Introduction
Over millennia, grazing ruminants have, for the most part, evolved on the great grasslands of the world, ranging from the savanna and tropical grasslands of the equatorial regions to the temperate grasslands at slightly higher latitudes. Less extensive are the high-altitude alpine pasture and marginal tundra biomes, in which many unique distinctive breeds and species of livestock have evolved, which, although numerically fewer, are nonetheless important to various, often migratory indigenous peoples. Due to seasonal changes in the quantity, morphology and mass of herbage species within these varied ecosystems, we should not be surprised to find that their ruminant species have evolved a range of strategies to optimise energy capture and utilisation in their changing environment.
Over recent decades, much research has been conducted to investigate the effect of differences in sward structure and composition on grazing behaviour of various economically important domesticated ruminants including sheep, beef or dairy cattle, buffaloes and yaks. However, for the purposes of the present paper, we will concentrate mainly on work conducted with dairy cattle, drawing on data from alternative sources only where these are absent for dairy breeds.
Black and Kenney (1984) used a novel approach of constructing artificial swards (sward boards) to examine the effects of sward height and density on diet selection by sheep. Subsequently, Laca et al. (1992a, 1992b) used a similar approach to examine the relationships between sward structure and mechanical aspects of the grazing process (bite mass, depth and area) by cattle. While such approaches were undoubtedly useful in demonstrating the functional relationships between sward characteristics and bite dimensions, as Penning (1986) pointed out, it is extremely difficult to replicate sward conditions found under field conditions, and such short-term measurements may not be representative of those achieved on swards in the field. More recently, Orr et al. (2005) developed a technique to grow swards of differing structure and different varieties in boxes (0.374 m2 ), which could be presented to cattle for shortterm grazing studies. The technique was used by Soder et al. (2009) to examine the relationship between sward surface height (SSH) in different grass species and bite mass.
In addition to these short-term studies, experiments to test various hypotheses concerning factors thought to initiate and terminate eating activity have been conducted. Chilibroste et al. (1997, 1998) used individually tethered dairy cows (a grazing system still used by small dairy farmers in many areas around the world) to examine the effect of the interaction of artificial rumen fill and fasting before grazing, on the length of the first grazing bout, dry matter intake (DMI), short-term intake rate (IR), bite mass, bite rate, rumen fill and rumen fermentation. Chilibroste et al. (2000) used the same technique to examine the effect of sward height before grazing on the same response variables. Subsequently, Gregorini et al. (2009a, 2009b) used a similar approach to examine the effect of rumen fill on grazing behaviour, hunger-related hormone circulation and metabolite concentrations in dairy cows offered micro-swards.
However, while such short-term measurements are useful, they fail to represent the normal situation in which grazing ruminants find themselves. First, they do not represent the heterogeneity and complexity of field swards. Grazing activity itself continually modifies the sward at differing rates depending on management, and invariably results in heterogeneous patches differing in height, plant density, the ratio of plant tissues (e.g. leaf lamina, pseudostem, dead material) and even plant species. Second, they lack the time element of 24-h or longer periods. Many of these previously mentioned fundamental studies have been conducted over short time scales (e.g. minutes to few hours), which raises the question to what extent the underlying mechanisms remain unchanged throughout the day. While sward structure may constrain the mechanics of the grazing process, grazing ruminants are able to vary both the duration and the temporal pattern of grazing meals in response to constraints on IR and various internal conditions (e.g. lactation or hunger).
For the most part, earlier larger-scale studies with dairy cows were conducted single- or two-species temperate pastures, carefully managed to present the animals either with swards in which structure (i.e. height, mass and spatial heterogeneity) was maintained for the duration of the measurements (e.g. by continuous variable stocking), or in which the rate of modification of the sward structure was manipulated by controlling the grazing pressure (frequently defined in terms of ‘herbage allowance’). In this way, the effects of differences in sward structure and composition, and animal species, breed and physiological state on what may be described as the mechanics of the grazing process (bite mass, bite rate and IR), temporal aspects of feeding (principally grazing activity, but also ruminating and idling) and dietary selection have been studied (e.g. Gibb et al. 1999; McGilloway et al. 1999; Barrett et al. 2001; Rutter 2006).
Complementarily to this fundamental research, more applied experiments have been conducted to test whether these fundamental relationships can be assumed to apply under the conditions of farm feeding management and on pastures other than intensively managed, predominantly perennial ryegrass, temperate swards (e.g. Chilibroste et al. 2007, 2012; Mattiauda et al. 2013; Meikle et al. 2013; Soca et al. 1999, 2014). For lactating dairy cows, the main feeding management practices involve decisions at pasture level (changes in sward mass, herbage allowance, sward height), animal status (physiological state, fasting before grazing), feeding strategy (offer of supplements: level, type and timing), grazing management (continuous stocking vs daily strip, access time and timing of the grazing sessions) and available infrastructure (such as e.g. rest areas, pad conditions, walking distance). In recent years, milking frequency has also emerged as an issue, since grazing dairy cows that use to be milked following a regular schedule (usually two milkings with a fixed interval between morning and afternoon milkings) can now be milked on a voluntary basis, varying the frequency and interval between successive milkings (Lyons et al. 2014). Applied research has confirmed the relevance of short-term events (e.g. bite mass and IR) on daily DMI and eventually animal performance. However, it is much less conclusive concerning the potential to modify ingestive behaviour (e.g. temporal patterns of grazing and total grazing time). Indeed, few either fundamental or applied research studies have even reported information about grazing patterns along the day.
The objective of the present paper is to outline the main conclusions derived from fundamental and applied research and to discuss the opportunities for modifying grazing pattern by dairy cows and the implications for dairy-farm feeding strategies. In addition, implications for future research agenda on pastoral systems will be addressed.
Short-term intake rate (IR)
Of the more commonly used descriptors of sward state, bite mass is generally significantly correlated with SSH and green leaf mass; more so than with total herbage mass (Orr et al. 2004). However, SSH is the more easily determined and is more often used to demonstrate the relationships among sward state, bite mass, bite rate and IR (Mezzalira et al. 2014). Research conducted with lactating Friesian cows grazing perennial ryegrass swards maintained by continuous variable stocking management at different overall mean SSH showed responses in bite mass, bite rate and IR (Gibb et al. 1999). Bite mass was largely constrained by sward conditions, whereby when SSH was reduced, bite mass declined. In response to reduced bite mass, cows increased their bite rate, as a compensatory mechanism facilitated by a reduction in the proportion of grazing jaw movements required for manipulation and mastication of the herbage harvested within the smaller bites. Nevertheless, despite such increases in bite rate, only partial compensation was achieved, and IR (the product of bite mass and bite rate) declined with reduced SSH. The primary response by cows to constraints imposed by reduced SSH was to increase the total time spent grazing each day. However, even by increasing their grazing time, cows were unable to fully compensate for the reduction in IR, and daily intake of herbage was reduced on shorter swards.
