Introduction
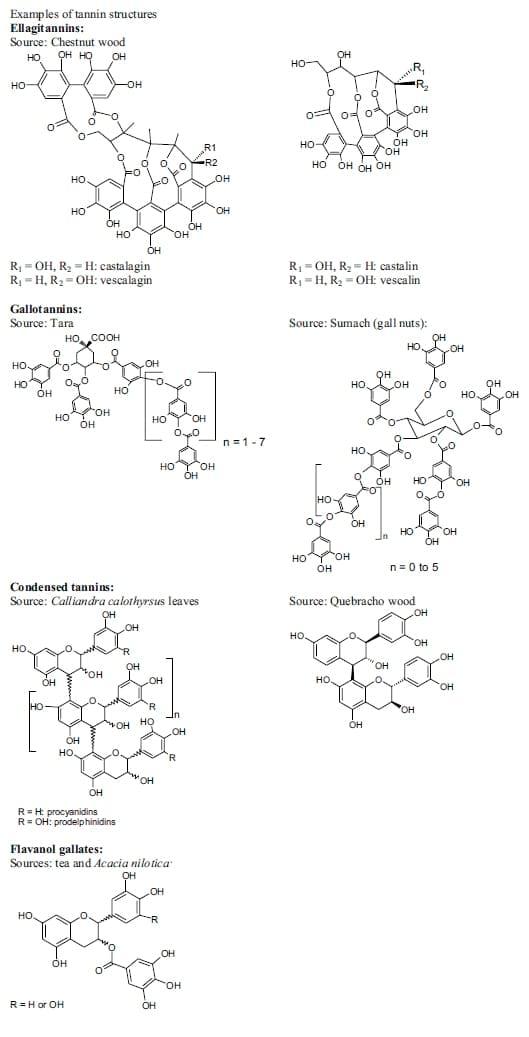
Tannins, polyphenolic compounds which occur naturally in the bark, leaves, seeds and roots of many plants 1. They encompass a wide range of molecular weights ranging from 500 to over 20,000 Daltons 2 and confer the astringent taste often found in tea, chocolate, wine and fruits 1, 3. Due to their large number of phenolic groups and aromatic rings they readily form complexes via hydrophilic and hydrophobic interactions with proteins, polysaccharides, polyethylene glycol and lipids 4, 5. Hydrolysable (HT) and condensed tannins (CT) form the two main tannin classes; however, some tannins contain the structural features of both classes (e.g. flavanol gallates, Fig. 1) 2, 6, 7.
The antimicrobialproperties of tannins are of interest in several different fields, such as food science, wood science, soil science, plant pathology, pharmacology, human and animal nutrition 8. In animal husbandry, high dietary concentrations of tannins are known to reduce the digestibility of the fodder and adversely to affect ruminant health 9. However, at moderate concentrations tannins can reduce the risk of bloating, increase the uptake of essential amino acids and proteins, enhance the production of milk and wool, and be effective against gastrointestinal parasites 10, 11.
The present study determined in vitro the effect of different types of tannins on the growth and survival of the Gram-negative pathogen Salmonella Typhimurium. Since January 2006, antibiotics have been banned in routine animal production systems. Their use as growth promoters has led to an increased occurrence of antibiotic-resistant bacteria in farm animals and colonisation of human consumers by antibiotic-resistant bacteria 12. Antibiotic-resistant Salmonella has been detected in retail meats 13. Thus, Salmonella is of concern for animal welfare since resistance in animals can lead to economic losses through death or poor performance of infected animals, but also as a public health concern from transmission of Salmonella to humans via meat. An understandingof alternative strategies, to antibiotics, is needed for preventing or suppressing sub-clinical Salmonella infection in farm animals. An in vitro study was carried out to test the ability of various tannins to suppress Salmonella growth in order to identify candidate feed additives.
Materials and Methods
Test materials: The following commercial tannin products were provided by Silvateam (San Michele Mondovi, Italy): extracts from chestnut wood (75.2% tannin content), chestnut wood (91.6%), quebracho (77.5%), quebracho (Tannino QS-SOL, 92.1%), tara (95.1%), Chinese galls (sumach) (94.0%) and green tea (84.9%), (Table 1). Tannins were used either as received or subjected to column chromatography on Sephadex LH-20 (Amersham Pharmacia, Uppsala, Sweden) and eluted with acetone/water (7:3, v/v) (described here as 'tannin fraction') 14. Acetone was evaporated below 35°C and the remaining water phase freezedried. Tannins from the leaves of Calliandra calothyrsus (San Ramón and Patulul accessions) and the fruits of Acacia nilotica were isolated and also purified on Sephadex LH-20 as described by Stewart et al. 14.
