The potential of microalgae meals in compound feeds for aquaculture
Published: October 17, 2013
By: Nathan Atkinson, MSc Sustainable Aquaculture Systems student, Fish Nutrition and Aquaculture Health Group, Plymouth University, United Kingdom
Intensive production of mainly carnivorous fish has resulted in fish feeds containing high levels of fishmeal and fish oil, with Europe requiring around 1.9 million tonnes a year. Although this use of fishmeal was initially the recycling of waste from fishing through the use of bycatch and trimmings, due to the rapid development of aquaculture this reliance on fishmeal and fish oil is environmentally unsustainable. This has resulted in other sources of fish feed being investigated. This literature review will focus on microalgae; the composition in terms of nutritional quality, the current methods of production and associated costs along with potential future uses such as feed in aquaculture.
Algae overview
Marine algae are distributed from the polar regions to tropical seas in nutrient rich and poor environments. Algae are photoautotrophs and are characterised by their lack of roots, leaves and presence of chlorophyll a. They range in size from microscopic individual cells called microalgae to seaweeds that can be greater than 30m in length (Qin 2012).
Marine microalgae are the dominant primary producers in aquatic systems and account for a similar level of carbon fixation as terrestrial plants (40-50%) but represent only 1 percent of the planetary photosynthetic biomass (Stephenson 2011). Microalgae are sometimes directly consumed by humans as health supplements due to this high nutritional value and abundance (Dallaire 2007) but this is relatively rare.
As carnivorous fish ingest algae as a food source (Nakagawa 1997) there has been a move to utilise them for fish feed. Currently 30 percent of the world algal production is used for animal feed (Becker 2007) but the use in aquaculture is mainly for larval fish, molluscs and crustaceans (FAO 2009a). As mentioned above the fishmeal and oil use in aquaculture is unsustainable and algae have the potential to reduce this dependence. This is due to the algae being photosynthetic so have the ability to turn the sun’s huge amount of energy, 120,000 TW of radiation, into protein, lipids and nutrients. More energy from the sun hits the surface of the earth in one hour than the energy used in one year and this is a huge amount of untapped, sustainable energy can be exploited by algae. This is a relatively new area of research but has many positive aspects that give it a large amount of potential for future use.
Microalgae
The term ‘microalgae’ is often used to refer specifically to eukaryotic organisms, both from freshwater and marine environments but can include prokaryotes such as cyanobacteria (Stephenson 2011). Microalgal production has received some attention recently due to its potential use as a biofuel (Slocomb 2012), use in animal feed, human consumption and recombinant protein technology (Becker, 2007; Potvin and Zhang 2010; Williams and Laurens, 2010). This has resulted in a huge amount of knowledge and research into microalgae and resulted in reviews being published about specific subjects such as genetic engineering of algae (Qin 2012), potential use as biofuel (Demirbas 2011) and novel methods to measure such important components such as protein (Slocomb 2012).
This interest and knowledge in the area has allowed aquaculture to essentially piggy back the research being performed by the biodiesel industry and even act synergistically with it by consuming the by-products produced (Ju 2012).Currently microalgae have been used in aquaculture as food additives, fishmeal and oil replacement, colouring of salmonids, inducing biological activities and increasing the nutritional value of zooplankton which are fed to fish larvae and fry (Dallaire 2007).
Although the biodiesel industry has been conducting a large amount of research this has mainly been focused towards species that have high lipid contents whereas species in aquaculture must be of appropriate size for ingestion and be readily digested. They must also have rapid growth rates, be able to be cultured on a mass scale, be robust enough to cope with fluctuations in temperature, light and nutrients and have a good nutrient composition (Brown 2002).
Varying nutritional values
The nutritional value of any algal species depends on its cell size, digestibility, production of toxic compounds and biochemical composition. This, along with differences among species and method of production, explains the variability in the amount of protein, lipids and carbohydrates, which are 12-35 percent, 7.2-23 percent, and 4.6-23 percent respectively (FAO 2009a) (Figure 1). This level of fluctuation can be influenced by the culture conditions (Brown et al., 1997) but rapid growth and high lipid production can be achieved by stressing the culture.
