis: Special reference to minimum inhibitory concentration (MIC). In: Proceedings of the 11th International Congress on Animal Reproduction and Artificial Insemination, 26-30 June 1988. University College, Dublin, Ireland. p. 513.
Erskine, R. J., R. J. Eberhart, L. J. Hutchinson, and R. W. Scholz. 1987. Blood selenium concentrations and glutathione peroxidase activities in dairy herds with high and low somatic cell counts. J. Am. Vet. Med. Assoc. 190:1417.
GOTO, Y., NODA, Y., NARIMOTI, K., UMAOKA, Y. & MORI, T. (1992) Oxidative stress on mouse embryo development in vitro. Free Radical Biology and Medicine 13,47-53.
GWAZDAUSKAS, F. C., BIBB, T. L., McGILLIARD, M. L. & LINEWEAVER, J. A. (1979) Effect of prepartum selenium-vitamin E injection on time for placenta to pass and on productive functions. Journal of Dairy Science 62,978-981.
HADDEN, J. W. (1987) Neuroendocrine modulation of the thymus-dependant immune system. Annals of the New York Academy of Science 496, 39
Harrison JH, Hancock DD, St. Pierre N, Conrad HR, Harvey WR. 1986 Effect ofprepartum selenium treatment on uterine involution in the dairy cow. J Dairy Sci;69:1421-1425.
Harrison, J. H., D. D. Hannock, and H. R. Conrad. 1984. Vitamin E and selenium for reproduction of the dairy cow. J. Dairy Sci. 67: 123.
Hartley WJ. Selenium and ewe fertility. Proc NZ Soc Anim Prod 1963;923:20-27.
Hogan, J. S., K. L. Smith, W. P. Weiss, D. A. Todhunter, and W. L. Shockey. 1990. Relationships among vitamin E, selenium, and bovine blood neutrophils. J. Dairy Sci. 73:2372.
HOGAN, J. S., WEISS, W. P. & SMITH, K. L. (1993) Role of vitamin E and selenium in host defence against mastitis. Journal of Dairy Science 76,2795-2803.
HOGAN, J. S., WEISS, W. P., SMITH, K. L., SORDILLO, L. M. & WILLIAMS, S. N. (1996) Alpha-tocopherol concentrations in milk and plasma during clinical Escherichia coli mastitis. Journal of Dairy Science 79,71-75.
HOGAN, J. S., WEISS, W. P., TODHUNTER, K. L., SMITH, K. L. & SCHOENBERGER, P. S. (1992) Bovine neutrophil responses to parenteral vitamin E. Journal ofDairy Science 76,399-405
Joosten, I.. J. Stelwagen, and A. A. Dijkhuiim. 1988. Economic and reproductive consequences of retained placenta in dairy cattle. Vet. Rec. 12353.
Julien, W. E., H. R. Conrad, J. E. Jones and A. L. Moxon. 1976. Selenium and vitamin E and incidence of retained placenta in parturient dairy cows. J. Dairy Sci. 59:1954.
KIGOSHI, S. & ITO, R. (1973) High levels of free fatty acids in lymphoid cells, with special reference to their cytotoxicity. Experientia 15,1408-1410.
Kincaid, R.L., M. Rock and F. Awadeh. 1999. Selenium for ruminants: comparing organic and inorganic selenium for cattle and sheep. In: Biotechnology in the Feed Industry, Proceedings of the 15 Annual Symposium.(T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, UK.
Köhrle, J., R. Brigelius-Flohe, A. Bock, R. Gartner, O. Meyer and L. Flohe. 2000. Selenium in biology: facts and medical perspectives. Biol. Chem. 381(9-10):849-864.
Lacetera N, Bernabucci U, Ronchi B, Nardone A. 1996. Effects of selenium and vitamin E administration during a late stage of pregnancy on colostrum and milk production in dairy cows,and on passive immunity and growth of their offspring. Am J Vet Res;57:1776-1780.
Lamming GE 1980 Milk progesterone for assessing response to treatment of sub fertile cattle [ Proc 8th Int Congr Anim Reprod and AI, Madrid, vol II pp143.151
Laven R. A and Peters, A. R. 1996. Bovine retained placenta: aetiology, pathogenesis and economic loss. Veterinary Record 139: 465-471
Maas J, Peauroi JR, Tonjes T, Karlonas J, Galey FD, Bin H. 1993. Intramuscular selenium:administration in selenium-deficient cattle. J Vet Int Med;7:342-348.
