Introduction
Influenza A virus (IAV) causes significant epidemics of respiratory disease in humans that result in human deaths and raise public health concerns that require a deeper understanding of IAV epidemiology and control. IAV is shared among animals and people and novel viruses capable of causing pandemics are the result of reassortant viruses from different species. Despite evidence that reassortment can happen in various species, swine is often labeled as the “mixing vessel” since swine have receptors capable to replicate influenza viruses of avian, human and swine origin. Because these viruses can infect humans, understanding transmission of swine-origin IAVs should be a priority.
In addition to IAV being a major pathogen for humans, IAV is also a serious problem in swine causing frequent outbreaks that involve both animal illness and zoonotic infections [1– 3]. In swine, IAV is distributed worldwide and is endemic in the US swine herd [2]. For almost a century, classical H1N1 viruses were the dominant IAVs until the appearance and subsequent circulation of double and triple reassortants since 1998 [4–6]. More recently, the 2009 pandemic virus [7], and the on-going influx of human-origin IAVs in swine [8, 9] has led to a more complex epidemiologic picture, making control of influenza in swine very difficult. The 2009 H1N1 pandemic, as well as outbreaks of variant H3N2 (H3N2v) influenza have demonstrated the potential for swine origin IAVs to cause significant morbidity and mortality globally, impacting the general public, swine workers and animal agriculture [10, 11]. Swine workers in particular, and their non-swine-exposed spouses, have been shown to be at a higher risk of swine-origin IAV infections than the general public [12], leading to calls for including such workers in pandemic preparedness and surveillance [13]. Since both direct and indirect contact exposures in commercial swine and agricultural fairs have been suspected in IAV zoonotic infections [10, 13], influenza prevention efforts involving swine production need to address multiple potential exposure routes.
While it is known that transmission of IAV occurs by direct contact, IAV can also be transmitted through indirect routes. Transmission of IAV via contaminated personnel and fomites has been documented in pigs [14] and aerosol transmission of IAV has been reported in various species [15–21]. In swine, IAV has been detected in aerosols from immune swine [22–24] and more recently IAV was isolated from air samples from inside and outside swine farms [23], and live animal markets in Minnesota [3].
Despite the growing evidence of indirect transmission of swine-origin IAV, there is limited information on the natural dynamics of IAV outbreaks in swine environments including production facilities. Information is lacking on levels of exposure encountered by both swine and people exposed to swine aerosols or contaminated surfaces in swine facilities during outbreaks of IAV. Therefore, our objective was to characterize viral load, viability and persistence of IAV in the air and on surfaces during periods of active IAV outbreaks in swine production facilities. This knowledge would further our understanding of the risk of IAV transmission between swine and people, and help inform prevention efforts.
Material and Methods
Procedures and protocols used in this study were approved by the University of Minnesota Institutional Animal Care and Use Committee protocol # 1207B17281 and the Institutional Biosafety Committee protocol # 1208H18341. Prior to the start of the study signed consent forms were obtained from the participating herds and forms were signed by herd owners or the production managers. No protected species were sampled.
Farm identification and selection
Eleven investigations of IAV outbreaks in six swine farms were conducted from October 2012 to May 2013. Farms with suspected outbreaks were identified by contacting veterinarians in Southern Minnesota and Northern Iowa. Veterinarians were asked to alert the investigators upon sudden onset of respiratory clinical signs suggestive of acute influenza in a swine herd (i.e. rapid onset of widespread dry hacking cough, sneezing, rhinorrhea, anorexia and lethargy). Each investigation consisted of visiting a candidate farm multiple times to assess herd health, collect samples, and gather additional information including temperature and relative humidity data. Farms were included in the study if the veterinarian made a presumptive diagnosis of IAV infection in the herd or was able to collect samples and confirm the diagnosis within 4 days from the onset of clinical signs, and was able to communicate with the investigators within 3 days from the onset of disease.
The investigators visited the farm within 1 to 3 days of being contacted and the clinical history of the outbreak was reviewed after interviews with farm personnel. During each visit air samples from inside and outside, pig oral fluids and surface samples were collected. The number of visits in each investigation varied based on diagnostic results on samples from the prior visit. If a herd tested negative in oral fluids, the investigation for that herd was concluded in terms of additional visits for most of the cases. The number of visits per farm ranged between 1 and 10 and the longest outbreak was 42 days. A summary of the farm characteristics is shown in Table 1.