Protein
Most of the figures published in the literature on the concentration of algal proteins are based on estimations of crude protein and as other constituents of microalgae such as nucleic acids, amines, glucosamides and cell wall materials which contain nitrogen; this can result in an overestimation of the true protein content (Becker 2007).
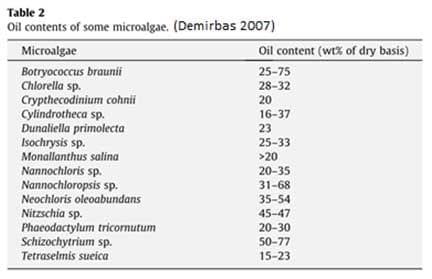
The non-protein nitrogen can be up to 12 percent in Scenedesmus obliquus, 11.5 percent in Spirulina and 6 percent in Dunaliella. Even with this overestimation the nutritional value of the algae is high with the average quality being equal, sometimes even superior, to conventional plant proteins (Becker 2007) (Table 1).
The amino acid composition of the protein is similar between species and is relatively unaffected by the growth phase and light conditions (Brown et al., 1993a, b). Aspartate and glutamate occur in the highest concentrations (7.1-12.9%) whereas cysteine, methionine, tryptophan and histidine occur in the lowest concentrations (0.4-3.2%) with other amino acids ranging from (3.2-13.5%) (Brown 1997).
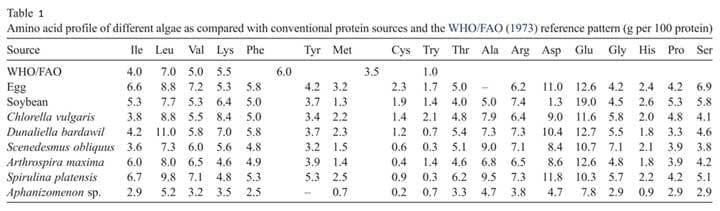
Figure 1: Percentages (dry weight basis) of protein, lipid and carbohydrate in microalgae: The range of values is shown by range bars (Brown 1997)
Lipids
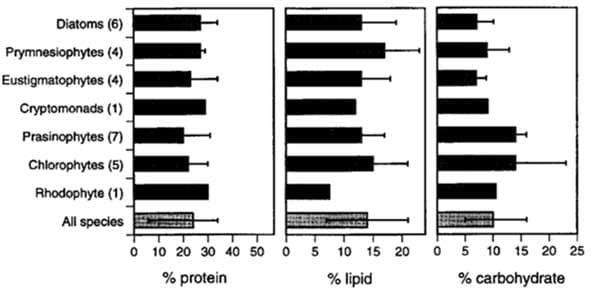
The lipids in microalgal cells have roles as both energy storage molecules and in the formation of biological membranes and can be as high as 70 percent dry weight in some marine species (Stephenson 2011) (Table 2). Under rapid growth conditions these lipid levels can drop to 14-30 percent dry weight, which is a level more appropriate for aquaculture. These lipids are composed of polyunsaturated fatty acids such as docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA) and arachidonic acid (AA) (Brown 2002) and in high concentrations; most species have percentages of EPA from 7-34 percent (Brown 2002) (Figure 2).
These fatty acids are highly sought after and as they currently cannot be synthesised in a laboratory are usually obtained through fish oil and are a limiting factor in vegetable oils such as palm, soybean and rapeseed oil use in aquaculture. The fatty acid composition is associated with light intensity, culture media, temperature and pH. Appropriate measures and control, along with the suitable selection of a species, is necessary to produce algae with the desired lipid level and composition.
Vitamins
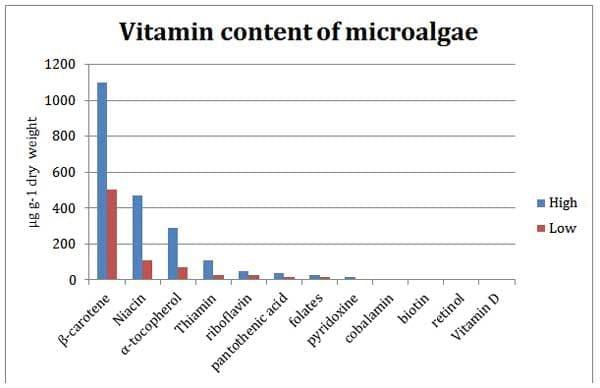
Microalgae also contain vitamins which can be beneficial to the health of the consumer but vary greatly between species (Brown 2002). This variation is greatest for ascorbic acid (Vitamin C), which varies from 1-16mg g-1 dry weight (Brown & Miller, 1992), but other vitamins typically show a 2-4 x difference between species (Brown et al., 1999) (Figure 3).