While SSH is a major determinant of bite mass, and in turn bite rate and IR, these relationships can be affected by several plant and animal factors. Not surprisingly, the bulk density of herbage within swards can have a significant impact of bite mass. Mayne et al. (1997) reported a significant effect of herbage bulk density, as well as SSH, on bite mass and hourly herbage DM intake rate, although the beneficial effect of greater bulk density on intake rate declined as SSH increased. The contrasting morphology of different pasture species can also affect the relationship between SSH and bite. In an experiment to examine the diurnal patterns in the components of grazing behaviour in sheep grazing perennial ryegrass or white clover swards of initially similar SSH, Orr et al. (1997) found significantly lower bite rates, but significantly greater bite masses and IRs on clover than on ryegrass swards. In a subsequent experiment to examine the effect of modification of sward state during pasture depletion on rotationally stocked perennial ryegrass or white clover pastures, Orr et al. (2004) found bite mass to be better correlated with SSH and green leaf mass on white clover than on perennial ryegrass swards.
The primary animal factor affecting the relationship between SSH and bite mass is physiological state, either long-term (lactating vs dry, parity, body condition score) or short-term (fasting). Gibb et al. (1999) demonstrated the differences between lactating and dry cows in daily herbage organic matter (OM) intake, total grazing time (P < 0.001) and shortterm OM intake rate (P = 0.062). Later, Chilibroste et al. (2012), working with early lactating primiparous dairy cows, showed a linear and significant effect of days in milk (0–60 DIM), both in grazing time and bite rate. Patterson et al. (1998), examining the effect of duration of fasting on grazing behaviour, reported that cows fasted for 6–13 h achieved significantly higher bite masses, bite rate and total DMI over the subsequent 1 h at pasture, than did cows fasted for 1–3 h. The longevity of such increased rates was, however, not examined. In another experiment, Chilibroste et al. (1998) imposed two lengths of fasting before grazing (16 and 2.5 h) and found no impact on bite mass. While this is not consistent with Patterson et al. (1998), who found that bite mass after a 6-h fast was 25% higher than in cows fasted for 1–3 h, Chilibroste et al. (1998) used a grazing-bout duration of 138 min after fasting, whereas Patterson et al. (1998) grazed cows for 60 min. Differences in the length of these grazing sessions on bite mass may support the hypothesis that the effect of fasting on increasing bite mass is a transient event. Soca et al. (1999) also reported longer grazing sessions in fasted cows grazing later in the day. However, in the experiment by Gibb et al. (1999), the greater intake requirement of lactating cows than non-lactating cows had no significant effect on bite mass or IR. The strategy employed by the lactating cows to meet their increased nutrient requirement was to significantly increase the duration of their grazing meals and total daily grazing time. Early lactating primiparous dairy cows showed a significant linear effect of DIM (0– 60DIM) both in grazing time and bite rate (Chilibroste et al. 2012).
Under the changing sward conditions that occur while cows graze short-term paddocks provided under rotational stocking management (12 h to 7 days), similar effects of sward state on bite mass, bite rate and IR might be expected. However, when Barrett et al. (2001) recorded grazing behaviour by dairy cows at 0700 hours, 1100 hours, 1400 hours and 1800 hours following release onto a fresh paddock after morning milking, they found no significant effect of sward depletion on grazing behaviour. In contrast, when dairy cows were stocked for 4 days over a succession of rotationally stocked paddocks, Abrahamse et al. (2008) found that as SSH and herbage mass (kg DM/ha) declined over the 4 days, bite rate and the total number of bites each day showed a significant increase, which was accompanied by a significant linear decrease in milk yield.
Summary of the discussion so far
Progress in understanding the functional relationships between sward characteristics and intake rate and bite dimensions has been significant and relevant. However, most of the information lacks the link with heterogeneous sward structures, herd social relationships (e.g. competition, dominance), physiological state and some management practices such as feeding strategies (supplementation with concentrates and roughage), weather (mud, heat stress) and the interaction among these factors.
Temporal pattern of grazing
Apart from the effect of sward structure and the physiological state of the animals, there appears to be a temporal element affecting grazing behaviour. In pastures maintained under continuous variable stocking management, sward structure remains more or less constant throughout the day. Nevertheless, in dairy cows (Gibb et al. 1998) and sheep (Orr et al. 1997), bite mass, bite rate and IR have been shown to change through the course of the day, even where SSH remains constant. Total daily grazing time is the cumulative outcome of all meals (grazing events). Hence, the diurnal grazing pattern emerges from a series of grazing decisions such as ‘when’ to begin, the intensity (i.e. herbage intake rate), ‘what’ frequency and ‘how’ to distribute the grazing events in time (Gregorini 2012). Circadian patterns are seen in these decisions, which clearly influence the form and rate of substrate supply for digestion, metabolism and growth (Gregorini et al. 2006). This raises the question of what causes the initiation and termination of a grazing event?
Research at EEMAC Research Station in Uruguay with dairy cows given access to pasture for 6–8 h between morning and afternoon milkings and supplemented with concentrate and silage has shown a systematic pattern of grazing. Initially, all cows grazed for ~100 min, followed by a period ruminating and idling. At any point in time during the remainder of the period at pasture, ~50% of the cows could be found grazing. Regarding pasture depletion, over 70% of the pasture consumed was eaten during the first half of the grazing session (Chilibroste et al. 2007). A similar pattern has been described by Dobos et al. (2009) for dairy cows given limited access (1–15 h/day) to graze tropical pasture species. Also, Chilibroste et al. (2012) found a similar temporal pattern of grazing by primiparous dairy cows stocked for 7 days over a succession of rotationally stocked paddocks, with herbage allowances from 7.5 to 30 kg DM/ cow.day.
When access to pasture is not limited, the strategy employed by dairy cows to attempt to compensate for a reduction in IR is to increase total grazing time; the question must be asked as to why cows on moderate or tall swards do not graze for as long as those on short swards, thereby increasing their daily herbage intake. The answer lies in the fact that where greater IRs and daily intakes can be achieved, grazing time is limited by the time requirements to ruminate and idle (i.e. non-grazing and non-ruminating activity). Experiments involving the artificial reduction and increase of rumen fill conducted by Chilibroste et al. (1997, 1998) have shown that when fasting induced high fresh-herbage intake rates, ingestive mastication of the herbage was reduced, resulting in the swallowing of larger particles with high water content. When eating fresh grass cows did not show evidence of having problems in accommodating large volumes of material in the rumen, but they failed to pack it properly creating a filling effect even though the DM and neutral detergent fibre rumen digesta pool was low. Gregorini et al. (2006) hypothesised that the circadian release of two interdependent hormones (melatonin and serotonin) may shape the diurnal grazing pattern of ruminants. Also, Gregorini (2012) pointed out that high concentrations of insulin have been observed in grazing dairy cows during the dusk grazing event. High concentrations of insulin weaken the action of glucagon and reduce gluconeogenesis, thereby delaying satiation. Reductions in plasma glucose also induce the release of ghrelin, and the highest concentrations of ghrelin occur before the dawn and dusk grazing events. Ghrelin is a powerful hormone stimulating intake (Roche et al. 2008) and has been related to feeding behaviour (Gregorini et al. 2009a, 2009b).