Table 1. Summary of farm characteristics.
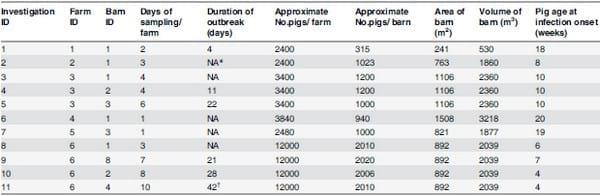
*NA: Non-applicable (investigation was IAV negative)
†: This investigation was considered two be two outbreaks
Sampling procedures and sampling scheme
Oral fluids. Oral fluids were collected to determine whether IAV was present in the swine at the time of sampling. Swine oral fluids were sampled using ropes as described previously [25, 26]. Briefly, two 3-strand twisted cotton ropes (WebRiggingSupply.com, Barrington, IL 60010, USA) were placed in 2–4 pens for 20 min for the swine to chew on the ropes. Each rope was estimated to sample approximately between 20 to 50 swine depending on pen size. Oral fluids were extracted from the rope immediately after collection by wringing the wet portion into a plastic bag and then the fluid was transferred into a 5 ml plastic sterile tube, and samples refrigerated at 4°C until processing. Oral fluid samples were processed within 24 hours of collection, centrifuged for 10 min at 5,000 RPM, and stored at -80°C until tested by RRT-PCR (real time reverse transcriptase polymerase chain reaction) and virus isolation.
Air sampling. Upon arrival at the farm, the first set of air samples was collected outside the barn approximately 25 m upwind (n = 2). After that, the second set was collected downwind (n = 2) from the facility at approximately the same distance, and lastly the final set was collected in the barn interior (n = 2). For the air interior samples, air collectors were placed within the barn at approximately 1/3 and 2/3 the length of the building and 1.5 m above the floor. Each set of samples was collected simultaneously as duplicates. Swine did not have direct contact with the air collectors.
Air samples were collected using a liquid cyclonic collector (Midwest Micro-Tek, Brookings, SD, USA) capable of processing 200 L / min of air [23, 27]. Briefly, 10 mL of minimum essential medium (MEM) solution supplemented with 2% bovine serum albumin (BSA) were added to the collection vessel, and samples collected for 30 minutes. About 4 mL of collection media were recovered for each sample, media were transferred into a plastic vial with a syringe and stored on ice until transport within 12 hours to the laboratory.
Surface sampling. Surface samples were collected from areas considered to have high contact by humans working in the barns including pen railings (n = 2) and door handles from doors leading into the swine barns (n = 1). Surface samples were collected using a 2”x2” sterile gauze dipped into sterile MEM supplemented with 2% BSA. Sections of 1 m of pen railing with approximately 0.08 m2 (800 cm2) of surface, were wiped for 30 seconds using sterile gloves. Door handles were wiped for 15 seconds and both the exterior and interior handles were sampled. Pigs did not have direct contact with the pen railing as only the top railing was sampled. Gauzes were placed into individual tubes and samples stored on ice for transport and processing within 24 hours.
Oral fluids and surface samples were collected simultaneously at the same time that the air interior was being sampled
Diagnostic procedures
Oral fluid samples were first screened at the University of Minnesota Veterinary Diagnostic Laboratory for influenza A RNA by a RRT-PCR targeting the matrix gene [28]. Samples with a cycle threshold (ct) value <35 were considered positive, 35–40 suspect, and >40 negative. Samples with ct < 40 were further tested using a quantitative RRT-PCR test as described previously [23]. RRT-PCR positive samples were cultured for virus isolation using Madin-Darby Canine Kidney (MDCK) cells [29, 30] and subtyped using the Path-ID Multiplex One-Step RRT-PCR kit (Applied Biosystems, Foster City, CA, USA) and custom subtyping assay primers and probes (Life Technologies) [31].
Swine clinical scores
Swine were visually inspected during each visit by a veterinarian member of the study team. Clinical scores consisted of coughing and sneezing and were measured following previously described procedures [32]. Briefly, the number of cough and sneeze episodes observed in 4 pens during 3 minutes were recorded. A cough or sneeze episode was defined as one or several coughs or sneezes in a sequence by an individual pig. The percentage of coughing or sneezing swine was calculated by dividing the number of swine observed coughing or sneezing by the total number of animals observed in the pens. The total number of swine evaluated in each visit ranged from 100 to 400 depending on pen and barn size.
