Influence of dietary Bio-Mos® on rainbow trout and common carp
Influence of dietary Bio-Mos® on growth, survival and immune status of rainbow trout (Salmo gairdneri irideus G.) and common carp (Cyprinus carpio L.)
The success of intensive aquaculture has been the direct result of improved genetics, nutrition and management as well as maximising disease control. A commercial production system can only be economically viable if these factors are optimised. Key zootechnical parameters to evaluate the efficacy of a system include growth rate, feed conversion ratio (FCR) and mortality, which are influenced primarily by the quality of feed and feed intake.
For many years water quality has been the most important limitation to fish production, however substantial advances in recent years have alleviated this, and nutrition is now regarded the key limitation to increased production efficiency as well as the growth and propagation of new aquaculture species.
Interest in fish nutrition has increased markedly over the past two decades, largely due to a global increase in aquaculture production (Lall, 2000). In recent years much of the research has focused on meeting the nutritional requirements of the fish and the benefits of supplementing feed with pro-nutrients (Spring, 1999).
Pro-nutrients are not essential to the fish; however, they can benefit animal health and performance through improving availability or utilization of nutrients in a variety of ways.
This paper discusses immune defence in fish and describes a series of trials investigating the effect of the pro-nutrient mannan oligosaccharide (Bio-Mos®, Alltech Inc.) on performance and the non-specific (innate) immune system of rainbow trout and common carp.
Trout and carp: important species in Bulgarian aquaculture
Rainbow trout, (Salmo gairdneri irideus G.) is one of the most important species commercially grown in Bulgaria, and fish are raised in either raceways or net cages suspended in lakes and artificial dams.
Commercially prepared diets for salmonids have been developed based on numerous research studies evaluating the specific nutritional requirements of these species, their production systems and their life stages.
Common carp (Cyprinus carpio L.) is an important commercially produced fish all over the world. In many countries rearing of carp species is conducted using extensive and semi-intensive production technologies.
During the last few decades Bulgarian fish farmers started rearing common carp in intensive systems.
Market demand is for a 1.5 kg 2-year old common carp; and in order to achieve these weights, more intensive methods are required. Farmers are increasingly looking to alternative natural strategies to support good levels of growth and performance, as well as maintaining health and survival whilst reducing reliance on antibiotics and chemical growth enhancers.
Immunity in fish
Fish are constantly under threat from a large number of pathogens consisting of bacteria, viruses, fungi, and parasites (Iwama and Nakanishi, 1996). The immune system is an intricate and complex system that protects fish from disease and prevents the spread of infection.
Immune systems of most fish are very similar (Zelikoff et al., 2000), and have certain similarities to mammals (Zelikoff, 1998; Beaman et. al., 1999). The non-specific immune system consists of several components that provide innate protection against infection, regardless of the pathogen type (Sunyer and Lambris, 1998).
Lysozyme, one component of innate immunity, destroys bacteria via hydrolase activity, which breaks down ß- (1-4) linkages in the peptidoglycan layer of bacterial cell walls. Bacteria are either destroyed directly or are opsonized, enabling destruction through phagocytosis.
While Gram(+) bacteria are more sensitive to lysozyme than Gram(-) bacteria in mammals, fish lysozyme is capable of responding to both Gram(+) and Gram(-) bacteria.
Lysozyme often works simultaneously with other mechanisms of the non-specific immune system, such as complement and macrophage activity (Siwicki et al., 1998). Lysozyme is continuously expressed by neutrophils, monocytes and macrophages, and lysozyme activity tends to increase with stimulation of the immune system. Changes in lysozyme activity are common once a fish becomes infected with a pathogen, depending on the level of infection (Siwicki et al., 1998).
Lysozyme is commonly found in the kidney, spleen, skin mucus, gills, blood plasma, and ovaries of fish. The enzyme may act as a first defence against bacteria through its presence on external surfaces such as on the skin mucus and gills. These defence strategies, together with the internal function of lysozyme within the major organs, contribute greatly to preventing bacterial infection.
Lysozyme activity in fish can vary with genetic, physiological and environmental factors, and increases have been associated with elevations in temperature.