Having evolved as herd animals, even modern domesticated ruminants exhibit atavistic behaviour patterns. Both sheep and cattle show considerable group synchronicity in the initiation of grazing activity, although slightly less so in terminating this activity (Rook and Penning 1991; Rook and Huckle 1995). Sheep kept in small groups (<4 individuals) spend less time grazing and had reduced intakes compared with sheep in larger groups, possibly because of an atavistic fear of predation (Penning et al. 1993). Ruminants generally exhibit a cyclical peak in grazing activity at ~8-h intervals, as reported by Champion et al. (1994) and Somparn et al. (2007), which has been postulated to represent the optimal cycle duration for maintaining rumen function (Phillips 1992). However, in dairy cows, due to external influences such as removal for milking and darkness, some deviation from a regular 8-h cycle is frequently observed.
The accumulated evidence of the longest grazing meal and highest IRs occurring during the late afternoon and early evening when plant non-structural carbohydrate content was highest prompted various investigations of whether under daily rotational stocking management, moving cows to new daily paddocks following afternoon milking, rather than after morning milking, might benefit animal production. The results of such studies have, however, been equivocal. Offering the same daily herbage allowance, Orr et al. (2001) moved cows to fresh paddocks following either morning or afternoon milking. Time of allocation had no effect on total daily grazing time, but had a profound effect on the temporal pattern of meals. Cows given access to their new paddock in the afternoon had grazing meals of more than 4-h duration, similar to the long meals observed previously under continuous variable stocking management. In contrast, cows given access to their new paddock in the early morning had a more fragmented temporal pattern of grazing activity during the afternoon. Although milk yields between the two treatments did not differ significantly, there was a cumulative effect on production amounting to a 5% benefit to the cows moved to their new paddocks in the afternoon. Abrahamse et al. (2009) conducted a similar experiment moving cows to fresh daily paddocks following either morning or afternoon milking. They reported findings similar to those of Orr et al. (2001), with no significant treatment effect on total grazing time. However, cows given their fresh allocation following afternoon milking showed a proportionate shift of their main grazing activity to the late afternoon and evening. Mattiauda et al. (2013) examined the effect of a 4-h grazing session allocated either early morning (0700 hours to 1100 hours) or late morning (1100 hours to 1500 hours). Cows on the early morning treatment grazed for 36 min longer than those on the later treatment, and performed more bites and non-biting grazing jaw movements. There was, however, no significant difference between treatments in the mean bite rates and total time spent ruminating or idling. However, herbage DMI did not differ significantly between the two treatments, which suggests that without any significant difference in bite rate, cows on late morning grazing (1100 hours to 1500 hours) were able to achieve a greater bite mass and higher IR. Despite the lack of significant changes in total daily grazing time or in milk yield, both these experiments demonstrated that lactation performance can be enhanced, albeit moderately so, by the simplest of changes in grazing management.
As stated before, much of the information derived to support the management of the animal–plant interface at the farm level has resulted from short-term, rather than long-term measurements. There are several reasons for this approach. First, few researchers have access either to automatic recording equipment or staff prepared to undertake visual recordings over long periods. Second, on rotationally stocked paddocks, the main focus of attention has been the effect of declining herbage mass on bite mass, bite rate and IR (e.g. Barrett et al. 2001). Third, due to the response in bite mass, bite rate and short-term IR to rapidly changing sward conditions, it is very difficult to obtain sufficient representative measurements of short-term IR from which to calculate daily intake (short-term IR x total grazing time). Thus, while short-term IR can be assessed by measuring weight change (corrected for insensible weight loss) together with bite rate over relatively short periods (~1 h), few researchers have paid any attention to long-term measurements of grazing behaviour. The rapid and continuous development in the area of information and communication technologies should provide opportunities for studies capable of allowing the integration of short- and long-term determinations, and with a high level of precision. This approach is essential so as to improve our knowledge of persistence and consistency of functional responses in time, and to allow the development of robust feeding strategies at dairy farm level.
Extrapolation from short-term to daylength scenarios
Grassland-based livestock production systems require a balance to be achieved between pasture production and sustainability and short- and long-term animal fitness (production, reproduction, health). Irrespective of whether a continuous or rotational stocking system is used to manage grassland-based livestock production, grazing pressure provides the basis by which pasture utilisation and animal intake and production are regulated.
Under continuous variable stocking management, grazing pressure is adjusted either by varying the number of animals or pasture area, the objective being to provide sward conditions conducive to high daily intake rates and animal production. However, the difficulty faced by pasture managers is to regulate grazing pressure so as to provide optimal sward conditions (e.g. tall, dense swards), while, at the same time, avoiding the accumulation of large areas of ungrazed and potentially senescent, low-quality plant material. Having achieved the desired sward condition (e.g. SSH), a balance has to be maintained between herbage production and herbage consumption
Under rotational grazing management, grazing pressure, usually defined in terms of its reciprocal ‘daily herbage allowance’ is recalculated at frequent intervals of 1–7 days. The herbage allowance expressed as weight of herbage (DM or OM) per animal or relative to animal liveweight (LW) is used to calculate the required paddock area after measurement of herbage mass (DM or OM kg/ha) of the pasture. Under such management, however, grazing of the herbage modifies pasture conditions, which become less conducive to the maintenance of high daily IRs. Thus, while this may initially appear straightforward, as under continuous variable stocking management, a compromise has to be made. While a generous herbage allowance reduces the rate of modification of the sward, thereby lessening the constraint on bite mass, and short-term and daily IRs, it is inevitable that utilisation (kg herbage consumed/kg herbage provided) will be low. Under such conditions, unless the remaining un-utilised herbage is removed by additional grazing using other animals or mechanical recovery, animals moved to the pasture during successive rotations will be presented with increasing amounts of senescing herbage.
Herbage allowance
The curvilinear nature of the relationship between herbage allowance and DMI by cattle and sheep has long been recognised (Greenhalgh et al. 1966; Gibb and Treacher 1978), with successive increments in herbage allowance resulting in progressively smaller increases in DMI. In practice, however, notable additional factors affecting the precise relationship between herbage allowance and DMI are the height above which the herbage mass is measured and the production potential of the animals, as illustrated in the review by (Baudracco et al. 2010). The effect of herbage allowance on DMI and milk production has been extensively investigated in mid-lactation dairy cows (e.g. Peyraud et al. 1996; Delagarde and O’Donovan 2005; Baudracco et al. 2010), although most of this research did not include measurements of grazing behaviour. In earlier studies (Chilibroste et al. 2007), we have examined adaptive changes in grazing behaviour of lactating dairy cows under contrasting feeding strategies involving groups of primiparous and multiparous dairy cows that were past peak milk production (i.e. over 60 DIM). However, due to the scarcity of investigations of the effects of daily herbage allowance on productive performance and feeding behaviour in early lactation primiparous cows, there is a critical deficiency regarding the impact of DMI during early lactation on subsequent animal performance and metabolism.