Environmental conditions
Temperature and relative humidity inside and outside the barns were recorded at the time of collection using a weather meter (Kerstrel 3000, Nielsen-Kellerman, PA, USA).
Statistical and influenza modeling in indoor air and statistical analysis
Statistical analyses were conducted using R programming language [33].
To look for associations between the count of positive samples of each type compared with each other type, we performed pair-wise Kendall’s rank correlation tests, corrected for multiple comparisons with the Bonferroni-Holm adjustment. Correlations between quantity of IAV RNA copies between samples of oral fluids, surface and air inside the barns were also computed using Kendall’s correlation. Correlations of these samples with clinical scores were computed as well. Correlations between quantity of IAV in indoor air with recorded measurements of relative humidity and temperature were also determined.
To compute correlations and modeling of IAV in indoor air, data from four investigations with at least 5 days of samples were used (investigations 5, 9, 10 and 11). Data were limited to the first 21 days after the reported onset given that most of the farms tested negative after that, and the mean indoor air IAV quantity was calculated for each visit. The concentration of IAV in the air inside the barn as a function of time over the outbreak was modeled using a quasipoisson model with log link to appropriately handle both the days at which zeros were recorded and the fact that the variance increased with the quantity of virus detected. Additionally, as the reported day of onset may have been early or late relative to the true progress of the infection, the reported day of onset was allowed to shift relative to the estimated maximum for each investigation to minimize the deviance of the fit.
A quadratic effect was used for day and an additive blocking effect was used to allow each investigation to have a different maximum value; the fitted equation was:
Indoor air quantity = M_exp(-0.035_day—0.082_day^2), where Mis the maximum value for that investigation, and day is relative to the day of the maximum.
Results
Clinical signs
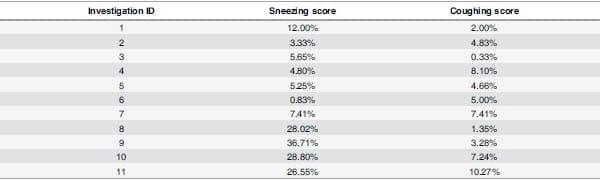
Clinical scores of coughing and sneezing were recorded in both IAV positive and negative investigations. Mean scores ranged from 0.83% to 36.71% and 0.33% to 10.27% for sneezing and coughing, respectively (Table 2) and there was variation in the scores throughout the course of the investigations (results not shown).
Table 2. Average clinical scores of sneezing and coughing for each investigation.
Influenza infections
Eleven suspected IAV outbreak investigations in barns corresponding to 6 farms were identified during the study. There were a total of 49 farm visits, which took place between 2 to 8 days apart for 4 to 42 days after the initial visit. Six of the 11 barn investigations, corresponding to three different farms, were confirmed positive for IAV by RRT-PCR testing in aerosols, surfaces, and/or swine oral fluid samples.
Forty-seven out of 98 (48%) oral fluid samples tested were RRT-PCR positive for IAV while 32 of 84 (38%) pen railing samples, and 35 of 82 (43%) indoor air samples tested positive for IAV (Table 3). There were two door handle samples that tested positive at low levels. All air samples collected outdoors tested negative. There was a significant positive correlation of 0.69 between the count of oral fluid positive samples and air (p = 0.0001), of 0.47 between oral fluids and pen railing (p = 0.009) and 0.42 between indoor air and pen railing (p = 0.01).
Table 3. Total number of samples positive for influenza A virus real time reverse transcriptase polymerase chain reaction (RRT-PCR), per sample type and investigation.