Figure 1. Tannins from chestnut wood (Castanea sativa), quebracho (Schinopsis sp), tara (Caesalpinia spinosa), gall nuts of sumach (Rhus coriaria), tea (Camelia sinensis), Calliandra calothyrsus, Acacia nilotica
Bacterial strains and culture conditions: A stock culture collection was maintained at -70ºC in 15% (w/w) glycerol. Salmonella Typhimurium strain SL1344nalr was a nalidixic acid- resistant derivative of the wild-type strain SL1344, a gift from the Department of Food and Environmental Safety (VLA, Veterinary Laboratories Agency, Weybridge, UK). SL1344nalr was cultured for 24 h in 10 ml of Luria-Bertani broth (Difco Laboratories, Detroit, USA) in a shaking incubator (150 rpm) or in Brilliant Green agar (BGA; Oxoid, Basingstoke, Hampshire, UK: proteose peptone 10.0 g l-1; yeast extract 3.0 g l-1; lactose 10.0 g l-1; sucrose 10.0 g l-1; sodium chloride 5.0 g l-1; phenol red 0.08 g l-1; brilliant green 0.0125 g l-1; agar 12.0 g l-1; [pH 6.9 ± 0.2]) including nalidixic acid (15μg ml-1) to facilitate enumeration. These mutations did not affect virulence of the strains. Selenite Cystine broth (Difco Laboratories, Detroit, USA) was used as the liquid growth medium due to its low protein content. Other growth media such as glucose-free MRS (De Man, Rogosa, Sharpe) medium (Oxoid: bacteriological peptone 10 g l-1; "Lab- Lemco" meat extracts 8 g l-1; yeast extract 4 g l-1; dipotassium hydrogenated phosphate 2 g l-1; sodium acetate 5 g l-1; triammonium citrate 2 g l-1; MgSO4 0.2 g l-1; MnSO4 0.05 g l-1; Tween80 1 ml; cysteine HCl 0.5 g l-1; rezasurine 4 ml) and buffered peptone water (Oxoid), were also tested.
Anti-microbial activity: Antimicrobial activity of tannins on selected bacterial strains was measured in liquid culture by a plate count method and disc diffusion assay according to Puupponen-Pimiä et al. 15. Briefly, 10 ml of fresh selenite cystine growth medium was inoculated with 1% (v/v) of an overnight culture (107 CFU ml-1;). Tannins were added to the culture medium to give final concentrations of 1, 3 and 6 mg ml-1. The experimental set up consisted of 1 ml aqueous tannin solution of one of the three different concentrations and 107 CFU ml-1 of an overnight culture of Salmonella Typhimurium in 10 ml selenite broth. The cultures were shaken well and incubated at 37°C for 24 h. The positive control contained only 107 CFU ml-1 of Salmonella Typhimurium but no tannins. Selenite broth containing tannins (1 ml) without Salmonella served as negative controls. This was to ensure that no growth took place due to contamination and to observe whether any precipitate was formed when the tannins came into contact with the growth medium.
Bacterial growth was followed by taking samples from the cultures during an incubation period of 24 h at times 0, 5, 10 and 24 h. Culture pH and optical density at 600 nm (OD 600) were measured for all time points.
Table 1. List of tannins investigated and their known antibacterial properties.
Plate count method: The sample of the experimental sets and the positive control at times 0, 5, 10 and 24 h were plated onto BGA with nalidixic acid (BGA+nal) at different dilutions (from 102 to 108 CFU ml-1) in triplicate for each time point to measure bacterial growth. Dilutions were prepared using phosphate buffered saline (PBS).
The specific growth rate was calculated using the loge of microbial count results of time 0 and 10 h from the plating experiments. This time period represents exponential growth. When the stationary phase was reached before 10 h, time 0 and 5 h were used instead. The decrease in Salmonella counts after 24 h was calculated by subtracting the counts of Salmonella in presence of tannins from those in absence of tannins (positive control).
Diffusion assay: Of overnight bacterial culture 80 ml was diluted to 105 CFU ml-1 and transferred onto a BGA+nal plate. Sterile paper discs (6 mm diameter) prepared from Whatman Grade Number 17 filter paper (Fisher Scientific UK Ltd., Loughborough, UK) were dipped into the tannin solutions and placed on the plate containing the inoculum. A maximum of three discs was placed per plate. Inhibition zones were measured and recorded after 24 hours incubation. First, the diameter of the whole disc (i.e. disc plus inhibition zone) was measured from 3-4 different sites and then 0.6 cm was subtracted from the average to obtain the diameter of the inhibition zone around the disc.