Despite the variation in vitamin C all the species would provide an adequate supply to cultured animals which are reported to only require 0.03-0.2 mg g-1 of the vitamin in their diet (Durve and Lovell, 1982). However every species of algae had low concentrations of at least one vitamin (De Roeck-Holtzhauer et al., 1991) so a careful selection of a mixed algal diet would be necessary to provide all the vitamins to cultured animals feeding directly on microalgae.
Figure 3: Concentrations of different vitamins in microalgae in µg g-1. Graph adapted from Brown 2002 with data collected from Seguineau et al., 1996 and Brown et al., 1999
Algae in aquaculture
The use of algae as an additive in aquaculture has received a lot of attention due to the positive effect it has on weight gain, increased triglyceride and protein deposition in muscle, improved resistance to disease, decreased nitrogen output into the environment, increased fish digestibility, physiological activity, starvation tolerance and carcass quality (Becker, 2004; Fleurence 2012). Li (2009) showed that the addition of dried microalgae in the diet, albeit at low concentrations 1.0-1.5 percent, resulted in increased weight gain of the channel catfish (Ictalurus punctatus)along with improving the feed efficiency ratio and levels of poly-unsaturated fatty acids. Ganuza (2008) showed that algal oil can be an alternative source of DHA (docosahexaenoic acid) to fish oil in gilthead seabream (Sparus aurata) microdiets although it did not allow for the complete substitution of fisheries products due to the low EPA (eicosapentaenoic acid) levels in the species of algae used.
These were at relatively low-level inclusions; at greater levels it can have a detrimental effect. At 12.5 percent inclusion algae caused reduced growth performances in rainbow trout and at 25 percent and 50 percent this substitution of fish feed caused nutritional deficiencies that led to decreased growth, feed efficiency and body lipids (Dallaire 2007).
Levels of algal inclusion of 15 percent and 30 percent also reduced feed intake and growth rate in Atlantic cod (Walker 2011). As Atlantic cod are known to have a robust digestive system it was suggested that this was due to reduced palatability which could be an issue for algal use in aquaculture.
High levels of inclusion does not cause such negative effects in all species raised in aquaculture, 50 percent replacement did not have a negative effect on shrimp (Ju 2012), but is generally experienced among finfish.
Algae production
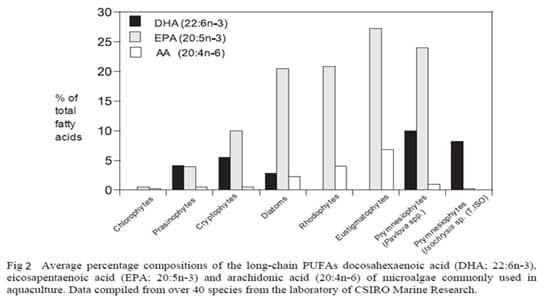
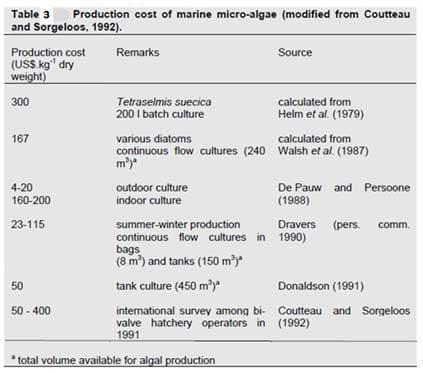
The production of algae, in particular microalgae, is a rapidly developing industry due to the biofuel research that is currently taking place. The annual world production of all species is estimated to be 10,000t year-1 (Richmond, 2004) with the main limit to production currently being the cost. Production costs are currently range from US$4-300 per kg dry weight (FAO 2009a) depending on the type of production method employed (Table 3).