Other environmental conditions such as salinity, dissolved oxygen, food availability, and the existence of harmful pollutants can also influence the lysozyme response (Ingram, 1987). Lysozyme activity has also been found to vary under different conditions of stress. While lysozyme activity is suppressed following a chronic stress, such as long term crowding of fish, it is often elevated after fish are exposed to an acute stress. It has been suggested that elevated lysozyme activity may be more of an indication of stress, rather than resistance to disease when the fish is exposed to acute stress.
Other physiological changes in fish, such as parr-smolt transformations in salmonids, have been found to reduce lysozyme activity. This reduction often coincides with a depression in the number of leukocytes, and is thought to be an indicator of immunosuppression. Lysozyme activity varies among species of fish, genetic strains and different pathogens (Kiczka, 1994).
The complement system is an important element of both the innate and adaptive immune system, and the alternative, lectin and classical pathways have been described in fish (Sakai, 1992; Matsushita et al., 1998; Sunyer and Lambris, 1998). The alternative and lectin pathways are ancient mechanisms present in invertebrates as well as vertebrates. The alternative pathway is activated non-specifically by various microorganisms. The lectin pathway, which has not been much studied in teleosts (Matsushita et al., 1998), is activated by the binding of mannose on the surface of microorganisms to a complex of mannose-binding lectin (MBL) and MBL-associated serine protease (MASP).
All three pathways converge in the lytic pathway, leading to opsonization or direct destruction of the microorganism. The complement system is composed of numerous proteins, and all pathways generate factor C3, which has been described and isolated from teleost species (Nakao and Yano, 1998).
Fish complement is, in many ways, comparable to mammalian complement, but some important differences have been described. In general, the complement system of the alternative pathway is more active in fish than in mammals, is more heat labile and therefore difficult to preserve. Also, fish complement has a lower optimum reaction temperature than its mammalian counterpart, and its components, like C3, are more polymorphic (Nonaka et al., 1981; Sakai, 1983, 1984a, 1984b, 1992; Nakao and Yano, 1998; Sunyer et al., 1997; Sunyer and Lambris, 1998).
IMMUNE SYSTEM MODULATORS
One approach to enhancing fish health is by optimising immune defence with pro-nutrients. Immune modulators improve performance when used to reduce mortality due to disease, and to enhance resistance of farmed species (Raa, 2000). Immunomodulatory compounds can also reduce the negative effects of stress on the immune systems of fish in intensive production systems (Klesius et al., 2000; Siwicki et al., 1998).
Feeding of fish with ß-1,3/1,6-glucan may enhance the production of antibodies against pathogens present in the local environment and thus help to strengthen resistance against pathogens, as well as enhancing nonspecific cellular defence (Raa et al., 1992; Raa, 1996).
Peptidoglycans, ß-glucans and mannans have been shown to have non-specific immunomodulatory effects in several animal species, including fish (Matsuo and Miyazono, 1993; Yoshida et al., 1995).
Supplementation of yeast-derived mannans in the form of Bio-Mos® has been reported to improve the composition of the intestinal microflora in chickens as well as calves, resulting in performance benefits (Spring et al., 2000). Zhou and Li (2004) investigated the effects of dietary Bio-Mos® in aquaculture, and reported an increase in performance of fish. Zhou et al. (in press), based on FCR and weight gain responses of Cyprinus carpio var. Jian, concluded that the optimum dietary inclusion rate of Bio-Mos® was 2.4 g/kg.
The same researchers observed that immune function of carp was optimized at the higher inclusion rate of 5.5 g Bio- Mos®/kg. Hanley et al. (1995) reported that dietary Bio-Mos® might greatly improve survival, biomass recovered and the economics of production of hatcheryreared tilapia. Fisher et al. (2001) reported that the same mannan-based product added at either 2 or 4 g/kg improved immune capacity of the white shrimp Litopenaeus vannamei, cultivated at low or normal salinity.
Reports of improved performance in fish are in agreement with findings in other animal species, where enhanced body weight, survival rate and improved FCR have been reported (Spais et al., 2003; Fritts and Waldroup, 2003; Hooge et al., 2003; Sims et al., 2004; Waldroup et al., 2003).