Only tannins that led to a lower specific growth rate compared to the control and which showed good reduction in Salmonella counts after 24 h with the plate count method, were used for testing in the disc diffusion assay.
Statistical analysis: The significance of differences was tested using the parametric analysis of variance (ANOVA). The SAS statistical package (version 6.12) was used to perform statistical analyses. Differences were considered to be statistically significant when the P value was below 0.05.
Results
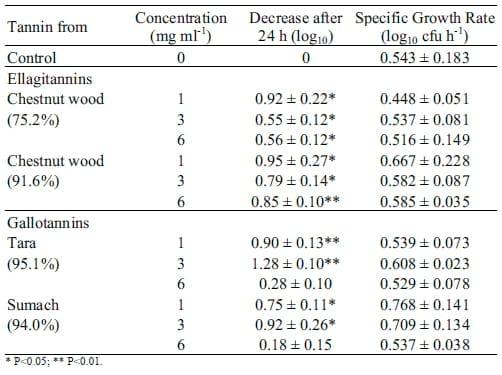
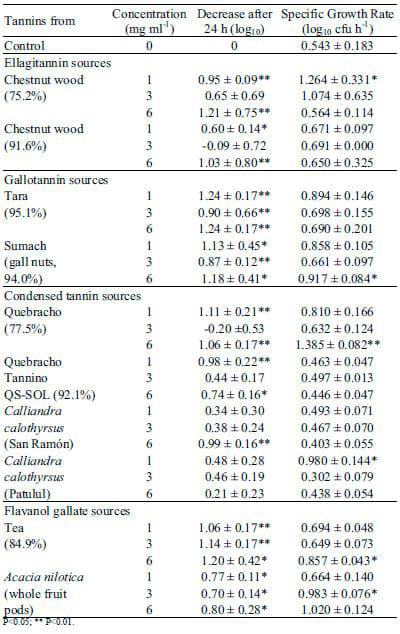
Plate count method: The colour of the tannin solutions was too dark for optical density measurements to be recorded accurately making it necessary to follow bacterial growth by using the plate count method instead. The tested tannins clearly reduced Salmonella numbers in our in vitro approach. As shown in Tables 2 and 3 all commercial tannins and most tannin fractions significantly decreased Salmonella counts compared to the positive control (P<0.05). Two of the tannin fractions from chestnut 91.6% and quebracho (77.5%) did not inhibit Salmonella growth at a concentration of 3 mg ml-1 however, they successfully reduced Salmonella at 1 or 6 mg ml-1. The observed suppression of Salmonella numbers was independent from a reduction in specific growth rates of Salmonella. Apart from some tannins which actually increased the growth rate of Salmonella – probably due to impurities of the tested products – the specific growth rate of Salmonella in presence of tannins did not differ from the specific growth rate without addition of tannin. The final bacterial count for Salmonella in presence of most tannins was significantly lower compared to Salmonella in broth alone (positive control).
Table 2. Influence of commercial tannins on Salmonella counts
Commercial gallotannins performed slightly better than the commercial ellagitannins in reducing bacterial counts. However, commercial gallotannins were only active at concentrations of 1 and 3 mg ml-1 and not at 6 mg ml-1 (P<0.01) (Table 2), while the tara and sumach tannin fractions were also active 6 mg ml-1. For these gallotannin fractions, reduction was usually greater than 1 log unit after 24 h.
Both, the quebracho profisetinidin and prorobinetinidin tannins, obtained either from the 77.5% or the 92.1% commercial preparations, significantly inhibited Salmonella. The tannins extracted from Calliandra calothyrsus were relatively ineffective. In contrast, flavanol gallates from green tea or Acacia nilotica showed a significant antimicrobial effect at all concentrations.