Maas, J. 1998. Studies on selenium metabolism in cattle: deficiency, supplementation and environmental fate of supplemented selenium. In: Selenium-Tellurium Development Association, 6th International Symposium, May 10-12, Scottsdale, AZ.
MacPherson, A. 1994. Selenium, vitamin E and biological oxidation. In: Recent Advances in Animal Nutrition (P.C. Garnsworthy and D.J.A. Cole, eds). Nottingham University Press, Nottingham, UK.
Marin-Guzman,J, D. C. Mahan, Y. K. Chung, J. L. Pate and W. F. Pope.1997. Semen quality, and subsequent fertilization rates in mature gilts Effects of dietary selenium and vitamin E on boar performance and tissue responses. J Anim Sci 75:2994-3003.
MILLER, J. K., BRZEZINSKA-SLEBODZINSKA, E. & MADSEN, F. C. (1993)Oxidative stress, antioxidants, and animal function. Journal of Dairy Science 76: 2812-2823
Miller, J. K., E. Brzezinska-Slebodzinska, and F. C. Madsen. 1993. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 76:2812.
Miller, J. K., F. J. Mueller, D. G. Thomas, and F. C. Madsen. 1991. Vitamin E and reproduction in dairy cows. In: M. B. Coelho (Ed . ) Vitamin E in Animal Nutrition and Management. p 159. BASF, Parsippany, NJ.
Mohanty B N. Parihar N S and Luktude S N. 1980. Tuberculosis of genital organs causing repeat breeding in cattle. Indian Veterinary Journal 57: 859-860.
MUDRON, P., KOVAC, G., BAJOVA, V., PISTL, J., CHROMA, J., BARTKO, P. & SCHOLZ, H. (1994) Effect of vitamin E on some leucocytic parameters and functions in transported calves. Deutsche Tierarztliche Wochenschift 101, 47-49.
Namboothripad T R B and Raja C K S V. 1976. A study of the aetiology of repeat breeding in cows. Kerala Journal of Veterinary Science 7: 195
Ndiweni N, Finch JM. 1996. Effects of in vitro supplementation with a-tocopherol and selenium on bovine neutrophil functions: implications for resistance to mastitis. Vet Immunol Immunopathol;51:67-78.
Paape, M. J., D.B. Bannerman, X. Zhao and J.-W. Lee. 2003. The bovine neutrophil: structure and function in blood and milk. Vet. Res. 34:597-627.
Politis I, Hidiroglou N, Batra TR, Gilmore JA, Gorewit RC, ScherfH. 1995 Effects of vitamin E on immune function &dairy cows. Am J Vet Res;56:179-184.
Politis I, Hidiroglou N, White JH, Gilmore JA, Williams SN, ScherfH, Frigg M. 1996. Effects of vitamin E on mammary and blood leukocyte function, with emphasis on chemotaxis, in periparturient dairy cows. Am J Vet Res;57:468-471.
Putman, M. E., and N. Comben. 1987. Vitamin E. Vet. Rec. 121:541.
RAPOPORT, R., SKLAN, D., WOLFENSON, D., SHAHAM-ALBALANCY, A. & HANUKOGLU, I. (1998) Antioxidant capacity is correlated with steroidogenic status of the corpus luteum during the bovine oestrus cycle. Biochimica et Biophysica Acta 1380, 133-140
Rayman, M.P. 2000. The importance of selenium to human health. The Lancet 356:233- 241.
REDDY, P. G., MORRILL, J. L., MICOCHA, H. C. & STEVENSON, J. S. (1987) Vitamin E is immunostimulatory in calves. Journal of Dairy Science 70,993- 999.
Rice, D. A., and S. Kennedy. 1988. Assessment of vitamin E, selenium and polyunsaturated fatty acid interactions in the aetiology of disease in the bovine. Proc. Nutr. Soc. 47:177.
Sanchez, J. P. Montes, A. Jimenez, and S. Andres. 2007. Prevention of Clinical Mastitis with Barium Selenate in Dairy Goats from a Selenium-Deficient Area. J. Dairy. Sci. 2007 90: 2350-2354.
Segerson, E. C., W. R. Getz, and B. H. Johnson. 1981. Selenium and reproductive function in boars fed a low selenium diet. J. Anim. Sci. 53:1360-1367.
Segerson EC Jr, Murray FA, Moxon AL, Redman DR, Conrad HR. 1977. Selenium/vitamin E: role in fertilization of bovine ova. J Dairy Sci; 60:1001-1005.