Bio-Mos® response in rainbow trout and common carp
Bio-Mos® has been shown to enhance immune protection in different animal species and by doing so enhance overall animal performance. However, information in aquaculture is limited. To maximise the effects of Bio- Mos®, factors including timing, dosage, method of administration and physiological condition of fish must be taken into consideration. The purpose of the present study is to evaluate benefits of Bio-Mos® as a supplement at 2 g/kg in pelleted diets on the growth performance, survival and immune function of rainbow trout and common carp.
METHODS
Two trials were conducted in rainbow trout and one with common carp. A net cage trial was carried out at a trout farm (‘Nomicom’) in the Radnevo region of Bulgaria using plastic cages, each measuring 3 x 3 x 2.5 m. Trout (14,400) with initial mean body weights of 30 g, were randomly distributed to eight cages, giving four replicate cages per dietary treatment (Table 1). No adaptation period was imposed. Dietary treatments consisted of pelleted extruded formulations (2.8 mm) containing either no supplement (Control) or Bio-Mos® (2 kg/t). Duration of the experiment was 42 days.
A raceway trial (using older trout with an average weight of 101 g) was conducted at a fish farm (‘Ribena’) in the Zlatna Panega region of Bulgaria. Fish were randomly distributed to eight raceways, giving four replicate raceways per dietary treatment. As in the net cage trial, no adaptation period was imposed (Table 1).
The extruded pelleted (3 mm) diet contained either no supplement (Control) or 2 kg/Bio-Mos®/t. Test diets were fed for a 90 day period.
| Table 1. Crude protein, fat and fibre contents of basal diets used in net cage and raceway trout and common carp trials. |
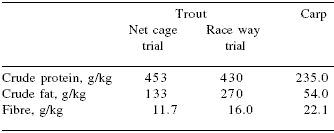 |
For both trout trials rations were supplied in six equal meals per day, fed at 08.00; 10.00; 12.00; 14.00; 16.00 and 18.00 hrs.
The carp trial was conducted at the Nomicom fish farm using an eight cage production system. Four replicate cages measuring 3 m x 3 m x 2.5 m were used per treatment, each housing 675 carp with an average initial weight of 140 g. Trial diets (4 mm extruded pellets) were based on commercial formulas and differed only in the absence (Control) or presence of Bio-Mos® (2 kg/t) (Table 1). The trial lasted for 60 days.
Samples were taken in all trials at the start, mid point and end of each trial period. Samples of 40 fish from each replicate cage or raceway were individually weighed, and body weight, weight gain, mortality and feed intake (calculated FCR) were recorded. In addition, immune parameters of fish were also determined using the following methodologies:
• Serum lysozyme concentration (Lie, 1985),
• Alternative pathway of complement activation (APCA) (Sotirov, 1986),
• Classical pathway of complement activation (CPCA) (Stelzner and Stain, 1971).
All data were subjected to ANOVA with confidence limits set at 95%.
PERFORMANCE AND MORTALITY RESPONSES
Rainbow trout (net cage systems)
Trout raised in net cages were 9% more efficient (P<0.05) when receiving Bio-Mos®-supplemented feed (Figure 1). At the end of the three week trial period, mean body weight of trout recieving Bio-Mos® was 8% higher than controls (P<0.05). At the end of the 42 day trial improved growth rates and higher body weights (+13.7%) were noted for fish given Bio-Mos®- supplemented diets (P<0.001) (Table 2). Total fish weight from the control diet group was 611.44 kg, 15% lower than the Bio-Mos® group (P<0.001) (Figure 2). In addition, mortality was reduced in fish given Bio- Mos® (P<0.001).
| Table 2.Effect of dietary Bio-Mos® on body weight, weight gain and mortality of rainbow trout in a net cage system. |
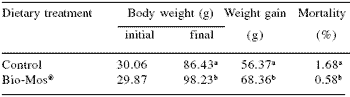 |
| abDifferent superscripts within a column are different (P<0.05) |
Rainbow trout (raceway systems)
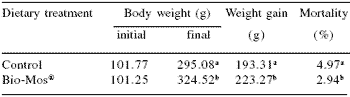
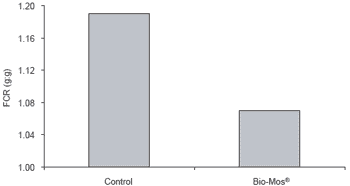
Responses to Bio-Mos® supplementation in fish reared in raceway systems were similar to those noted in the net cage trial. Feed efficiency was improved (P<0.05, Figure 3), and over the 90 day trial period, trout receiving Bio-Mos® gained weight at a 15% higher rate (P<0.05) (Table 3).