There has been a shift away from typical systems such as outdoor ponds and raceways to large-scale photobioreactors which have a much higher surface area to volume ratio and could potentially reduce the production cost (Brown 2002). These photobioreactors could yield 19,000 - 57,000 litres of microalgal oil per acre per year, which is over 200 times the yield from the best performing vegetable oils (Chisti 2007), and reduce the cost of algal oil from $1.81 to $1.40 per litre (Demirbas 2011).
However, for algal oil to be competitive with petrodiesel, it should be less than $0.48 per litre. This is achievable through economies of scale (Demirbas 2011) and would make it a cheap and sustainable oil for the aquaculture industry. There are also other developments such as increasing the specific activity of the enzyme RUBISCO which would increase yields, transgenic studies, increasing the proportion of photo protective pigments which would improve the light-dependant reactions and selecting for algae with small antennas which is fundamental to achieving high yields in biomass dense cultures (Stephenson 2011). This research is essential as the production costs of microalgae are still too high to compete with traditional protein sources for aquaculture (Becker 2007).
Benefits and obstacles
Algae have a great potential for use in sustainable aquaculture as they are not only a source of protein, lipids and have other nutritional qualities but they are phototrophic so produce these directly from sunlight. Producing 100 tons of algal biomass also fixes roughly 183 tons of carbon dioxide which has obvious implications in this period of climate change.
The production does not always require freshwater, compete for fertile land and are not nutritionally imbalanced with regard to the amino acid content like soybean.
There are still some obstacles such as the powder-like consistency of the dried biomass and applications to feed manufacture, the production costs and pests and pathogens that will effect large scale algal cultivation sustainability (Hannon et al., 2010), which is an area that little is known about.
There still needs to be many feeding trials as the majority of research has focused on improving the nutritional value of rotifers and not as algae as a potential replacement of fishmeal and fish oil. There is also interest into storing algal pastes which have extended shelf life (2-8 weeks) or the use of defatted microalgae meal from the biodiesel industry.
The use of algae in aquaculture is a promising and young area of research and when compared to agriculture, which has increased crop productivity by 138 percent in a 50 year period, it demonstrates the great potential that algae has.
References
Andrews, J.T. and Whitney, S.M. (2003) Manipulatingribulosebisphosphate carboxylase/oxygenase in the chloroplasts of higher plants. Arch. Biochem. Biophys. 414, 159–169
Becker, E. W. (2007). Microalgae as a source of protein.Biotechnology advances, 25(2), 207–10. doi:10.1016/j.biotechadv.2006.11.002
Becker, E.W., 2004. Microalgae in human and animal nutrition. In: Richmond, A. (Ed.), Handbook of Microalgae Culture. Biotechnology and Applied Phycology. Blackwell Science, Oxford, pp. 312–351
Becker EW. Microalgae - Biotechnology and Microbiology. Cambridge: Cambridge University Press; 1994
Ben-Amotz, A., Tomabene, T.G. and Thomas, W.H., 1985.Chemical profile of selected species of microalgae with emphasis on lipids..I. Phycol., 21: 72-81
Brown, M. R., Miller, K. A., 1992. The ascorbic acid content of eleven species of microalgae used in mariculture. Journal of Applied Phycology, 4: 205-215.
Brown, M. R., Dunstan, G. A., Jeffrey, S. W., Volkman, J. K., Barrett, S. M. and LeRoi, J. M. (1993a).The influence of irradiance on the biochemical composition of the prymnesiophyteIsochrysis sp. (clone T-ISO). Journal of Phycology, 29: 601-612.
Brown, M. R., Garland, C. D., Jeffrey, S. W., Jameson, I. D., Leroi, J. M., 1993b. The gross and amino acid compositions of batch and semi-continuous cultures of Isochrysis sp. (clone T.ISO), Pavlovalutheri and Nannochloropsisoculata. Journal of Applied Phycology, 5: 285-296.
Brown, M. R., Jeffrey, S. W., Volkman, J. K., Dunstan, G. A., 1997.Nutritional properties of microalgae for mariculture. Aquaculture, 151: 315-331.
Brown, M. R., Mular, M., Miller, I., Trenerry, C., Farmer, C., 1999.The vitamin content of microalgae used in aquaculture. Journal of Applied Phycology, 11: 247-255.
