1. Introduction
Aflatoxins are naturally occurring mycotoxins that are produced by some strains of Aspergillus species which are commonly found in cereals worldwide and bring significant threats to the food industry and animal production [1]. At least 14 different types of aflatoxins are produced in nature [2,3]. Aflatoxin B1 (AFB1) is considered the most toxic and is produced by both Aspergillus flavus and Aspergillus parasiticus [4]. Several physical and chemical methods have been developed to reduce aflatoxins [2,3]. Unfortunately, these methods have restrictions in terms of product nutrition, organoleptic qualities, and adverse health effects, which motivate emphasis on biological methods of degradation of aflatoxins [5–7]. Bacillus spp. are probiotics accepted by human or animals as direct fed microbials (DFM). Our laboratory has showed the safety and efficacy of individual monocultures for prophylactic and/or therapeutic efficacy against Salmonella infections under both laboratory and field conditions as well as the development of a novel, cost-effective DFM with potential for widespread utilization and improved production, delivery and clinical efficacy for poultry [8–14].
The aim of this study was to screen Bacillus candidates capable of biotransforming AFB1. Hence, the DFM candidates could not only be used as probiotics but also as an antidote for aflatoxins.
2. Materials and methods
2.1. Isolation and characterization of Bacillus spp.
Previous research conducted in our laboratory focused on isolation of several Bacillus spp. from environmental and poultry sources [8–10,12]. Identification was carried out by using a bioMerieux API 50 CHB (catalog No. 50 430, Biomerieux, Durham, NC) test kit. Isolates generally recognized as safe (GRAS) was affirmed as described by Wolfenden et al. [8]. For our preliminary experiment, sixty-nine isolates were chosen based on consistent in vitro anti- Salmonella spp., Clostridium spp., and Campylobacter spp. activity (Data not shown).
2.2. In vitro evaluation of biodegradation of AFB1
A modified Czapek-Dox medium with the following composition per liter proved satisfactory: sucrose, 3.000%; NaNO3, 0.300%; K2HPO4, 0.100%; MgSO4, 0.050%; KCl, 0.050%; FeSO4, 0.001%; yeast extract (Difco, BD, Becton, Dickinson and Company; Sparks, MD 21152, USA; 38800 Le Pont de Clair, France), 0.005%; agar, 2.000% [5]. To evaluate AFB1 (Sigma–Aldrich, Oakville, ON) inhibition, standard solutions were diluted in chloroform and added to the medium to reach a final concentration of 1 µg/mL of medium, and while it was still hot, the chloroform was driven off. About 30 mL of the medium was added to each Petri dish and allowed to solidify. Sixty nine GRAS isolates were grown in tryptic soy broth (TSB) (catalog No. 211 822, Becton Dickinson, Sparks, MD) for 24 h at 37 °C and then washed 3 times in 0.9% sterile saline by centrifugation (3 900 r/min, 4 °C, 15 min). About 10 µL of each isolate was placed on the center of the Petri dish plate with modified Czapek-Dox medium. After point inoculation, the plates were incubated at 37 °C and examined at intervals of 1–2 days for up to 2 weeks under ultraviolet light (UV) for AFB1 utilization. On initial examination, plates had to be exposed to UV for about 15 min to develop fluorescence. Utilization of toxin was indicated by a zone of non-fluorescence in the colony.
2.3. Identification of candidate isolates
Out of the 69 GRAS isolates, three showed capacity to biodegrade AFB1 (data not shown). Those isolates were further identified by 16S rRNA sequence analysis (Microbial ID Inc., Newark, DE 19713, USA). Then, the candidate Bacillus strains were chosen for physiological tests as described by Menconi et al. [12], and sporulated. The biological detoxification of AFB1 was determined in an in vitro digestion model as described below.