Final body weight of fish given Bio-Mos® was 10% higher than those receiving control diets (P<0.01). Mortality of fish in the control group was 41% higher than those given Bio-Mos® (P<0.001).
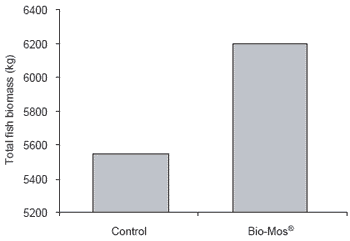
Total fish production from the control group was 12% lower than that from fish receiving Bio-Mos® (P<0.001) (Figure 4).
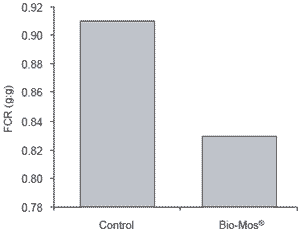 |
| Figure 1.Effect of Bio-Mos® on feed efficiency of trout reared in net cages. |
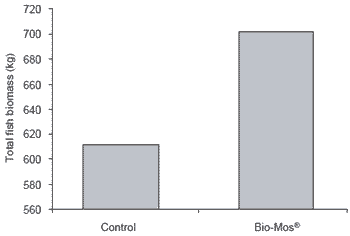 |
| Figure 2.Effect of Bio-Mos® on total trout production from a net-based system. |
| Table 3.Effect of dietary Bio-Mos® on body weight, weight gain and mortality of rainbow trout in a raceway. |
 |
| abDifferent superscripts within a column are different (P<0.05). |
IMMUNE RESPONSES OF RAINBOW TROUT
Rainbow trout (net cage systems)
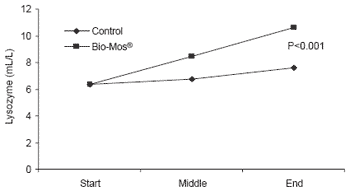
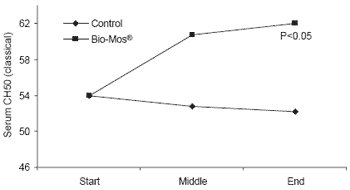
At the end of the 42 day trial, trout given Bio-Mos® had serum lysozyme levels 40% higher than controls (P<0.001) as well as higher complement activity triggered by either the alternative or classical pathways (P<0.05) (Figure 5).
Rainbow trout (raceway systems)
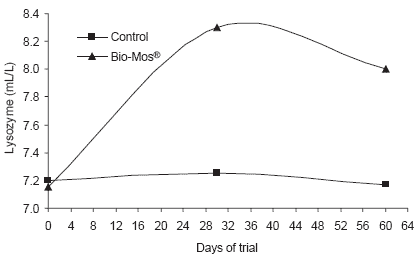
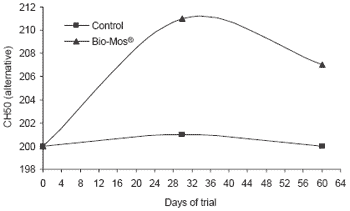
Similar trends in immune response were noted in response to Bio-Mos® in diets fed trout reared in raceways. Lysozyme concentrations in serum were higher at the end of the 90 day trial (+19%, P<0.001) (Figure 6). Likewise, complement activity (alternative and classical pathways) increased in fish given Bio-Mos® over the trial period and was significantly higher at the end of the trial.
PERFORMANCE RESPONSES IN COMMON CARP
Overall, carp receiving Bio-Mos® supplementation had signficantly better weight gain, feed conversion and lower mortality compared to the carp fed control diets (Table 4). Effects of Bio-Mos® on body weight were evident throughout the trial.