Brown, M. R., 2002. Nutritional value of microalgae for aquculture. In: Cruz-Suárez, L. E., Ricque-Marie, D., Tapia-Salazar, M., Gaxiola-Cortés, M. G., Simoes, N. (Eds.). Avances en NutriciónAcuícola VI.Memorias del VI Simposium Internacional de Nutrición Acuícola. 3 al 6 de Septiembre del 2002. Cancún, Quintana Roo, México.
Chisti Y. Biodiesel frommicroalgae. BiotechnolAdv 2007;25:294–306.
Couteau, P., Sorgeloos, P., 1992. The use of algal substitutes and the requirement for live algae in the hatchery and nursery rearing of bivalve molluscs: an international survey. Journal of Shellfish Research, 11: 467-476.
Darcy-Vrillon, B. (1993). Nutritional aspects of the developing use of marine macroalgae for the human food industry. International Journal of Food Sciences and Nutrition, 44,23e35.
Dallaire, V., Lessard, P., Vandenberg, G., & De laNoüe, J. (2007). Effect of algal incorporation on growth, survival and carcass composition of rainbow trout (Oncorhynchus mykiss) fry.Bioresource technology, 98(7), 1433–9. doi:10.1016/j.biortech.2006.05.043
Demirbas, A., &FatihDemirbas, M. (2011). Importance of algae oil as a source of biodiesel.Energy Conversion and Management, 52(1), 163–170. doi:10.1016/j.enconman.2010.06.055
Denis, C., Masse, A., Fleurence, J., & Jaouen, P. (2010). Concentration and pre purification with ultrafiltration of a R-phycoerythrin solution extractedfrom macro-algaeGrateloupiaturuturu: processdefinition and up-scaling. Separation and Purification Technology, 69,37e42.
De Roeck-Holtzhauer, Y., Quere, I. and Claire, C., 1991. Vitaminanalysis of five planktonicmicroalgae and one macroalga. J. Appl. Phycol., 3: 259-264.
Durve, V.S. and Lovell, R.T. 1982. Vitamin C and disease resistance in channel cattish (Ictaluruspunctatus). Can. J. Fish. Aquat. Sci. 39: 948-951.
Enright, C. T., Newkirk, G. F., Craigie, J. S., Castell, J. D., 1986. Growth of juvenile Ostreaedulis L. fed ChaetoceroscalcitransSchütt of varied chemical composition Journal of Experimental Marine Biology and Ecology, 96: 15–26.
Falkowski PG, Barber RT, Smetacek V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998;281:200–6.
FAO. 2009b. Fisheries Department, Fishery Information, Data and Statistics Unit.Fishstat Plus: Universal software for fishery statistical time series. Version 2006-11-02. Release data March2, 2006 (available at: www.fao.org/fi/statist/FISOFT/FISHPLUS.asp).
FAO 2009amacrophytes as feed in small-scale aquaculture for link see: Use of algae and aquatic macrophytes as feed in small-scale aquaculture http://www.fao.org/docrep/012/i1141e/i1141e.pdf
Fleurence, J., Morançais, M., Dumay, J., Decottignies, P., Turpin, V., Munier, M., Garcia-Bueno, N., et al. (2012). What are the prospects for using seaweed in human nutrition and for marine animals raised through aquaculture? Trends in Food Science & Technology, 27(1), 57–61. doi:10.1016/j.tifs.2012.03.004
Fleurence J. Seaweed proteins: biochemical, nutri- tional aspects and potential uses. Trends Food Sci Technol. 1999a;10:25–38.
Fleurence J. The enzymatic degradation of algal cell walls: a useful approach for improving protein ac- cessibility? J ApplPhycol. 1999b;11:313–314.