2.4. Bile salt tolerance
The method of Gilliland et al. (1984) [15], with some modifications, was used to determine bile salt tolerance. TSB containing 0.000%, 0.037%, 0.075%, 0.150%, and 0.300% of bile salts No. 3 (Catalog No. 213 010, Becton Dickinson and Co., Sparks, MD 21152, USA) was inoculated with 107 CFU/ mL of each potential probiotic strain, after being centrifuged at 5 000 r/min for 15 min and washed 3 times from their 24 h growth cultures. Samples were incubated for 24 h at 37 °C with shaking at 100 r/min. Growth in control (no bile salts) and test cultures was evaluated at 2, 4, and 24 h by streaking samples on trypticase soy agar (TSA) (catalog No. 211 822, Becton Dickinson, Sparks, MD) for presence or absence of growth.
2.5. Resistance in conditions of the intestinal tract evaluation: pH, temperature, and NaCl
A basal TSB medium was used in these series of in vitro studies. A 24 h culture of each isolate was used as the inoculum whereby the cells were spun down and re-suspended in 0.9% sterile saline. Then, 100 µL of the suspension was inoculated into 10 mL of TSB of each test tube. Two incubation time points, i.e. 2 h and 4 h, were evaluated for each of the variables (pH, temperature, and NaCl). The rationale for these two points was mainly based on the transit time of food matter in the gastrointestinal tract of poultry. The temperatures tested were 15 and 45 °C. The concentrations of NaCl tested were 3.5% and 6.5% (w/v). The isolates were tested for growth at pH of 2 and 3. The tubes were incubated with reciprocal shaking, at the specific test temperatures or 37 °C for the tests on pH and concentrations of NaCl. At the time points evaluated, each sample was streaked on TSA for presence or absence of growth, to confirm livability of the strains. The turbidity of each tube was also noted as an indication of growth or no-growth. Each treatment was tested with triplicate tubes.
2.6. Antibiotic resistance
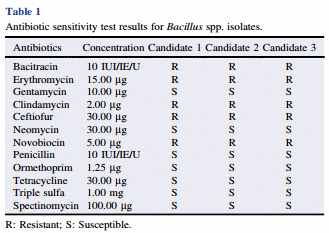
Selected colonies on TSA plates were inoculated and cultured for 24 h in TSB at 37 °C. Strains were then sent to a Veterinary Diagnostic Laboratory (University of Arkansas, Division of Agriculture, Fayetteville, AR, 72703, USA) for antibiotic sensitivity analysis by using Kirby–Bauer methodology. The diameter of the inhibition zones and the interpretative zone sizes were reported. Twelve antibiotics were tested, and their concentrations were reported as shown in Table 1. The results were expressed in terms of resistant, and susceptible.
2.7. Sporulation procedure
In an effort to grow high numbers of viable spores, a solid state fermentation (SSF) media developed by Zhao et al. was selected and modified for use in these experiments [16]. Briefly, a Rosario Galarza-Seeber et al./Asian Pac J Trop Biomed 2015; 5(9): 702–706 703 liquid media component was added to a mixture of 70% rice straw and 30% wheat bran at a rate of 40% by weight. The SSF media was added to a 250 mL Erlenmeyer flask and sterilized by autoclaving for 30 min at 121 °C. Candidate isolates were grown individually overnight at 37 °C in TSB, then 2 mL of a candidate culture was added to the prepared SSF media. The inoculated flasks were incubated for 24 h at 37 °C, and then incubated for another 72 h at 30 °C. The cultures were removed from their flasks, placed onto Petri dishes, and then dried at 60 °C. Following this, the cultures were aseptically ground into a fine powder to generate stable spores (~1011 spores/g). Spores were mixed into the feed with a rotary mixer for 15 min. Samples of feed containing the DFM candidate culture were taken and a 1:10 dilution was made with saline. All samples were subject to 100 °C for 10 min. These samples of ten-fold dilutions were placed on TSA, and incubated at 37 °C for 24 h to count the number of spores per gram of feed.