After 30 days, carp fed Bio-Mos® had 14% higher body weight than those receiving the control diet (P<0.001). After 60 days, body weight in the Bio-Mos® group was 12% higher than the control (P<0.05) (Figure 7). Overall, supplementation with Bio-Mos® increased weight gain by 17% (P<0.05) for an increase in total fish biomass of 13% (P<0.001) while feed efficiency improved by 17.75% (P<0.01).
 |
| Figure 3.Effects of Bio-Mos® on efficiency of trout reared in raceways. |
 |
| Figure 4. Effect of Bio-Mos® on total trout production from a raceway system. |
 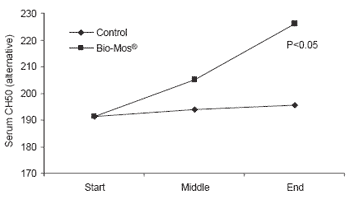 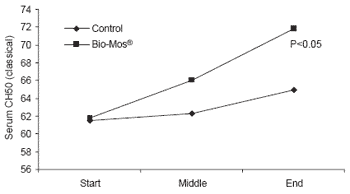 |
| Figure 5.Effect of Bio-Mos® on serum lysozyme and complement activity in trout reared in net cages. |
| Table 4.Effect of dietary supplementation of Bio-Mos® on weight gain, FCR and mortality of common carp. |
 |
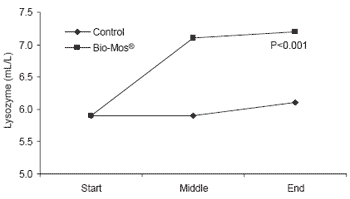 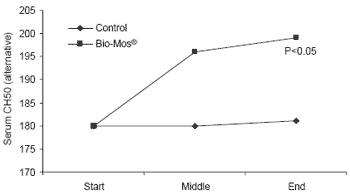  |
| Figure 6.Effect of Bio-Mos® on serum lysozyme and complement activity in trout reared in raceways. |
IMMUNE RESPONSES OF COMMON CARP
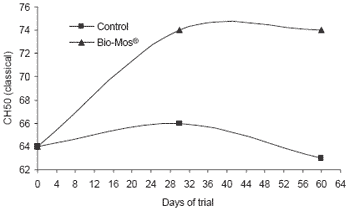
As in the trials with trout, serum lysozyme activity of carp increased during the trial in fish given Bio-Mos® and remained significantly higher at the end of the study (+13%, P<0.001, Figure 8). Likewise, after receiving Bio-Mos® in the diet for 60 days, carp had higher blood complement activity for both pathways (respectively P<0.05 and P<0.001).
Discussion
Advances in fish genetics and management have been substantial in recent years, and nutrition is increasingly becoming a key limitation to production efficiency as well as growth and propagation of new aquaculture species.
In order to maximise performance, the diet must supply essential nutrients in the right forms and concentrations, as well as pro-nutrients to maximizefish health and performance. Optimal nutrition can help fish to overcome health challenges and achieve full genetic potential.
Supplementation of trout and carp diets with 2 g Bio- Mos®/kg improved FCR of reared fish. These results are consistent with those of Hanley et al. (1995), obtained with red tilapia fed 6 g/kg Aqua-Mos in feed where FCR was 27% lower than in controls.
Bio-Mos® had a positive effect on growth rate, weight gain and total biomass production in trout and carp, which concurs with reports of Zhou and Li (2004). Mortality was also reduced, as was previously reported by Hanley et al. (1995) for red tilapia. Enhanced resistance to disease through feeding glucans in aquaculture feeds is thought to increase the survival rate of salmon and trout (Newman, 1994).
As an important component of nonspecific immune response, lysozyme is a useful indicative parameter (Kiczka, 1994; Bayne and Gerwick, 2001). Following antigen stimulation, regulatory and compensatory mechanisms that control lysozyme synthesis are activated (Ralph and Phi-van, 1998; Paulsen et al., 2001).
During the trials with trout and carp, a clear increase in lysozyme concentrations was recorded for fish receiving Bio-Mos® supplementation. Similar effects on the immune function of juvenile Jian Carp were recorded by Zhou and Li (2004) in response to Bio-Mos®-supplemented diets.