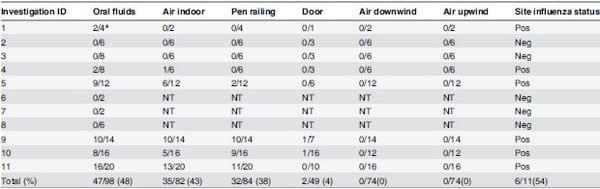
* Number of positive RRT-PCR/total number of samples; NT: not tested
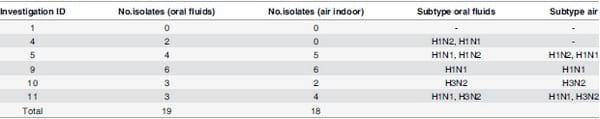
IAV was isolated by culture from 19 oral fluid and 18 indoor air samples (Table 4) representing five and four investigations, respectively. H1N1, H1N2 and H3N2 subtypes, and mixtures of these, were identified in both, oral fluid and indoor air samples. Virus isolation of surface samples did not yield positive results.
Table 4. Influenza A virus subtype and number of isolates cultured from oral fluids and indoor air samples in each confirmed positive investigation.
Viral quantification
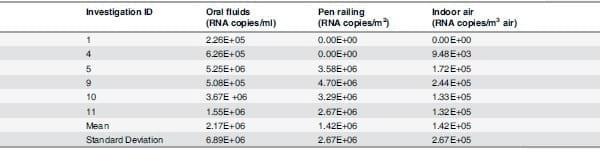
Influenza RNA levels in oral fluids, indoor air and pen railing varied between farms and throughout the course of the clinical outbreaks (Table 5). Individual sample viral levels ranged from0 to 4.03x107 RNA copies/ml in oral fluids, 0 to 4.16x107 RNA copies/m2) in pen railing surface and 0 to 1.25x106 RNA copies/m3 of air in indoor air samples. There was a significant positive correlation of 0.4 between quantity of IAV in the air and oral fluids (p = 0.015) and of 0.372 between quantity of IAV in oral fluids and coughing (p = 0.023) (Table 6). Correlations between quantity of IAV on the pen railing with both the air and oral fluids were not significant (p>0.05).
Table 5. Mean influenza A virus (IAV) quantitative real time reverse transcriptase polymerase chain reaction (RRT-PCR) results for oral fluids, pen railing and indoor air samples from IAV positive farms.
Table 6. Correlations of quantity of influenza A virus in oral fluids of groups of pigs and environmentand with percentage of pigs exhibiting coughing, sneezing or signs of illness.
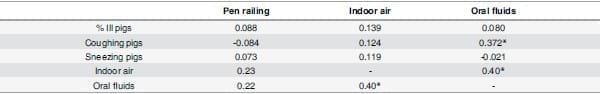
* Differences p<0.05
Environmental conditions
Measured mean indoor temperatures ranged between 19°C and 25°C while relative humidity ranged between 19°C and 25°C. Both of these ranges were within the expected ranges for swine commercial facilities. Correlations between quantity of IAV in indoor air samples with relative humidity and temperature were 0.26 (p = 0.12) and 0.01 (p = 0.93) respectively.
Influenza level modeling
Results ofmodeling of indoor air levels of IAV are shown in Fig 1 based on results from 4 investigations. The best fit for the reported day of onset relative to the estimated maximumwas -11, -11, -7 and -9 days for investigations 10, 11, 5 and 9, respectively. These results indicated a short spread between farms in the duration of IAV detection in the air. The model also showed the best fit for the estimated mean maximum RNA copy viral load (x10^4 RNA copies/m3) with 95% confidence interval for each investigation at 12 (4.5, 31), 38 (21, 68), 60 (38, 97), and 71 (49, 103) for investigations 11, 5, 10, and 9 respectively, indicating differences in the modeled levels of airborne IAV between farms and throughout the duration of an outbreak. Data from oral fluids and surfaces could not be modeled because of a lack of a common pattern in the data obtained.
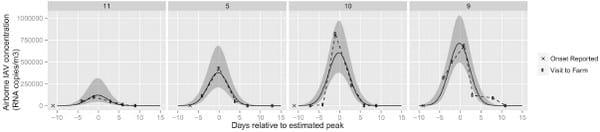
Fig 1. Modeled influenza A virus quantity (RNA copies/m3) in indoor air. Fig shows the data, connected by dashed lines, and the fitted values in a solid line, as well as 95% confidence interval for the fitted values. Axis x represents the days relative to the estimated peak of the outbreak. The date when the onset was reported is also shown.