2.8. Determination of biological detoxification of AFB1 in an in vitro digestion model
Freshly prepared, unmedicated corn-soy based starter feed was used for all in vitro trials. DFM candidates were incorporated into 1 of 3 experimental feed groups: 1) Negative control group, with unmedicated starter broiler feed without AFB1; 2) Positive control group, with negative control feed contaminated with 0.01% AFB1; 3) DFM treated group, with positive control feed supplemented with 109 spores/g. In vitro digestion of the three diets with or without DFM supplementation was performed by triplicate according to previously published methods, with minor modifications [17]. All in vitro digestion steps were carried out at 40 °C to simulate avian body temperature, by using a water-jacketed incubator (Forma Scientific Inc., Marietta, OH, USA) customized with bars that rotated the tubes horizontally at 19 r/min. To mimic crop digestion, 50 g of each diet and 100 mL of 0.03 mol/L HCl were placed in 50 mL polypropylene centrifuge tubes and mixed vigorously. The pH was measured (ranged from 5.19 to 5.22) and the tubes were incubated for 30 min. Next, to mimic proventricular digestion, 150 000 IU pepsin (Sigma–Aldrich Canada Ltd Oakville, Ont., Canada) and 25 mL of 1.5 mol/L HCl were added to each tube. Values of pH were measured (ranged from 1.37 to 1.96) and the mixtures were then incubated for a further 45 min. Following this, 341.5 mg of 8'pancreatin (Sigma–Aldrich Canada Ltd.) was added in 32.5 mL of 1.0 mol/L NaHCO3, and the pH was adjusted to between 6.3 and 6.7 with 1.0 mol/L NaHCO3. Volumes were equalized in the tubes by adding distilled water, and the samples were incubated for a further 2 h. After removal of solids and awns, the samples were first centrifuged at 4 100 r/min for 5 min. After digestion time (3:15 h), supernatants and digesta (by triplicate) were collected for AFB1 analysis by highperformance liquid chromatography with fluorescence detection (HPLC-FLD) method by using a Romer Derivatization Unit (Romer Labs, Inc., MO 63084-1156, USA).
2.9. Statistical analysis
Data of the determination of biological detoxification of AFB1 by HPLC-FLD of the DFM candidates in an in vitro digestion model were subjected to ANOVA as a completely randomized design by using the GLM procedure of SAS/STAT® 9.2. Data were expressed as mean ± SE. Significant differences among the means were determined by using Duncan's multiplerange test at P < 0.05.
3. Results
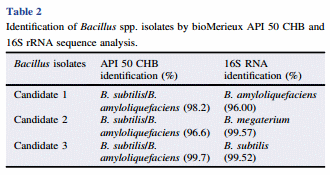
Table 2 shows the identification of Bacillus spp. isolates by bioMerieux API 50 CHB and 16S rRNA sequence analyzes. The three isolates were characterized as Bacillus subtilis (B. subtilis)/ Bacillus amyloliquefaciens (B. amyloliquefaciens) by the bioMerieux API identification kit. However, further sequence analysis of 16S rRNA, which is the predominant molecular technology currently available for microbial identification revealed that B. amyloliquefaciens was for candidate 1, Bacillus megaterium (B. megaterium) for candidate 2 and B. subtilis for candidate 3 (Table 2).
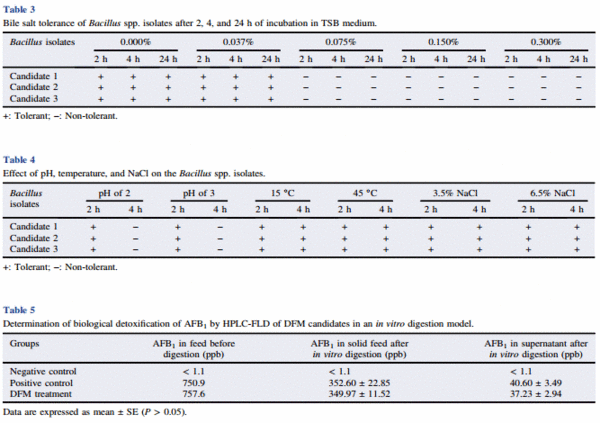
The results of the bile salt tolerance of the Bacillus spp. isolates after 2, 4, and 24 h of incubation are summarized in Table 3. All the three DFM candidates were able to grow when cultured at 0.037% bile salt concentration for 2 h, 4 h, and 24 h of incubation. The results of the effect of pH, temperature, and NaCl on the three DFM candidates are summarized in Table 4. Vegetative cells were evaluated for conditions similar to those found in the stomach. All three candidates were able to survive at pH 2 and pH 3 for 2 h. Furthermore, vegetative cells grew at 15 °C and 45 °C at both times of incubation of 2 h and 4 h and were also able to tolerate up to 6.5% of NaCl (Table 4).