More recent research by Chilibroste et al. (2012) and Meikle et al. (2013) has sought to address this knowledge deficiency and has examined primiparous grazing dairy cows, integrating measurements of ingestion, production and metabolism. Three groups of 11 cows were allowed to graze on a mixed sward of tall fescue (Festuca arundinacea), bird’sfoot trefoil (Lotus corniculatus) and white clover (Trifolium repens) between 0800 hours and 1500 hours daily over 7 days. Paddock areas for the three groups were 1.0, 0.5 and 0.25 ha, thereby providing herbage allowances of 36.5, 17.2 and 9.1 kg DM/cow.day. Behavioural measurements were made every day during the third, fifth and seventh 7-day grazing rotation. On behaviour-measurement days, cows were observed at 15-min intervals to record their activity. In addition, on three alternate days, bite rates were determined visually at 15-min intervals over 1-h periods beginning at 0800 hours, 1030 hours and 1330 hours, nominally initial, middle and late sessions, respectively. The probability of cows grazing increased significantly from Week 3 to Weeks 5 and 7 of the rotation, and over the 7 days within each rotation. However, there were no significant treatment effects or treatment by week interactions. On the high, medium and low allowances, the mean probability of cows grazing throughout the 7 h at pasture was 61.9%, 63.8% and 63.2%, respectively. Although the probability of cows grazing within the initial session was significantly (P < 0.01) higher than that in the middle and late sessions, it did not differ significantly among the high, medium and low allowances (91%, 86% and 92%, respectively). Nevertheless, in the final session, the probability of grazing tended to be higher for cows on the low allowance than for those on the medium and high allowances (P < 0.07), namely, 62% compared with 53% and 52%, respectively. There was a significant (P < 0.01) effect of allowance on mean bite rates, which were 46.6, 49.5 and 43.4 bites/min for the high-, mediumand low-allowance cows, respectively. Similar to the probability of grazing, bite rate increased during the experiment, with the lowest values in Week 3 (39.8 bites/min) and the highest in Week 8 (60 bites/min). Although the treatment groups did not differ in mean grazing time during the experiment, the change in this variable over time suggests that cows on all treatments did not graze for as long during the first weeks of lactation as was reported previously by Chilibroste et al. (2007) and Gibb et al. (1999). The reduction in grazing time associated with low bite rate suggests that actual DMI relies mainly on bite mass, which ultimately is determined by herbage mass, structure and allowance (Gibb 2006). Nevertheless, it is noteworthy that for the three herbage allowances, the probability of cows grazing was always high (86–92%) during the initial measurement session, but declined during the middle and final sessions, thereby highlighting the importance of behavioural changes of cows as adaptations to changes in sward conditions (Gibb 2006). This also suggests that differences in herbage DMI among daily herbage allowance levels must be constrained by mean bite mass rather than to grazing time and/or bite rate because the cows exhibited a similar temporal grazing pattern after calving across the three herbage allowance treatments.
Supplementation
Another common practice at farm level to reach equilibrium between short-term and long-term requirements of both pasture and animal has been the use of supplements. The interaction among herbage allowance, supplementation and breed has been recently reviewed by Baudracco et al. (2010). However, few studies investigating the impact of supplementation on animal performance have examined the effects on grazing behaviour. The exception has been the studies carried out by the research group at the Institute of Grassland and Environmental Research, UK
During an experiment to examine the effect of SSH on grazing behaviour (Gibb et al. 1997), cows were withdrawn from the pasture for milking at 0500 hours and 1430 hours and returned at 0630 hours and 1600 hours, respectively. In three subsequent experiments to examine the impact of level, composition and temporal pattern of concentrate supplementation on grazing behaviour (Gibb et al. 2000, 2002a, 2002b), cows were stocked on perennial ryegrass swards maintained at a SSH of 7–8 cm and were similarly absent for milking from 0500 hours to 0630 hours and 1430 hours to 1600 hours. In all four experiments, the temporal pattern of grazing meals during daylight was remarkably similar. Although there was some slight variation, the basic temporal pattern was as follows. The first meal following turnout at 0630 hours very rarely exceeded 2 h in duration. A second meal usually commenced between 1030 hours and 1100 hours and was of slightly longer duration. The longest meal of the day started directly after return to the pasture at 1600 hours and frequently continued without a break for up to 6 h. In a separate experiment, but following similar management practices, the cows exhibited elevated bite rates and bite masses (DM mg/bite), resulting in significantly higher shortterm intakes than those measured during the morning meals.
The occurrence of grazing activity in these experiments during the night was variable, but the cows were generally reluctant to graze during darkness. A possible explanation is that the time requirement for ruminating and idling following the large evening meal precluded grazing. However, this appears doubtful because occasional incidents of group nighttime grazing were recorded. Alternatively, although cattle and buffaloes have good night vision, the cows may have been wary of grazing on the darkest of nights so as to avoid possible injury. Certainly, the occurrence of moonlight has been shown to have a significant effect on night-time grazing in swamp buffaloes over the course of a year (Somparn 2004) and a similar effect was reported in a subsequent experiment with dairy cows (Gibb et al. 2005).
The conclusion to be drawn from this series of experiments on supplementation is that where cows have access to pasture throughout 24 h, with the exception of removal for milking, supplementation with a pelleted concentrate has little effect on grazing behaviour. Level of concentrate supplementation provided in the parlour does not affect grazing behaviour (Gibb et al. 2002a). Even when cows have access to their concentrate supplement while at pasture, there is sufficient time available (>20 h) not to affect total grazing time (Gibb et al. 2000). The only significant effect on grazing behaviour occurred where a high starch-content diet was offered and cows exhibited a greater number of shorter grazing meals. This experiment also demonstrated that some control over access to concentrate supplements needs to be exercised, because cows will invariably eat concentrates in preference to grazing herbage. However, these findings should be treated with caution, because pelleted concentrates allow high IRs (250–320 g fresh matter/min), allowing rations to be consumed in less than 30 min. Provision of roughage diets, which impose low IRs, could have an impact on grazing behaviour and require investigation as to their effect on grazing behaviour.
Grazing management
In recent years, a growing interest on the manipulation of grazing-session length has emerged in the major grasslandbased dairy production systems (Chilibroste et al. 1997, 2007, 2012; Kristensen et al. 2007; Pérez-Ramírez et al. 2008; Abrahamse et al. 2009; Dobos et al. 2009; Gregorini et al. 2009b; Kennedy et al. 2009; Mattiauda et al. 2013; Soca et al. 2014).