Segerson EC, Libby DW. 1982 Ova fertilization and sperm number per fertilized ovum for selenium and vitamin E-treated Charolais cattle. Thariogenology 1982;17:333-341.
Segerson EC, Riviere G, Bullock TR, Thimaya S, Ganapathy SN. 1980. Uterine contractions and electrical activity in ewes treated with selenium and vitamin E. BiolReprod;23:1020-1028.
SegersonEC, GanapathySN 1981. Fertilization of ova in selenium/vitamin E-treated ewes maintained under two planes of nutrition. J Anim Sci 1981;51:386-394.
Shanks, R. D., A. S. Freeman. and F. N. Dickinson. 1981. Postpartum distribution of costs and disorders of health. J. Dairy Sci. 64:683.
Singh C S P. Singh S K and Singh B. 1981. Studies on the incidence of infertility in cows. Indian Veterinary Journal 58: 909-912 (Veterinary Bulletin 51: 7266).
Smith, K. L., J. H. Harrison, D. D. Hancock, D. A. Todhunter, and H. R. Conrad. 1984. Effect of vitamin E and selenium supplementation on incidence of clinical mastitis and duration of clinical symptoms. J. Dairy Sci. 67:1293.
Spears, J.W. 2000. Micronutrients and immune function in cattle. Proc. Nutr. Society 59: 587-594.
STAATS, D. A., LOHR, D. P. & COLBY, H. D. (1988) Effects of tocopherol depletion on the regional differences in adrenal microsomal lipid peroxidation and steroid metabolism. Endocrinology 123,975-980.
Stapleton, S.R. 2000. Introduction: the selenium conundrum. Cell Mol. Life Sci. 57:1823- 1824.
Stowe HD, Thomas JW, Johnson T, Marteniuk JV, Morrow DA, Ullrey DE. Responses of dairy cattle to long-term and short-term supplementation with oral selenium and vitamin E. J Dairy Sci 1988;71:1830-1839.
Thirunavukkarasu, M.2006.Conception rates in artificially inseminated bovines - a field survey. Tamilnadu J. Veterinary & Animal Sciences 2(5): 186-187.
Underwood, E. J. 1977. Selenium. In Trace Elements in Human and Animal Nutrition (3rd Ed.).
Ursini, F., S. Heim, M. Kiess, M. Maiorino, A. Roveri, J. Wissing and L. Flohe. 1999. Dual function of the selenoprotein PHGPx during sperm maturation. Science 285:1393-1396.
Vandeplassche M. 1982. Reproductive efficiency in castle: A guideline for projects in developing countries. FAO Animal Production and Health Paper No. 25. FAO (Food and Agriculture Organization of the United Nations), Rome, Italy. 118 pp.
Watson P F. 1979. The preservation of semen in mammals. Oxford Review of Biology 1: 283.
Malbe, M., M. Klaassen, W. Fang, V. Myllys, M. Vikerpuur, K. Nyholm, W. Sankari, K. Suoranta, and M. Sandholm. 1995. Comparisons of selenite and selenium yeast feed supplements on Se-incorporation, mastitis, and leukocyte function in Se-deficient dairy cows. J. Vet. Med. (Ser. A) 42:111-121.
Rotruck, J. T., A. L. Pope, H. E. Ganther, A. B. Swanson, D. G. Hafeman, and W. G. Hoekstra.1973. Selenium. Biochemical role as a component of glutathione peroxidase. Science 179:588.
Ortman, K., and B. Pehrson. 1999. Effect of selenate as a feed supplement in dairy cows in comparison to selenite and selenium yeast. J. Anim. Sci. 77:3365-3370.
Gyang, E. O., J. B. Stevens, W. G. Olson, S. D. Tsitsamis, and E. A. US 1984. Effects of seleniumvi- E injection on bovine polymorphonucleated leukocytes phagocytosis and killmg of Sruphylococcus uureus. Am. J. Vet. Res. 45:175.
Grasso, P. J., R. W. Scholz, R. J. Erskine, and R. J. Eberhart. 1990. Phagocytosis, bactericidal activity, and oxidative metabolism of mammary neutrophils from dairy cows fed selenium-adequate and selenium deficient diets. Am. J. Vet. Res. 51:269.
Cebra, C. K., J. R. Heidel, R. O. Crisman, and B. V. Stang. 2003. The relationship between endogenous cortisol, blood micronutrients, and neutrophil function in postparturient Holstein cows. J. Vet. Intern. Med. 17:902-907.
Related topics
Authors:

Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post



You may be interested in