The antibiotic resistance and susceptibility of the DFM candidates to twelve antibiotics are summarized in Table 1. All three DFM candidates were sensitive to gentamycin, neomycin, penicillin, ormethoprim, tetracycline, triple sulfa, and spectinomycin, and resistant to bacitracin, erythromycin, clindamycin, ceftiofur, and novobiocin (Table 1).
Table 5 summarizes the determination of biological detoxification of AFB1 by HPLC-FLD of the DFM candidates in an in vitro digestion model. In the present study, no significant differences (P > 0.05) were observed in the concentrations of AFB1 in neither the supernatants nor digesta samples evaluated by HPLC-FLD between positive control or DFM treated groups.
4. Discussion
Antibiotics as growth promoters in livestock have been in practice for over five decades. However, rising socio-political concerns with their use has prompted a quest for alternative methods of disease intervention and optimization of growth promotion in commercial poultry farming. The use of DFM as an alternative approach has gained momentum in recent years [8–14]. The advantages of application, pathogen reduction, immunomodulation, performance enhancement and synthesis of antimicrobials and enzymes have given probiotics and DFM a clear edge over antibiotics making their use highly sustainable [8–14]. Conversely, biological degradation of aflatoxins occurs in nature since aflatoxins are chemically stable but not appear to accumulate in natural environments [5]. Several investigators have demonstrated that microorganisms in the environment can be chosen as sources for biological degradation of aflatoxins[6,18].
Earlier research conducted in our laboratory focused on isolation of sixty nine GRAS Bacillus spp. isolates with consistent in vitro anti- Salmonella spp., Clostridium spp., and Campylobacter spp. activity [8–14]. In the present study, three out of those sixty nine DFM candidates previously evaluated, in addition showed ability to biodegrade AFB1 in vitro, based on growth as well as reduction of fluorescence and area of clearance around each colony (data not shown). Analysis of 16S DNA identified the strains as B. amyloliquefaciens, B. megaterium and B. subtilis; all three were considered GRAS organisms. Furthermore, their physiological properties, tolerance to acidic conditions and high osmotic pressure and relative tolerance to bile salts make them suitable candidates as DFM. In the present study, in vitro digestion time was not enough to confirm biodegradation of AFB1. Further studies to evaluate the possible biodegradation effects of the Bacillus-DFM when continuously administered in broiler chickens feed contaminated with AFB1, are in progress.
Conflict of interest statement
We declare that we have no conflict of interest.
Acknowledgments
This article was supported by the Autogenous Vaccine Research Project of the Poultry Health Laboratory, Poultry Science Department, University of Arkansas.
This article was originally published in Asian Pacific Journal of Tropical Biomedicine, Volume 5, Issue 9, September 2015, Pages 702–706. https://doi.org/10.1016/j.apjtb.2015.07.014. This is an Open Access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
References
[1] Smith RB Jr, Griffin JM, Hamilton PB. Survey of aflatoxicosis in farm animals. Appl Environ Microbiol 1976; 31(3): 385-8.
[2] Abramson D, Mills JT, Marquardt RR, Frohlich AA. Mycotoxins in fungal contaminated samples of animal feed from western Canada, 1982–1994. Can J Vet Res 1997; 61(1): 49-52.