The interest in restricting time at pasture has increased in recent years due to the potential impact on animal performance (Chilibroste et al. 2007), environmental benefits (Gregorini 2012), herbage production and animal energy expenditure (Gregorini et al. 2009b; Dohme-Meier et al. 2014). Gregorini et al. (2009b) demonstrated that dairy cows reacted to restrictions in their time at pasture mainly by changing locomotory behaviour (stride length, rate of walking and distance walked while eating), which led to considerable changes in the area explored while eating. Dohme-Meier et al. (2014) showed that grazing cows spent proportionally more time walking and less time standing and lying than did zerograzing cows. Within a 6-h measurement period, grazing cows expended 19% more energy than zero-grazing cows and differences in energy expenditure did not change with increasing DIM. The proportion of time spent eating was greater and that of time spent ruminating was lower for grazing than for zero-grazing cows. These authors concluded that lower feed intake along with the unchanged milk production indicated that grazing cows mobilised body reserves to cover additional energy requirements which were at least partly caused by more physical activity. Such findings have important implications for new grassland-based dairy farms which are continuously increasing in herd size. Nowadays, 800–1000 cow-grazing herds are not uncommon in Oceania and South America, where they are required to walk long distances (2–6 km/day) and the herbage allowance in the paddock is limited because the main objective is to increase herd yield rather than individual cow yield. In addition to the increased energy expenditure, such systems also have an impact on animal health, welfare and competition between activities (mainly grazing and idling).
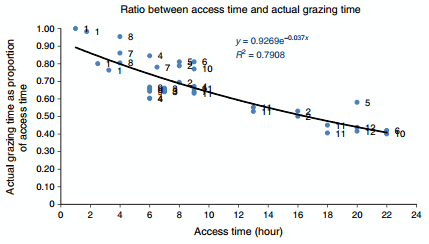
To determine the relationship between the time spent on a paddock and the actual time devoted to grazing, we analysed the data from 12 experiments (Fig. 1).
In Fig. 1, access times shorter than 10 h correspond to one single access to a paddock, typically between two consecutive milkings. The high correlation between access time and grazing time is remarkable, considering that the data are drawn from experiments using different levels of herbage allowance, types and level of supplementation and cows at different stages of lactation. Cows exhibit an exponential decay in terms of their time allocation for grazing activities when access time increases (Fig. 1). Empirical data show that 4–6 h on the paddocks may allow a very efficient grazing session, while longer stays reduce the efficiency with which cows use their time allocation at pasture for grazing. However, this assertion must be considered carefully because Mattiauda et al. (2013) reported a significant reduction in milk production and DMI when supplemented cows were restricted from 8 to 4 h of grazing
Integrating this information with that provided in the previous section concerning potential IRs should form the basis for developing feeding and grazing management strategies for use in modern grassland-based dairy farms. Nevertheless, it must be recognised that these data, although highly valuable, are derived from controlled experiments with a limited number of animals per treatment. To what extend the pattern described in Fig. 1 will be reproduced in large herds (supplemented or not) that walk several kilometres before reaching the grazing paddock, requires further research.
One versus two grazing sessions
Besides the interest in reducing the period of daily access to pastures, a debate has emerged concerning the possible benefits or otherwise of dividing the reduced duration of access into two subperiods (Chilibroste et al. 2007; Gregorini et al. 2009b, Kennedy et al. 2009). Does the grazing management modify actual grazing time? If so, how?
Unfortunately, relatively few reported studies examining the effect of access time on grazing behaviour (e.g. Fig. 1) have been designed to answer these questions. In Table 1, we present data extracted from three experiments where this question can be addressed. In the three experiments cited in Table 1, grazing and ruminating activity were recorded using IGER grazing recorders (Rutter et al. 1997) and interpreted by the software ‘Graze’ (Rutter 2000).
Chilibroste et al. (2007) examined the interaction between herbage allowance and permitted grazing time. Cows were provided with a daily herbage allowance of either 30 or 60 kg/ cow.day, on a mixed legume (45%), grass (19%) and chicory (8%) pasture. On each allowance treatment, two groups of cows were present on the pasture either for 8 h (GT8) from 1800 hours to 0200 hours, or for 16 h (GT16) in two 8-h periods from 1800 hours to 0200 hours and from 0700 hours to 1500 hours. The study was conducted in early spring during mild weather when animal performance was expected to be optimal. Estimated pasture DMI was similar between the treatments, but with the higher producing cows, there were effects of allowed grazing time and sward allowance on milk production and composition. The cows on treatment GT16 grazed for significantly longer than cows on treatment GT8, at both the high herbage allowance (509 vs 333 min) and the low herbage allowance (481 vs 378 min). Although the cows on treatment GT16 were allowed to graze for 8 h longer than cows on GT8, they used only 140 min (or 30%) of the allowed extra time for grazing activities.
Fig. 1. Relationship between the time spent on paddocks and the actual time devoted to grazing activities. Series number refer to the data source: 1, Chilibroste et al. (1997); 2, Chilibroste et al. (2007); 3, Chilibroste et al. (2012); 4, M Fajardo, DA Mattiauda, G Motta, TC Genro, A Meikle, M Carriquiry, P Chilibroste (unpubl. data); 5, Gregorini et al. (2009b); 6, Kennedy et al. (2009); 7, Kristensen et al. (2007); 8, Mattiauda et al. (2013); 9, DA Mattiauda (pers. comm.); 10, PérezRamírez et al. (2008); 11, Soca et al. (2014); 12, Abrahamse et al. (2009).
Table 1. Experiments where the effect of one versus two grazing sessions on grazing time has been evaluated HA, herbage allowance; TMR, total mixed ration
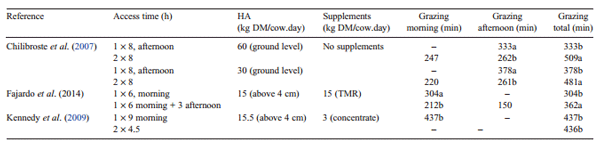
This finding is in accordance with the results of Mattiauda et al. (2004) who examined interactions between permitted grazing time and herbage allowance. Cows were permitted to graze for 8 h (GT8) from 1800 hours to 0200 hours or for a total of 16 h (GT16,) in two periods from 1800 hours to 0200 hours and from 0700 hours to 1500 hours, on pasture strips providing either a high or low herbage allowance (40 or 25 kg/cow.day, respectively). All cows were moved to a fresh pasture strip at 1800 hours, and those on treatment GT16 returned to that same strip at 0700 hours. The experiment was conducted in late spring–early summer when heat stress negatively affected animal performance. Total daily grazing times on the GT8 treatments were the same (397 min), irrespective of the herbage allowance provided. Cows allowed access to pasture for an additional 8 h at the high herbage allowance grazed for an additional 39 min. In contrast, cows allowed access to pasture for an additional 8 h at the low herbage allowance grazed for an additional 87 min. However, irrespective of the herbage allowance, the cows on treatment GT16 exploited only ~30–90 min of their additional 8 h at pasture. The high percentage (83%) of the permitted time at pasture occupied in grazing by the cows on treatment GT8 is probably due to the 16-h fast before grazing (Chilibroste et al. 2007). Such a period of fasting will have induced hunger, which, facilitated by unfouled herbage with a DM mass of 2.6 t/ha, probably induced the cows to achieve a high short-term IR during long uninterrupted grazing bouts, compared with the cows on treatment GT16 that endured only a 3- or 5-h enforced period of fasting. Further analysis of the data from the experiment described by Mattiauda et al. (2004) and Chilibroste et al. (2007) has shown that differences in ingestive behaviour were not observed until the second half of the grazing session at a time when the cows might begin to sense satiety and/or the declining quality of the sward deterred further grazing activity (Barrett et al. 2001).