Ganuza, E., Benítez-Santana, T., Atalah, E., Vega-Orellana, O., Ganga, R., &Izquierdo, M. S. (2008).Crypthecodiniumcohnii and Schizochytrium sp. as potential substitutes to fisheries-derived oils from seabream (Sparusaurata) microdiets.Aquaculture, 277(1-2), 109–116. doi:10.1016/j.aquaculture.2008.02.005
Guckert, J.B. and Cooksey, K.E., 1990. Triglyceride accumulation and fatty acid profile changes in Chlorella (Chlorophyta) during high pH-induced cell cycle inhibition. J. Phycol., 26: 72-79
Hannon M, Gimpel J, Tran M, Rasala B, Mayfield S. Biofuels from algae: challenges and potential. Biofuels 2010;1:763–84
Horie, Y., Sugase, K., &Horie, K. (1995). Physiological difference of soluble and insoluble dietary fibre fractions of brown algae and mushrooms in pepsin activity in vitro and protein digestibility. Asian Pacific Journal of Clinical Nutrition, 4, 251e255
Huntington, T. (2009). Use of wild fish and other aquatic organisms as feed in aquaculture – a review of practices and implications in Europe 1, (518), 209–268.
Jeffrey, S. W., LeRoi, J-M., Brown, M. R., 1992. Characteristics of microalgal species for Australian mariculture. In: G. L. Allan and W. Dall (Editors), Proceedings of the National Aquaculture Workshops, Pt. Stephens, NSW Australia, April 1991, pp. 164-173.
Jensen, A. (1993). Present and future needs for algae and algal products.Hydrobiologia, 260/261,15e23.
Ju, Z. Y., Deng, D.-F., &Dominy, W. (2012). A defatted microalgae (Haematococcuspluvialis) meal as a protein ingredient to partially replace fishmeal in diets of Pacific white shrimp (Litopenaeus vannamei, Boone, 1931).Aquaculture, 354-355, 50–55. doi:10.1016/j.aquaculture.2012.04.028
Kawamura, T., Roberts, R. D., Nicholson, C. M., 1988. Factors affecting the food value of diatom strains for post-larval abalone Haliotis iris. Aquaculture, 160: 81-88
Khozin-Goldberg, I. and Cohen, Z. (2006) The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyteMonodussubterraneus.Phytochemistry 67, 696–701
Li, M. H., Robinson, E. H., Tucker, C. S., Manning, B. B., &Khoo, L. (2009). Effects of dried algae Schizochytrium sp., a rich source of docosahexaenoic acid, on growth, fatty acid composition, and sensory quality of channel catfish Ictaluruspunctatus. Aquaculture, 292(3-4), 232–236. doi:10.1016/j.aquaculture.2009.04.033
Macartain, P., Gill, C. I. R., Brooks, M., Campbell, R., & Rowland, I. R. (2007). Nutritional Value of Edible Seaweeds, 2007(December), 535–543. doi:10.1301/nr.2007.dec.535
Marrion O, Fleurence J, Schwertz A, et al. Evaluation of protein in-vitro digestibility of Palmaria pal- mata and Gracilariaverrucosa. J ApplPhycol. 2005; 17:99–102.
Marrion O, Schwertz A, Fleurence J, GueantJL, Villaume C. Improvement of the digestibility of the proteins of the redalgaPalmariapalmata by physicalprocesses and fermentation. Nahrung/Food. 2003;47:339–344.
Marsham S, Scott GW, Tobin ML. Comparison of nutritive chemistry of a range of temperate seaweeds. Food Chem. 2007;100:1331–1336.
Masuda, T. et al. (2003) Chlorophyll antenna size adjustments by irradiance in Dunaliellasalina involve coordinate regulation of chlorophyll a oxygenase (CAO) and Lhcb gene expression. Plant Mol. Biol. 51, 757–771
Melis, A. (2009) Solar energy conversion efficiencies in photosynthesis: minimizing the chlorophyll antennae to maximize efficiency. Plant Sci. 177, 272–280
Muller-Feuga, A. 2004. Microalgae for aquaculture. The current global situation and future trends.pp. 352-364. In A. Richmond, ed. Handbook of Microalgal Culture. Oxford, Blackwell.
Mustafa, M.G. and Nakagawa, H. (1995) ‘A review: dietary benefits of algae as an additive in fish feed’ in The Isreali Journal of Aquaculture 47, 155±162
Mustafa, M.G., Wakamatsu, S., Takeda, T.A., Umino, T. and Nakagawa, H. (1995) ‘Effects of algae meal as feed additive on growth, feed efficiency, and body composition in Red Sea Bream’ in Fisheries Sciences 61, 25±28
Nakagawa, H. (1997). Effect of dietary algae on improvement H Nakagawa of lipid metabolism in fish mg FA / g g, 345–348.