[3] Greco MV, Franchi ML, Rico Golba SL, Pardo AG, Pose GN. Mycotoxins and mycotoxigenic fungi in poultry feed for foodproducing animals. Sci World J 2014; http://dx.doi.org/10.1155/ 2014/968215.
[4] Yunus AW, Razzazi-Fazeli E, Bohm J. Aflatoxin B(1) in affecting broiler's performance, immunity, and gastrointestinal tract: a review of history and contemporary issues. Toxins (Basel) 2011; 3(6): 566-90.
[5] Ciegler A, Lillehoj EB, Peterson RE, Hall HH. Microbial detoxi- fication of aflatoxin. Appl Microbiol 1996; 14(6): 934-9.
[6] Farzaneh M, Shi ZQ, Ghassempour A, Sedaghat N, Ahmadzadeh M, Mirabolfathy M, et al. Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control 2012; 23(1): 100-6.
[7] Khan FA, Zahoor M. In vivo detoxification of aflatoxin B1 by magnetic carbon nanostructures prepared from bagasse. BMC Vet Res 2014; 10: 255.
[8] Wolfenden RE, Pumford NR, Morgan MJ, Shivaramaiah S, Wolfenden AD, Tellez G, et al. Evaluation of a screening and selection method for Bacillus isolates for use as effective direct-fed microbials in commercial poultry. Int J Poult Sci 2010; 9(4): 317-23.
[9] Wolfenden RE, Pumford NR, Morgan MJ, Shivaramaiah S, Wolfenden AD, Pixley CM, et al. Evaluation of selected direct-fed microbial candidates on live performance and Salmonella reduction in commercial turkey brooding houses. Poult Sci 2011; 90(11): 2627-31.
[10] Shivaramaiah S, Pumford NR, Morgan MJ, Wolfenden RE, Wolfenden AD, Torres-Rodr´iguez A, et al. Evaluation of Bacillus species as potential candidates for direct-fed microbials in commercial poultry. Poult Sci 2011; 90(7): 1574-80.
[11] Tellez G, Pixley C, Wolfenden RE, Layton SL, Hargis BM. Probiotics/direct fed microbials for Salmonella control in poultry. Food Res Int 2012; 45(2): 628-33.
[12] Menconi A, Morgan MJ, Pumford NR, Hargis BM, Tellez G. Physiological properties and Salmonella growth inhibition of probiotic Bacillusstrains isolated from environmental and poultry sources. Int J Bacteriol 2013; http://dx.doi.org/10.1155/2013/958408.
[13] Latorre JD, Hernandez-Velasco X, Kallapura G, Menconi A, Pumford NR, Morgan MJ, et al. Evaluation of germination, distribution, and persistence of Bacillus subtilis spores through the gastrointestinal tract of chickens. Poult Sci 2014; 93(7): 1793-800.
[14] Latorre JD, Hernandez-Velasco X, Kogut MH, Vicente JL, Wolfenden R, Wolfenden A, et al. Role of a Bacillus subtilis directfed microbial on digesta viscosity, bacterial translocation, and bone mineralization in turkey poults fed with a rye-based diet. Front Vet Sci 2014; http://dx.doi.org/10.3389/fvets.2014.00026.
[15] Gilliland SE, Staley TE, Bush LJ. Importance of bile tolerance of Lactobacillus acidophilus used as dietary adjunct. J Dairy Sci 1984; 67(12): 3045-51.
[16] Zhao S, Deng L, Hu N, Zhao B, Liang Y. Cost-effective production of Bacillus licheniformis using simple netting bag solid bioreactor. World J Microbiol Biotechnol 2008; 24(12): 2859-63.
[17] Bedford MR, Classen HL. An in vitro assay for prediction of broiler intestinal viscosity and growth when fed rye-based diets in the presence of exogenous enzymes. Poult Sci 1993; 72(1): 137-43.
[18] Gao X, Ma Q, Zhao L, Lei Y, Shan Y, Ji C. Isolation of Bacillus subtilis: screening for aflatoxins B1, M1, and G1 detoxification. Eur Food Res Technol 2011; 232(6): 957-62.