Kennedy et al. (2009) found that by restricting access to pasture, either in a single 9-h session or two 4.5-h sessions each day reduced total grazing time by 113 min/day, but resulted in cows spending a greater percentage of their time at pasture grazing, than for cows allowed access for 22 h each day (81% vs 42%, respectively). However, permitting the cows either a single 9-h session or two 4.5-h sessions each day had no significant effect on total grazing time, IR or daily herbage DM intake. Unfortunately, Kennedy et al. (2009) did not report differences among treatments through the course of the grazing sessions. The high percentage (81%) of the permitted time at pasture occupied in grazing by the cows in the present study was associated with greater periods of fasting similar to that reported by Chilibroste et al. (2007) and with lower ruminating time.
A study (M Fajardo, DA Mattiauda, G Motta, TC Genro, A Meikle, M Carriquiry, P Chilibroste, unpubl. data), working with early lactation multiparous cows, compared access time to pasture, providing either 6 h of access to pasture in one grazing session and supplemented with 15 kg of a total mixed ration (TMR), or 9 h of access to pasture in two grazing sessions supplemented with the same amount of TMR. Both treatments accessed daily the same herbage DM allowance at pasture and the TMR differed only in the access time to paddocks (i.e. 6 vs 9 h/day, respectively). The 6-h cows had access to the paddock from 0800 hours to 1400 hours and stayed after the afternoon milking in confinement with access to the TMR and water until 0400 hours. The 9-h cows had access to the paddock from 0800 hours to 1400 hours and from 1700 hours to 2000 hours, remaining afterwards in confinement with access to TMR and water until 0400 hours. All cows were milked twice daily, at 0500 hours and 1500 hours. Cows allowed access to pasture for 9 h concentrated their grazing activity between 1700 hours and 2000 hours, rather than in the first 6 h at pasture. This behaviour is in agreement with the diurnal pattern reported by Gibb et al. (1998) where the cows graze for longer and achieve a higher IR in the afternoon than in the morning. Deferment of the major grazing activity to the later session at pasture also benefits the cows by allowing them to ingest herbage with higher DM and water soluble carbohydrate contents (Orr et al. 2001). However, in the experiment reported by Chilibroste et al. (2007), cows were given access to pastures for either one 8-h period (1800 hours to 0200 hours), or two 8-h periods (1800 hours to 0200 hours and 0700 hours to 1500 hours). During the evening grazing session common to both treatments (1800 hours to 0200 hours), cows provided with two 8-h access periods grazed for a shorter time, in more fragmented meals and with more rumination intervals during the first grazing session than did the cows that had a single grazing session. It is remarkable that cows provided a high daily ration of a well balanced supplement (M Fajardo, DA Mattiauda, G Motta, TC Genro, A Meikle, M Carriquiry, P Chilibroste, unpubl. data) consumed their grazing meals following the same temporal pattern as for the cows receiving no supplement, as reported by Chilibroste et al. (2007). A closer look at the behavioural data showed that the differences between treatments start after the first grazing meal in both experiments, which is in line with the observations reported by Gregorini et al. (2009b).
Conclusions
Progress in understanding the functional responses between intake rate and their main components (bite mass, bite area, bite depth and bite rates) has been substantial in the past 25 years. However, progress in understanding the factors that mediate the initiation and the end of individual meals has been more problematic. Evidence of a circadian effect on the modulation of temporal patterns of grazing is recognised and the main hypotheses regarding meal control have focussed on competition between behavioural requirements (grazing vs rumination and idling), physical and chemical signals generated at rumen level and on hunger-related hormones.
Much of the information generated has resulted from shortterm experiments with a limited number of experimental animals and mostly conducted on mono-specific uniform swards. Few studies have undertaken long-term measurements of grazing behaviour and multi-specific, heterogeneous pastures and structures.
In addition, the effect of animals’ long-term (e.g. parity, days in milk) and short-term (e.g. fasting, nutritional signal) physiological state have been investigated little. The energy cost associated with the grazing strategies developed by the animal as a response to different treatments has been almost ignored. These limitations hamper the extension of the functional responses obtained in short-term experiments because they may not be representative of those achieved at a farm level.
More integrated (sward, animal, management) and long-term research is required to have impact on feeding practices at a farm level and on farm design for the new generation of grasslandbased dairy production systems. The increasing availability of new technologies and information communication provides ample opportunities for further development in this area.
References
Abrahamse PA, Dijkstra J, Vlaeminck B, Tamminga S (2008) Frequent allocation of rotationally grazed dairy cows changes grazing behavior and improves productivity. Journal of Dairy Science 91, 2033–2045. doi:10.3168/jds.2007-0579
Abrahamse PA, Tamminga S, Dijkstra J (2009) Effect of daily movement of dairy cattle to fresh grass in morning or afternoon on intake, grazing behaviour, rumen fermentation and milk production. The Journal of Agricultural Science 147, 721–730. doi:10.1017/S0021859 609990153
Barrett PD, Laidlaw AS, Mayne CS, Christie H (2001) Pattern of herbage intake rate and bite dimensions of rotationally grazed dairy cows as sward height declines. Grass and Forage Science 56, 362–373. doi:10.1046/ j.1365-2494.2001.00286.x
Baudracco J, Lopez-Villalobos N, Holmes CW, Macdonald KA (2010) Effects of stocking rate, supplementation, genotype and their interactions on grazing dairy systems: a review. New Zealand Journal of Agricultural Research 53, 109–133. doi:10.1080/00288231003777665
Black JL, Kenney PA (1984) Factors affecting diet selection by sheep. 2. Height and density of pasture. Australian Journal of Agricultural Research 35, 565–578.