The complement system is one of the major mechanisms by which pathogen recognition triggers an effective host defence against infection (Buchmann, 1998). The complement is a system of plasma proteins that can be activated directly or indirectly by pathogens and their bound antibodies (Holand and Lambris, 2002).
Improved immune capacity of fish can be demonstrated by higher complement activity, triggered through both alternative and classical pathways. Bio-Mos® provides mannose attachments, and triggers the complement cascade, activating the immune system (Spring, 2002).
These effects have been described by Raa (2000) who investigated immune stimulants in both fish and shellfish. The results obtained from the recent trout and carp trials show that the levels of complement activity in the blood of fish fed Bio-Mos® increased during the investigation period.
Data from trials with trout and carp clearly suggest that 2 g/kg Bio-Mos® supplement in pellets improved growth rate, FCR, survival and immune response.
Conclusions
|
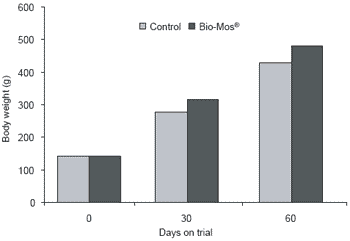 |
Figure 7. Effect of dietary Bio-Mos® on carp body weights at 30 and 60 days. |
|
Figure 8. Effect of Bio-Mos® on serum lysozyme and complement activity in common carp. |
Acknowledgments
These trials could not proceed without the active cooperation of many persons and companies, to whom we are deeply obliged. Our Research Team wishes to thank to Alltech Inc. for their financial support for the project defining the influence of Bio–Mos® as a supplement in pellets on the growth rate and immune status of rainbow trout and common carp.
References
Bayne, C.J. and L. Gerwick. 2001. The acute phase response and innate immunity of fish. Developmental and Comparative Immunology 25:725-743.
Beaman, J.R., H. Gardner, F. Hoffman, A. Rosencrance and J.T. Zelikoff. 1999. Mammalian immunoassays for predicting the toxicity of malathion in a laboratory fish model. J. Toxicol. Environ. Hlth. 56:523-542.
Buchmann K. 1998. Binding and lethal effect of complement from Oncorhynchus mykiss on Gyrodactylus derjavini (Platyhelminthes: Monogenea). Diseases of Aquatic Organisms Vol. 32(3):195-200.
Fischer, A., J. Arias, E. Motte, F. Peralta, V. Cedeno and E. Mialhe. 2001. Effect of Aqua-Mos and SP604 on immune response in the white shrimp (Litopenaeus vannamei). Poster presented at World Aquaculture, Jan. 21-25. Lake Buena Vista, FL, USA.
Fritts, C.A. and P.W. Waldroup. 2003. Evaluation of Bio-Mos® mannan oligosaccharide as a replacement for growth promoting antibiotics in diets of turkeys. Int. J. Poult. Sci. 2(1):19-22.
Hanley F., H. Brown and J. Carbery. 1995. First observations on the effects of mannan oligosaccharide added to hatchery diets for warmwater Hybrid Red Tilapia. Poster presented at Alltech’s 11th Annual Symposium on Biotechnology in the Feed Industry, Lexington, KY, USA.
Holand M.C.H. and J. D. Lambris. 2002 The complement system in teleosts. Fish & Shellfish Immunol. 12:399-420.
Hooge, D.M., M.D. Sims, A.E. Sefton, A. Connolly and P. Spring. 2003. Effect of dietary mannan oligosaccharide, with or without bacitracin or virginiamycin, on live performance of broiler chickens at relatively high stock density on new litter. J. Appl. Poult. Res. 12:461-467.
Ingram, G.A. 1987. Haemolytic activity in the serum of brown trout, Saimo trutta L. J. Fish Biol. 31(Suppl. A):9-17.
Iwama K. and T. Nakanishi. 1996. The fish immune system:organism, pathogen and environment. San Diego, Academic Press, ISBN: 0123504392.
Kiczka W. 1994. From lysozyme monomer to lysozyme dimer. Zycie Weterynaryjne 4A:131-136.