Discussion
Despite the common occurrence of IAV infections in swine, there is limited information on the levels and persistence of IAV in the air and environment of swine production facilities. This is the first study, to our knowledge, that has quantified and characterized the level of IAV in samples of aerosols and surfaces of swine environments during acute outbreaks of influenza infections in swine. We found that IAV could be isolated from indoor air of commercial swine production facilities, that airborne IAV levels were sustained for periods of 20 days and that there was a correlation between the number of positive samples of each type and the quantity of virus in the swine oral fluids and in the air. Our results provide a first estimation on levels of environmental IAV in swine commercial production facilities, and thus an assessment of potential sources of IAV exposure to swine workers or other pigs.
Detection of IAV in air was sustained throughout the course of the acute outbreaks and lasted approximately 20 days across the studied barns. The peak of detection of virus in air samples occurred between 7 and 11 days into the outbreaks. Maximum airborne levels varied between affected facilities and were in the order magnitude of 104 to 107 RNA copies/m3 of air.
We allowed our model to shift relative to the estimated maximum for each investigation given that the reported day may not have been when the infection truly started. Interestingly the IAV detection curves were similar across farms with limited variations in duration which suggests a similar course of disease between farms and, that similar measures could be implemented to minimize risk of IAV infections. There were differences in the levels of IAV found in the air between farms and we speculate that these differences could be due to varying levels of infection and immunity in the swine, type of IAV, number of pigs in the barn, barn volume, ventilation rates and farm and management characteristics. Thus, information from our model can be used to estimate risk of exposure to swine workers, other pigs and help target intervention strategies to mitigate the risk of IAV transmission between pigs and from pigs to people. A key aspect of determining the risk of IAV transmission is the relationship between number of RNA copies and infectivity. We estimated the ratio of viral particles to TCID50 (tissue culture infectious dose) at 3,000 RNA copies/TCID (results not shown) and based on the mean airborne IAV concentration, our results corresponded to 47 TCID50/m3. Similar TCID50 estimates were obtained in a health center [34] and although it is unclear how our results relate to transmission to swine or people, we speculate that they represent a significant risk to both people and swine, since IAV was readily isolated from the air multiple times throughout the duration of the outbreaks. Overall, our results provide evidence that air can be an important route of IAV transmission in swine production facilities. Furthermore, there was an association between the levels of IAV in oral fluids and the air indicating a direct relationship between level of virus in the swine and potential exposure through aerosols. However, further studies are needed to fully understand the relationship between airborne IAV levels and transmission. We isolated a mixture of genetically diverse IAV from swine and air samples representing the three most common IAV subtypes in swine, H1N1, H1N2 and H3N2. However, in contrast to a prior study [23], we did not detect IAV in air samples collected outside swine facilities. This difference is probably a result of sampling frequency and our investigations being carried out under colder environmental conditions and larger distances from the air exhaust site that likely negatively impacted both distribution of the virus and viral survivability. Therefore more research is needed to fully characterize the risk of IAV transmission outside swine production facilities.
IAV genetic material was also detected in surfaces, in particular on pen railings although we could not isolate IAV from surfaces. Source of IAV genetic material in the surfaces may be the result of deposition of airborne IAV particles. Whether inability to culture IAV from surfaces was due to lack of viable IAV in surfaces or conditions of sampling or limited sensitivity of the culture technique could not be assessed, but we speculate that viable IAV can still be present on surfaces from swine barns although with less quantity than in the air due to environmental conditions such as desiccation or preservation in dust. In deed surface contamination with viable IAV was shown in a live animal market housing swine [3]. Therefore, precautions to prevent exposure to contaminated surfaces should still be followed.
Clinical signs of coughing and sneezing can be an indicator of IAV infections and IAV can be found in both clinically and subclinically infected swine [2]. We found an association between coughing and levels of IAV in the swine but not in air or on surfaces. We did not have a common pattern on the presentation and evolution of clinical signs across farms. This could be due to presence of concomitant infections or farm factors not measured in this study. Overall our results indicate that clinical signs in swine cannot be used as a reliable indicator of the levels of IAV present in the environment and thus they should not be used to predict risk of exposure to people.
The farm investigations in this study were selected by convenience based on recognized acute clinical signs in the swine herds, thus results from this study should be interpreted carefully when extrapolating them to endemically infected farms. Furthermore data in this study was obtained from a limited number of farms which do not represent the full spectrum of types of production facilities and management conditions encountered in the swine industry. Thus further research is needed to characterize the levels and risk of IAV environmental exposure in non-outbreak situations and production facilities representative of different management and environmental conditions.