Champion RA, Rutter SM, Penning PD, Rook AJ (1994) Temporal variation in grazing behaviour of sheep and the reliability of sampling periods. Applied Animal Behaviour Science 42, 99–108. doi:10.1016/0168-1591 (94)90150-3
Chilibroste P, Tamminga S, Boer H (1997) Effect of length of grazing session, rumen fill and starvation time before grazing on dry matter intake, ingestive behaviour and dry matter rumen pool sizes of grazing lactating dairy cows. Grass and Forage Science 52, 249–257. doi:10.1111/j.1365-2494.1997.tb02355.x
Chilibroste P, Tamminga S, Van Bruchem J, Van der Togt PL (1998) Effect of allowed grazing time, inert rumen bulk and length of starvation before grazing, on the weight, composition and fermentative endproducts of the rumen contents of lactating dairy cows. Grass and Forage Science 53, 146–156. doi:10.1046/j.1365-2494.1998.5320146.x
Chilibroste P, Tamminga S, Boer H, Gibb MJ, den Dikken G (2000) Duration of regrowth of ryegrass (Lolium perenne) swards: effects on grazing behavior, dry matter intake, rumen fill and fermentation of lactating dairy cows. Journal of Dairy Science 83, 984–995. doi:10.3168/jds.S0022-0302(00)74963-0
Chilibroste P, Soca P, Mattiauda DA, Bentancur O, Robinson PH (2007) Short term fasting as a tool to design effective grazing strategies for lactating dairy cattle: a review. Australian Journal of Experimental Agriculture 47, 1075–1084. doi:10.1071/EA06130
Chilibroste P, Mattiauda D, Bentancur O, Soca P, Meikle A (2012) Grazing behaviour of early lactation primiparous dairy cows as affected by herbage allowance: effects on productive performance. Animal Feed Science and Technology 173, 201–209. doi:10.1016/ j.anifeedsci.2012.02.001
Delagarde R, O’Donovan M (2005) Modelling of herbage intake and milk production by grazing dairy cows. Utilization of grazed grass in temperate animal systems. In ‘Proceedings of a satellite workshop of the XXth international grassland congress, Cork, Ireland’. pp. 89–104. (Wageningen Academic Publishers: Wageningen, The Netherlands)
Dobos RC, Fulkerson WJ, Sinclair K, Hinch GN (2009) Grazing behaviour and pattern of intake of dairy cows grazing kikuyu (Pennisetum clandestinum) grass pastures in relation to sward height and length of grazing session. Animal Production Science 49, 233–238. doi:10.1071/ EA08212
Dohme-Meier F, Kaufmann LD, Görs S, Junghans P,MetgesCC, van Dorland HA, Bruckmaier RM, Münger A (2014) Comparison of energy expenditure, eating pattern and physical activity of grazing and zerograzing dairy cows at different time points during lactation. Livestock Science 162, 86–96. doi:10.1016/j.livsci.2014.01.006
Gibb MJ (2006) Grassland management with emphasis on grazing behaviour. In ‘Fresh herbage for dairy cattle: the key to a sustainable food chain’. (Eds A Elgersma, J Dijkstra, S Tamminga) pp. 141–157. (Springer Publishing: Dordrecht, The Netherlands)
Gibb MJ, Treacher TT (1978) The effect of herbage allowance on herbage intake and performance of ewes and their twin lambs grazing perennial ryegrass. Journal of Agricultural Science, Cambridge 90, 139–147. doi:10.1017/S0021859600048681
Gibb MJ, Huckle CA, Nuthall R, Rook AJ (1997) Effect of sward surface height on intake and grazing behaviour by lactating Holstein Friesian cows. Grass and Forage Science 52, 309–321.
Gibb MJ, Huckle CA, Nuthall R (1998) Effect of time of day on grazing behaviour by lactating dairy cows. Grass and Forage Science 53, 41–46. doi:10.1046/j.1365-2494.1998.00102.x
Gibb MJ, Huckle CA, Nuthall R, Rook AJ (1999) The effect of physiological state (lactating or dry) and sward surface height on grazing behaviour and intake by dairy cows. Applied Animal Behaviour Science 63, 269–287. doi:10.1016/S0168-1591(99)00014-3
Gibb MJ, Huckle CA, Nuthall R (2000) Effect of temporal pattern of supplementation on grazing behaviour and herbage intake by dairy cows. In ‘Grazing management: the principles and practice of grazing, for profit and environmental gain, within temperate grassland systems’. British Grassland Society occasional symposium No. 34. (Eds AJ Rook, PD Penning) pp. 91–96. (British Grassland Society: Reading, UK)
Gibb MJ, Huckle CA, Nuthall R (2002a) Effects of level of concentrate supplementation on grazing behaviour and performance by lactating dairy cows grazing continuously stocked grass swards. Animal Science 74, 319–335.
Gibb MJ, Huckle CA, Nuthall R (2002b) Effect of type of supplement offered out of parlour on grazing behaviour and performance by lactating dairy cows grazing continuously stocked grass swards. Animal Science 75, 153–167.
Gibb MJ, Huckle CA, Forbes AB (2005) Effects of sequential treatments with eprinomectin on performance and grazing behaviour in dairy cattle under daily-paddock stocking management. Veterinary Parasitology 133, 79–90. doi:10.1016/j.vetpar.2004.11.038
Greenhalgh JFD, Reid GW, Aitken JN, Florence E (1966) The effects of grazing intensity on herbage consumption and animal production. I. Short-term effects in strip-grazed dairy cows. The Journal of Agricultural Science 67, 13–23. doi:10.1017/S0021859600067526
Gregorini P (2012) Diurnal grazing pattern: its physiological basis and strategic management. Animal Production Science 52, 416–430.
Gregorini P, Eirin M, Refi R, Ursino M, Ansin OA, Gunter SA (2006) Timing of herbage allocation in strip grazing: effects on grazing pattern and performance of beef heifers. Journal of Animal Science 84, 1943–1950. doi:10.2527/jas.2005-537
Gregorini P, Soder KJ, Kensinger RS (2009a) Effect of rumen fill on foraging behavior, intake rate, and plasma ghrelin, serum insulin and glucose levels of cattle grazing a vegetative micro-sward. Journal of Dairy Science 92, 2095–2105. doi:10.3168/jds.2008-1803
Gregorini P, Clark CEF, Jago JG, Glassey CB, McLeod K, Romera AJ (2009b) Restricting time at pasture: effects on dairy cow herbage intake, foraging behavior, hunger-related hormones and metabolite concentration during the first grazing session. Journal of Dairy Science 92, 4572–4580. doi:10.3168/jds.2009-2322
Kennedy E, McEvoy M, Murphy JP, O’Donovan M (2009) Effect of restricted access time to pasture on dairy cow milk production, grazing behaviour, and dry matter intake. Journal of Dairy Science 92, 168–176. doi:10.3168/jds.2008-1091
Kristensen T, Oudshoorn F, Munksgaard L, Søegaard K (2007) Effect of time at pasture combined with restricted indoor feeding on production and behaviour in dairy cows. Animal 1, 439–448. doi:10.1017/S175173 1107694117
Laca EA, Ungar ED, Seligman NG, Ramey MR, Demment MT (1992a) An integrated methodology for studying short-term grazing behaviour of cattle. Grass and Forage Science 47, 81–90. doi:10.1111/j.1365- 2494.1992.tb02250.x