Klesius, P.H., J.J. Evans and C.A. Shoemaker. 2000. Communicable bacterial diseases in fish farming: Prevention by vaccination and immunostimulation. Panamerican Congress of Veterinarian Science.
Lall, S. 2000. Nutrition and health of fish. In: Avances en Nutricion Acuicola V. Memorias del V. Simposium Internacional de Nutricion Acuicola (L.E. Cruz- Suarez, D. Ricque-Marie, M.A. Olvera-Novoa and R. Civera-Cerecedo, eds). 19-22 Noviembre, 2000. Merida,Yucatan, Mexico.
Lie, O. 1985. Improved agar plate assays of bovine lysozyme and haemolytic complement activity. In: Markers of Resistance to Infection in Dairy Cattle. Dissertation. Oslo, Norway, National Veterinary Institute 5:1-12.
Matsuo, K. and I. Miyazono, 1993. The influence of long - term administration of peptidoglycan on disease resistance and growth of juvenile rainbow trout. Nippon Suisan Gakkaishi 59:1377-1379.
Matsushita, M., Endo, Y., Nonaka, M. and Fujita, T. 1998. Complement-related serine proteases in tunicates and vertebrates. Current Opin. Immunol. 10:29-35. Nakao, M. and T. Yano. 1998. Structural and functional identification of complement components of the bony fish, carp (Cyprinus carpio). Immunol. Rev. 166:27- 38.
Newman, K. 1994. Mannan oligosaccharides: natural polymers with significant impact on gastrointestinal microflora and the immune system. In: Biotechnology in Feed Industry, Proceedings of Alltech’s 10th Annual Symposium (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, UK, pp. 167-174.
Nonaka, M., N. Yamaguchi, S. Natsuume-Sakai and M. Takahashi. 1981a. The complement system of rainbow trout (Salmo gairdneri) I. Identification of the serum lytic system homologous to mammalian complement. J. Immunol. 126:1489-1494.
Paulsen S.M., R.E. Engstad and B. Robertsen. 2001. Enhanced lysozyme production in Atlantic salmon (Salmo salar L.) macrophages treated with yeast betaglucan and bacterial lipopolysaccharide. Fish & Shellfish Immunol. 11(1):23-37.
Raa, J. 1996. The use of immunostimulatory substances in fish and shellfish farming. Rev. Fisheries Sci. 4(3):229-288. CRC Press.
Raa, J. 2000. The use of immuno – stimulants in fish and shellfish feeds. In: Avances en Nutricion Acucola V. Memorias del V Simposium Int de Nutricion Acucola (L.E. Cruz-Suarez, D. Ricque-Marie, M.A. Olvera-Novoa and R. Civera-Cerecedo, eds). 19-22 Noviembre, 2000. Merida, Yucatan, Mexico.
Raa, J., G. Rorstad, R. Engstad and B. Robersten. 1992. The use of immunostimulants to increase resistance of aquatic organisms to microbial infections. In: Diseases in Asian Aquaculture (I.M. Shariff, R.P. Subasingle and J.R. Arthurs, eds). Fish Health Section, Asian Fisheries Society, Manila, Philippines, pp. 39- 50.
Ralph, G. and L. Phi-van. 1998. Posttranscriptional lipopolysaccharide regulation of the lysozyme gene at processing of the primary transcript in myelomonocytic HD11 cells. J. Immunol. 160:4970 4978.
Sakai, D.K. 1983. The activation of alternative pathway by pronase, LPS and Zymosan in the complement system of rainbow trout serum. Bulletin Japan Soc. Sci. Fisheries 49:347-351.
Sakai, D.K. 1984a. Opsonization by fish antibody and complement in the immune phagocytosis by peritoneal exudate cells isolated from salmonid fishes. J. Fish Dis. 7:29-38.
Sakai, D. K. 1984b. The non-specific activation of rainbow trout, Salmo gairdneri Richardson, complement by Aeromonas salmonicida extracellular products and the correlation of complement activity with the inactivation of lethal toxicity products. J. Fish Dis. 7:329-338.
Sakai, D.K. 1992. Repertoire of complement in immunological defense mechanisms of fish. Ann. Rev. Fish Dis. 223-247.