Lastly, although environmental conditions of relative humidity and temperature have been associated with IAV viability [35], in this study we did not see an association between quantity of IAV in air or on surfaces and relative humidity or temperature. This lack of association could be the result of the relatively stable indoor conditions throughout our study. Producers put a great deal of effort into maintaining a relatively stable temperature and relative humidity within these facilities to maintain the health and safety of the animals.
In summary, our results indicate that during outbreaks of IAV in swine, the air and surfaces in barns contain significant levels of IAV potentially representing an exposure hazard to both swine and people. Further studies are needed to evaluate the viability of IAV in the environment, evaluate strategies to mitigate the risk of indirect transmission of IAV, confirm the impact of personal protective equipment on exposure risk to people and explore strategies to prevent bidirectional transmission of IAV between humans and swine. Information from this study should help to develop evidence-based guidelines to minimize the impact of IAV infections on swine production.
Acknowledgments
Funding was provided by the National Pork Board. The authors would like to acknowledge the contributions of My Yang, Andres Diaz and Macarena Cortez for technical assistance.
Author Contributions
Conceived and designed the experiments: MT SGG PR. Performed the experiments: VN MT. Analyzed the data: AR VN MT. Contributed reagents/materials/analysis tools: SGG PR BP AR. Wrote the paper: MT VN. Reviewed critically the manuscript: SGG PR BP AR.
This article was originally published in PLoS ONE 11(1): e0146616. doi:10.1371/ journal.pone.0146616. This is an Open Access article distributed under the terms of the Creative Commons Attribution License.
References
1. Myers KP, Olsen CW, Gray GC. Cases of swine influenza in humans: A review of the literature. Clinical Infectious Diseases. 2007; 44(8):1084–8. doi: 10.1086/512813 PMID: WOS:000244928200012.
2. Corzo CA, Culhane M, Juleen K, Stigger-Rosser E, Ducatez MF, Webby RJ, et al. Active Surveillance for Influenza A Virus among Swine, Midwestern United States, 2009–2011. Emerging Infectious Diseases. 2013; 19(6):954–60. doi: 10.3201/eid1906.121637 PMID: WOS:000328173500015.
3. Choi MJ, Torremorell M, Bender JB, Smith K, Boxrud D, Ertl JR, et al. Live animal markets in Minnesota: a potential source for emergence of novel influenza A viruses and interspecies transmission. Clin Infect Dis. 2015. Epub 2015/08/01. doi: 10.1093/cid/civ618 PMID: 26223994.
4. Zhou NN, Senne DA, Landgraf JS, Swenson SL, Erickson G, Rossow K, et al. Genetic reassortment of avian, swine, and human influenza A viruses in American pigs. Journal of Virology. 1999; 73(10):8851– 6. PMID: WOS:000082554300105.
5. Webby RJ, Rossow K, Erickson G, Sims Y, Webster R. Multiple lineages of antigenically and genetically diverse influenza A virus co-circulate in the United States swine population. Virus Research. 2004; 103(1–2):67–73. doi: 10.1016/j.virusres.2004.02.015 PMID: WOS:000222065200012.
6. Karasin AI, Schutten MM, Cooper LA, Smith CB, Subbarao K, Anderson GA, et al. Genetic characterization of H3N2 influenza viruses isolated from pigs in North America, 1977–1999: evidence for wholly human and reassortant virus genotypes. Virus Research. 2000; 68(1):71–85. doi: 10.1016/s0168-1702 (00)00154-4 PMID: WOS:000088282200008.
7. Forgie SE, Keenliside J, Wilkinson C, Webby R, Lu P, Sorensen O, et al. Swine Outbreak of Pandemic Influenza A Virus on a Canadian Research Farm Supports Human-to-Swine Transmission. Clinical Infectious Diseases. 2011; 52(1):10–8. doi: 10.1093/cid/ciq030 PMID: WOS:000286214200003.
8. Nelson MI, Gramer MR, Vincent AL, Holmes EC. Global transmission of influenza viruses from humans to swine. Journal of General Virology. 2012; 93:2195–203. doi: 10.1099/vir.0.044974–0 PMID: WOS:000310043400012.