Laca EA, Ungar ED, Seligman NG, Demment MT (1992b) Effects of sward height and bulk density on bite dimensions of cattle grazing homogeneous swards. Grass and Forage Science 47, 91–102. doi:10.1111/j.1365-2494.1992.tb02251.x
Lyons NA, Kerrisk KL, Garcia SC (2014) Milking frequency management in pasture-based automatic milking systems: a review. Livestock Science 159, 102–116. doi:10.1016/j.livsci.2013.11.011
Mattiauda DA, Tamminga S, Elizondo F, Gibb M, Chilibroste P (2004) Effect of allowance and timing grazing session on dairy cows grazing permanent pasture. In ‘II international symposium on grassland ecophysiology and grazing ecology, Curitiba, Brazil’. Session ‘Grazing Systems’, 12th paper. Magnetic support. (Eds A Moraes, E Pizarro, PC Carvalho, S Carneiro da Silva, JR Dittrich) (Universidade Federal do Paraná: Curitiba, Brazil)
Mattiauda DA, Tamminga S, Gibb MJ, Soca P, Bentancur O, Chilibroste P (2013) Restricting access time at pasture and time of grazing allocation for Holstein dairy cows: ingestive behaviour, dry matter intake and milk production. Livestock Science 152, 53–62. doi:10.1016/j.livsci.2012. 12.010
Mayne CS, McGilloway D, Cushnahan A, Laidlaw AS (1997) The effect of sward height and bulk density on herbage intake and grazing behaviour of dairy cows. In ‘Proceedings of the XVIII international grassland congress’. Animal Intake and Grazing Systems, ID no. 1332. pp. 2.15–2.16. (Winnipeg, Manitoba, Canada)
McGilloway DA, Cushnahan A, Laidlaw AS, Mayne CS, Kilpatrick DJ (1999) The relationship between level of sward height reduction in a rotationally grazed sward and short-term intake rates of dairy cows. Grass and Forage Science 54, 116–126. doi:10.1046/j.1365-2494. 1999.00158.x
Meikle A, Adrien L, Mattiauda DA, Chilibroste P (2013) Effect of sward condition on metabolic endocrinology duringthe early postpartum period in primiparous grazing dairycows and its association with productive and reproductive performance. Animal Feed Science and Technology 186, 139–147. doi:10.1016/j.anifeedsci.2013.10.003
Mezzalira JC, De Faccio Carvalho PC, Fonseca L, Bremm C, Cangiano C, Gonda HL, Laca EA (2014) Behavioural mechanisms of intake rate by heifers grazing swards of contrasting structures. Applied Animal Behaviour Science 153, 1–9. doi:10.1016/j.applanim.2013.12.014 Orr RJ,
Penning PD, Harvey A, Champion RA (1997) Diurnal patterns of intake rate by sheep grazing monocultures of ryegrass or white clover. Applied Animal Behaviour Science 52, 65–77. doi:10.1016/S0168-1591 (96)01120-3
Orr RJ, Rutter SM, Penning PD, Rook AJ (2001) Matching grass supply to grazing patterns for dairy cows. Grass and Forage Science 56, 352–361. doi:10.1046/j.1365-2494.2001.00284.x
Orr RJ, Rutter SM, Yarrow NH, Champion RA, Rook AJ (2004) Changes in ingestive behaviour of yearling dairy heifers due to changes in sward state during grazing down of rotationally-stocked ryegrass or white clover pastures. Applied Animal Behaviour Science 87, 205–222. doi:10.1016/j.applanim.2004.01.009
Orr RJ, Young KL, Cook JE, Champion RA (2005) Development of a micro-sward technique for determining intake characteristics of perennial ryegrass varieties. Euphytica 141, 65–73. doi:10.1007/ s10681-005-5365-y
Patterson DM, McGilloway DA, Cushnahan A, Mayne CS, Laidlaw AS (1998) Effect of duration of fasting period on short-term intake rates of lactating dairy cows. Animal Science 66, 299–305.
Penning PD (1986) Some aspects of sward conditions on grazing behaviour and intake by sheep. In ‘Grazing research at northern latitudes’. (Ed. O Gudmundsson) pp. 219–226. (Plenum Publishing Corporation: New York, NY)
Penning PD, Parsons AJ, Newman JA, Orr RJ, Harvey A (1993) The effect of group size on grazing time in sheep. Applied Animal Behaviour Science 37, 101–109. doi:10.1016/0168-1591(93) 90103-V
Pérez-Ramírez E, Delagarde R, Delaby L (2008) Herbage intake and behavioural adaption of grazing dairy cows by restricting time at pasture under two feeding conditions. Animal 2, 1384–1392. doi:10.1017/S1751731108002486
Peyraud JL, Comeron EA, Wade MH, Lemaire G (1996) The effect of daily herbage allowance, herbage mass and animal factors upon herbage intake by grazing dairy cows. Annales de Zootechnie 45, 201–217. doi:10.1051/animres:19960301
Phillips CJC (1992) ‘Farm animals and the environment.’ (CABI Publishing: Wallingford, UK)
Roche JR, Blache D, Kay JK, Miller RD, Sheahan AJ, Miller W (2008) Neuroendocrine and physiological regulation of intake in domesticated ruminants: a review. Nutrition Research Reviews 21, 207–234. doi:10.1017/S0954422408138744
Rook AJ, Huckle CA (1995) Synchronization of ingestive behaviour by grazing dairy cows. Animal Science 60, 25–30.
Rook AJ, Penning PD (1991) Synchronisation of eating, ruminating and idling activity by grazing sheep. Applied Animal Behaviour Science 32, 157–166. doi:10.1016/S0168-1591(05)80039-5
Rutter SM (2000) ‘Graze’: a program to analyse recordings of jaw movements of ruminants. Behavior Research Methods, Instruments, & Computers 32, 86–92. doi:10.3758/BF03200791
Rutter SM (2006) Diet preference for grass and legumes in free-ranging domestic sheep and cattle: current theory and future application. Applied Animal Behaviour Science 97, 17–35. doi:10.1016/j.applanim. 2005.11.016
Rutter SM, Champion RA, Penning PD (1997) An automatic system to record foraging behaviour in free-ranging ruminants. Applied Animal Behaviour Science 54, 185–195. doi:10.1016/S0168-1591(96) 01191-4
Soca P, Chilibroste P, Mattiauda DA (1999) Effect of the moment and length of the grazing session on: 2. Grazing time and ingestive behaviour. In ‘Proceedings of international symposium grassland ecophysiology and grazing ecology’, Agosto 1999, Curitiba, Brasil. (Eds A Moraes, C Nabinger, PC Carvalho, SJ Alvez, SB Lustosa) pp. 295–298.
Soca P, González H, Manterola H, Bruni MA, Mattiauda DA, Chilibroste P, Gregorini P (2014) Effect of restricting time at pasture and concentrate supplementation on herbage intake, grazing behaviour and performance of lactating dairy cows. Livestock Science 170, 35–42. doi:10.1016/j. livsci.2014.07.011
Soder KJ, Orr RJ, Rubano MD, Rook AJ (2009) Use of a micro-sward technique for determining bite mass of four grass species in short-term tests. Euphytica 168, 135–143. doi:10.1007/s10681-009-9907-6
Somparn P (2004) Intensive grazing management strategies for managing swamp buffaloes during thermal stress. PhD Thesis, Kasetsart University, Bangkok, Thailand.
Somparn P, Gibb MJ, Markvichitr K, Chaiyabutr N, Thummabood S, Vajrbukkaa C (2007) Effect of supplementary lighting on eating behaviour in corralled swamp buffalo (Bubalus bubalis) heifer in Thailand. Songklanakarin Journal of Science and Technology 29, 399–411.





.jpg&w=3840&q=75)