Potvin, G., Zhang, Z., 2010. Strategies for high-level recombinant protein expression in transgenic microalgae: a review. Biotechnol. Adv. 28, 910–918.
Qin, S., Lin, H., & Jiang, P. (2012). Advances in genetic engineering of marine algae.Biotechnology advances, 30(6), 1602–13. doi:10.1016/j.biotechadv.2012.05.004
Richmond A. Handbook of Microalgal Culture. Biotechnology and Applied Phycology. Oxford: Blackwell Science; 2004.
Sanchez-Machado DI, Lopez-Hernandez J, Paseiro-Losada P, Lopez-Cervantes J. Fatty acids, total lipid, protein and ash contents of processed edible seaweeds. Food Chem. 2004;85:439–444.
Satoh, K.I., Nakagawa, H. and Kashara, S. (1987) ‘Effect of Ulva meal supplementation on Disease resistance of Red Sea Bream' in Nippon Suisan Gakkaishi 53, 1115±1120
Seguineau, C., Laschi-Loquerie, A., Moal, J., Samain, J. F., 1996. Vitamin requirements in great scallop larvae.Aquaculture International, 4: 315–324.
Shigueno, K. and Itoh, S., 1988. Use of Mg-L-ascorbyl-2-phosphate as a vitamin C source in shrimp diets. J. World Aquacult. Sot., 19: 168-174.
Slocomb, S. P., Ross, M., Thomas, N., McNeill, S., & Stanley, M. S. (2012).A rapid and general method for measurement of protein in micro–algal biomass.Bioresource Technology, 129, 51–57. doi:10.1016/j.biortech.2012.10.163
Stephenson, P. G., Moore, C. M., Terry, M. J., Zubkov, M. V, &Bibby, T. S. (2011). Improving photosynthesis for algal biofuels: toward a green revolution. Trends in biotechnology, 29(12), 615–23. doi:10.1016/j.tibtech.2011.06.005
Thompson, P. A., Guo, M.-X., Harrison, P. J., 1993. The influence of irradiance on the biochemical composition of three phytoplankton species and their nutritional value for larvae of the Pacific oyster (Crassostreagigas). Marine Biology, 117: 259-268.
Thompson, P.A., Harrison, P.J., 1992. Effects of monospecific algal diets of varying biochemical composition on the growth and survival of Pacific oyster Crassostreagigas larvae. Mar. Biol. 113, 564–645
Thompson, P.A., Harrison, P.J. and Whyte, J.N.C., 1990. Influence of irradiance on the fatty acid compositionof phytoplankton. J. Phycol., 26: 278-288.
Walker, Abigail B. Berlinsky, David L (2011). Effects of Partial Replacement of Fishmeal Protein by Microalgae on Growth, Feed Intake, and Body Composition of Atlantic Cod. North American Journal of Aquaculture Vol. 73, Iss. 1,
Webb, K. L., Chu, F. E., 1983. Phytoplankton as a food source for bivalve larvae. In: G. D. Pruder, C. J. Langdon and D. E. Conklin (Editors), Proceedings of the Second International Conference on Aquaculture Nutrition: Biochemical and Physiological Approaches to Shellfish Nutrition, Louisiana State University, Baton Rouge, LA, pp. 272–291.
USDE 2005 US Department of Energy Report on the basic energy science workshop on solar energy utilization. For link see: http://science.energy.gov/~/media/bes/pdf/reports/files/seu_rpt.pdf
Williams, P.J.leB., Laurens, L.M., 2010. Microalgae as biodiesel and biomass feedstocks: reviews and analysis of the biochemistry, energetics and economics. Energy Environ. Sci. 3, 554–590.
Related topics
Recommend
Comment
Share

31 de octubre de 2013
Hi Nathan, this is a very intesting topic and very well done for this fine article!
One (minor? :) point: when assessing nutritional value, I would suggest that we need to examine other lipid compounds (and not only EPA, DHA), see
1. http://onlinelibrary.wiley.com/doi/10.1111/1750-3841.12194/abstract and
2. http://www.mdpi.com/1660-3397/11/10/3676
Best,
Yannis
Recommend
Reply

Would you like to discuss another topic? Create a new post to engage with experts in the community.