Sims, M. D., K. A. Dawson, K. E. Newman, P. Spring and D. M. Hooge. 2004. Effects of dietary mannan oligosaccharide, bacitracin methylene disalicylate, or both on the live performance and intestinal microbiology of turkeys. Poult. Sci. 83:1148-1154.
Siwicki A.K., M. Morand, P. Klein, M. Studnicka and E. Terech-Majewska. 1998. Modulation of nonspecific defence mechanisms and protection against diseases in fish. Acta Vet. Brno. 67:323-328.
Sotirov L.K. 1986. Method for determination of the alternative pathway of complement activation in some animals and man. In: Fourth Scientific Conference of Agriculture, Stara Zagora, pp. 1-10.
Spais, A.B., A.I. Giannenas, P. Florou-Paneri, E. Christaki and N.A. Botsoglou. 2003. Effect of the feed supplement Bio-Mos®, a mannan ologosaccharide, on the performance of broiler chickens. Hellenic Vet. Med. Soc. 54:111-118.
Spring, P. 1999. Mannanoligosaccharide as an alternative to antibiotic use in Europe. Zoot. Intern. 22:38-41.
Spring, P. 2002. Manipulating gut microflora through phosphorylated mannanoligosaccharides. Proceedings of Alltech’s 16th Annual European Lecture Tour, pp. 13-25.
Spring, P., C. Wenk, K.A. Dawson and K.E. Newman. 2000. Effect of mannan oligosaccharide on different cecal parameters and on cecal concentrations on enteric bacieria in challenged broiler chicks. Poult. Sci. 79:205-211.
Stelzner, A. and G. Stein. 1971. Moglichkikten zur hamolytischen Aktivitatsmessung von Gesamtkomplement. Theoretische Grundlangen und methodische Hinweise. Wissenschaftliche Zeitschrift der Friedrich Schiler Universitat Jena. Mathematisch Naturwissenschaftlische Reihe 20, Jg H6:933-941.
Sunyer, J.O., L. Tort and J.D. Lambris. 1997. Diversity of the third form of complement, C3, in fish: functional characterization of five forms of C3 in the diploid fish Sparus aurata. Biochem. J. 326:877-881.
Sunyer, J.O. and J.D. Lambris. 1998. Evolution and diversity of the complement system of poikilothermic vertebrates. Immunol. Rev. 166:39-57.
Waldroup, P.W., E.O. Oviedo-Rondon and C.A. Fritts. 2003. Comparison of Bio-Mos® and antibiotic feeding programs in broiler diets containing cooper sulfate. Int. J. Poult. Sci. 2(1):28-31.
Yoshida, T., R. Kruger and V. Inglis. 1995. Augmentation of non-specific protection of African catfish, Clarias gariepinus (Burch), by the long-term oral administartion of immunostimulants. J. Fish Dis. 18:195-198.
Zelikoff, J. T. 1998. Biomarkers of immunotoxicity in fish and other non-mammalian sentinel species: predictive value for mammals. Toxicology 129:63- 71.
Zelikof, J.T, A. Raymond, E. Carlson, Y. Li, J.R. Beaman and M. Anderson. 2000. Biomarkers of immunotoxicity in fish: from lab to ocean. Toxicol. Lett. 112-113:325-331.
Zhou, X., Y. Li, Y. Wu, D. Fagan and C. Mao. (in press). The effect of mannan oligosaccharide (Aqua- Mos) on performance, intestinal microflora and immune function of juvenile Jian carp (Cyprinus carpio Var. Jian).
Zhou, X-Q. and Y-L. Li. 2004. The effect of Bio-Mos® on intestinal microflora and immune function of juvenile Jian carp (Cyprinus carpio Var. Jian). In: Nutritional Biotechnology in the Feed and Food Industries, Proceedings of Alltech’s 20th Annual Symposium (Suppl. 1: Abstracts of posters presented). Lexington, Ky, May 24-26, p. 109.
Authors: YORDAN STAYKOV1, PETER SPRING2 and STEFAN DENEV1
1 Trakia University, Stara Zagora 6000, Bulgaria
2 Swiss College of Agriculture, Zollikofen, Switzerland


.jpg&w=3840&q=75)