9. Nelson MI, Wentworth DE, Culhane MR, Vincent AL, Viboud C, LaPointe MP, et al. Introductions and Evolution of Human-Origin Seasonal Influenza A Viruses in Multinational Swine Populations. Journal of Virology. 2014; 88(17):10110–9. doi: 10.1128/jvi.01080-14 PMID: WOS:000341232300051.
10. Jhung MA, Epperson S, Biggerstaff M, Allen D, Balish A, Barnes N, et al. Outbreak of Variant Influenza A(H3N2) Virus in the United States. Clinical Infectious Diseases. 2013; 57(12):1703–12. doi: 10.1093/ cid/cit649 PMID: WOS:000327720400008.
11. Smith GJD, Vijaykrishna D, Bahl J, Lycett SJ, Worobey M, Pybus OG, et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature. 2009; 459(7250):1122–U107. doi: 10.1038/nature08182 PMID: WOS:000267636700042.
12. Gray GC, McCarthy T, Capuano AW, Setterquist SF, Olsen CW, Alavanja MC. Swine workers and swine influenza virus infections. Emerg Infect Dis. 2007; 13(12):1871–8. Epub 2008/02/09. doi: 10. 3201/eid1312.061323 PMID: 18258038; PubMed Central PMCID: PMCPMC2876739.
13. Myers KP, Olsen CW, Setterquist SF, Capuano AW, Donham KJ, Thacker EL, et al. Are swine workers in the United States at increased risk of infection with zoonotic influenza virus? Clinical Infectious Diseases. 2006; 42(1):14–20. doi: 10.1086/498977 PMID: WOS:000233698900009.
14. Allerson MW, Cardona CJ, Torremorell M. Indirect Transmission of Influenza A Virus between Pig Populations under Two Different Biosecurity Settings. Plos One. 2013; 8(6):9. doi: 10.1371/journal.pone. 0067293 PMID: WOS:000320846500134.
15. Brankston G, Gitterman L, Hirji Z, Lemieux C, Gardam M. Transmission of influenza A in human beings. Lancet Infectious Diseases. 2007; 7(4):257–65. doi: 10.1016/s1473-3099(07)70029-4 PMID: WOS:000245120700027.
16. Blachere FM, Lindsley WG, Pearce TA, Anderson SE, Fisher M, Khakoo R, et al. Measurement of Airborne Influenza Virus in a Hospital Emergency Department. Clinical Infectious Diseases. 2009; 48 (4):438–40. doi: 10.1086/596478 PMID: WOS:000262585700011.
17. Tellier R. Aerosol transmission of influenza A virus: a review of new studies. Journal of the Royal Society Interface. 2009; 6:S783–S90. doi: 10.1098/rsif.2009.0302.focus PMID: WOS:000271957900009.
18. Mubareka S, Lowen AC, Steel J, Coates AL, Garcia-Sastre A, Palese P. Transmission of Influenza Virus via Aerosols and Fomites in the Guinea Pig Model. Journal of Infectious Diseases. 2009; 199 (6):858–65. doi: 10.1086/597073 PMID: WOS:000263677900013.
19. Munster VJ, de Wit E, van den Brand JMA, Herfst S, Schrauwen EJA, Bestebroer TM, et al. Pathogenesis and Transmission of Swine-Origin 2009 A(H1N1) Influenza Virus in Ferrets. Science. 2009; 325 (5939):481–3. doi: 10.1126/science.1177127 PMID: WOS:000268255100058.
20. Yee KS, Cardona CJ, Carpenter TE. Transmission of low-pathogenicity avian influenza virus of subtype H6N2 from chickens to Pekin ducks and Japanese quail (Coturnix coturnix japonica). Avian Pathology. 2009; 38(1):59–64. doi: 10.1080/03079450802632023 PMID: WOS:000262514600010.
21. Schulman JL. Experimental transmission of influenza virus infection in mice. IV. Relationship of transmissibility of different strains of virus and recovery of airborne virus in the environment of infector mice. J Exp Med. 1967; 125(3):479–88. Epub 1967/03/01. PMID: 6016901; PubMed Central PMCID: PMCPMC2138293.
22. Loeffen WLA, Stockhofe N, Weesendorp E, van Zoelen-Bos D, Heutink R, Quak S, et al. Efficacy of a pandemic (H1N1) 2009 virus vaccine in pigs against the pandemic influenza virus is superior to commercially available swine influenza vaccines. Veterinary Microbiology. 2011; 152(3–4):304–14. doi: 10. 1016/j.vetmic.2011.05.027 PMID: WOS:000294937500012.
23. Corzo CA, Culhane M, Dee S, Morrison RB, Torremorell M. Airborne Detection and Quantification of Swine Influenza A Virus in Air Samples Collected Inside, Outside and Downwind from Swine Barns. Plos One. 2013; 8(8). doi: 10.1371/journal.pone.0071444 PMID: WOS:000323124000058.
24. Corzo CA, Allerson M, Gramer M, Morrison RB, Torremorell M. Detection of Airborne Influenza A Virus in Experimentally Infected Pigs With Maternally Derived Antibodies. Transboundary and Emerging Diseases. 2014; 61(1):28–36. doi: 10.1111/j.1865-1682.2012.01367.x PMID: WOS:000329310800005.
25. Prickett J, Simer R, Christopher-Hennings J, Yoon KJ, Evans RB, Zimmerman JJ. Detection of Porcine reproductive and respiratory syndrome virus infection in porcine oral fluid samples: a longitudinal study under experimental conditions. Journal of Veterinary Diagnostic Investigation. 2008; 20(2):156–63. PMID: WOS:000262533700003.
26. Romagosa A, Gramer M, Joo HS, Torremorell M. Sensitivity of oral fluids for detecting influenza A virus in populations of vaccinated and non-vaccinated pigs. Influenza and Other Respiratory Viruses. 2012; 6(2):110–8. doi: 10.1111/j.1750-2659.2011.00276.x PMID: WOS:000300689300006.
27. Dee S, Otake S, Oliveira S, Deen J. Evidence of long distance airborne transport of porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae. Veterinary Research. 2009; 40 (4):13. doi: 10.1051/vetres/2009022 PMID: WOS:000267963400009.
28. Slomka MJ, Densham ALE, Coward VJ, Essen S, Brookes SM, Irvine RM, et al. Real time reverse transcription (RRT)-polymerase chain reaction (PCR) methods for detection of pandemic (H1N1) 2009 influenza virus and European swine influenza A virus infections in pigs. Influenza and Other Respiratory Viruses. 2010; 4(5):277–93. doi: 10.1111/j.1750-2659.2010.00149.x PMID: WOS:000280976100005.
29. Detmer SE, Patnayak DP, Jiang Y, Gramer MR, Goyal SM. Detection of Influenza A virus in porcine oral fluid samples. Journal of Veterinary Diagnostic Investigation. 2011; 23(2):241–7. PMID: WOS:000288838300007.
30. Detmer S, Gramer M, Goyal S, Torremorell M, Torrison J. Diagnostics and Surveillance for Swine Influenza. In: Richt JA, Webby RJ, editors. Swine Influenza. Current Topics in Microbiology and Immunology. 370. Berlin: Springer-Verlag Berlin; 2013. p. 85–112.
31. Zhang JQ, Harmon KM. RNA Extraction from Swine Samples and Detection of Influenza A Virus in Swine by Real-Time RT-PCR. In: Spackman E, editor. Animal Influenza Virus, Second Edition. Methods in Molecular Biology. 1161. Totowa: Humana Press Inc; 2014. p. 277–93.
32. J S, A R, R R. Monitoramentos sanitarios. Goiania: Canone Editorial; 2007.
33. The R Project for Statistical Computing. Vienna, Austria: Foundation for Statistical Computing; 2008.
34. Yang W, Elankumaran S, Marr LC. Concentrations and size distributions of airborne influenza A viruses measured indoors at a health centre, a day-care centre and on aeroplanes. Journal of the Royal Society Interface. 2011; 8(61):1176–84. doi: 10.1098/rsif.2010.0686 PMID: WOS:000292083300010.
35. Irwin CK, Yoon KJ, Wang C, Hoff SJ, Zimmerman JJ, Denagamage T, et al. Using the Systematic Review Methodology To Evaluate Factors That Influence the Persistence of Influenza Virus in Environmental Matrices. Applied and Environmental Microbiology. 2011; 77(3):1049–60. doi: 10.1128/aem. 02733-09 PMID: WOS:000286597100041.