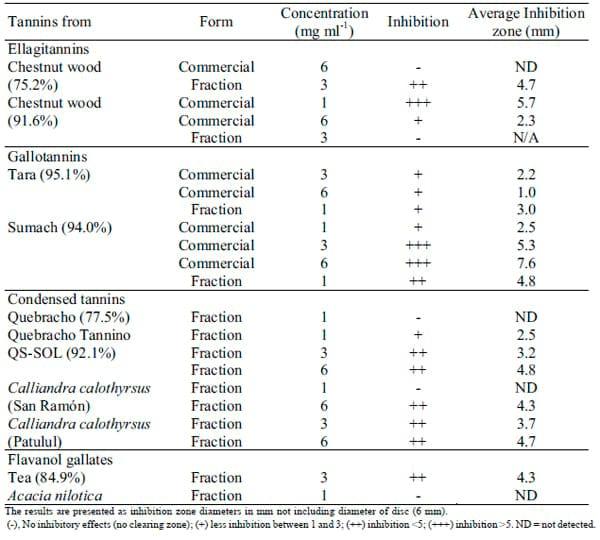
Disc diffusion assay: The disc diffusion assay was used to confirm antimicrobial activity of the tannins. Most of the tannins showed good reduction in Salmonella numbers with the plate count method but not all of these displayed an inhibition zone with the disc diffusion assay. All commercial tannins with the exception of the 75.2% chestnut extract showed inhibition zones. Sumach gave the best results with inhibition zones increasing in a dosedependent manner for the commercial extract and its tannin fraction gave very high inhibition with only 1 mg ml-1. However, for most of the fractionated tannins the lowest concentration (1 mg ml-1) failed to show inhibition in the disc diffusion assay (Table 4).
Table 3. Influence of isolated tannin fractions on Salmonella counts.
Surprisingly, there was no correlation between the appearance of inhibition zones and the decrease in log 10 values after 24 h.
Discussion
Despite their structural diversity, all tested tannins showed antimicrobial effects against Salmonella Typhimurium either in liquid culture or in the diffusion assay. The decrease of Salmonella Typhimurium counts after 24 h was not associated with an inhibition of growth rate in the logarithmic phase. Surprisingly, in several of our experiments there was no concentration related effect. There are two potential explanations for this. First, since these commercial tannin extracts are not pure, sugars or other constituents may have facilitated bacterial growth, or a more likely explanation is that some tannins precipitated at higher concentrations and were thus unable to act directly on Salmonella. Indeed, such precipitates were observed in our study e.g. with commercial sumach tannins at 6 mg ml-1 but not at 3 mg ml-1. While there were no differences in antimicrobial activity between the commercial product and the purified fractions for ellagitannins, fractions of tara and sumach preparations were more effective than their commercial precursors. This might be due to a higher concentration of active tannins. Previous work has suggested that lower molecular weight tannins exerted better anti-microbial properties than higher molecular weight tannins 16, 17. Another reason could be that high molecular weight tannins still present in the commercial extracts had weaker anti-microbial properties 18.
The mechanisms by which tannins inhibit bacteria have not yet been fully elucidated, but are probably related to several different factors such as (i) the number of ortho-phenolic groups that can chelate metal ions, (ii) the number of ortho-phenolic groups and aromatic rings that can bind substrates, (iii) the lipophilicity and molecular weight that determine the extent of interaction with bacterial membranes 7, 8, 19.Moreover, Gram-negative bacteria were less sensitive than Gram-positive bacteria in studies with green tea flavanols due to the presence of negatively charged lipids 20. Lipopolysaccharides are characteristic of the membranes of Gramnegative bacteria and interact with both condensed and hydrolysable tannins 16, 21, 22. Our results confirm that condensed, hydrolysable (gallo- and ellagitannins) and monomeric flavanol gallate tannins showed activity against pathogenic Salmonella Typhimurium. Gallotannins, in particular from sumach, seemed to be the most effective in both assays as crude product as well as their fractions. Interestingly, Rhus coriaria (sumach) berries are used in folk medicine against diarrhea and are sprinkled over kebabs and grilled meat in Mediterranean regions 23.Nasar-Abbas et al. 23 found that aqueous sumach berry extracts at a concentration of 10 mg ml-1 achieved 1.8 to 5.7 log10 reductions against various food-borne bacteria, including Salmonella enteritidis. At 5 mg ml-1 they observed a decrease of 0.97 log10 values for Salmonella enteritidis which corresponds to the values reported here for 3 mg ml-1of commercial as well as 3 mg ml-1 and 6 mg ml-1sumach tannin fraction.
Inhibition zones of 1.3- 9.7 mm (excluding the diameter of the cup) have been reported by Nasar-Abbas et al. 23 for Salmonella enteritidis for concentrations of 5 to 50 mg ml-1 of aqueous sumach extracts . Similar values have also been reported for the inhibition of mastitis-causing Staphylococcus aureus by sumach and quebracho at concentrations of 2 to 8 mg ml-124. We also found a dose-dependent increase in inhibition zones for commercial sumach extract and for the tannin fraction from quebracho (Tannino QS-SOL; 92.1%). It would appear that both tannin source and concentration are important factors in the inhibition of bacterial growth 24. Therefore, the failure of some of the tannin extracts to produce inhibition in the disc diffusion assay might be due to relatively low concentrations. Shanmugavelu et al. 25 reported that the minimum inhibitory concentration against Clostridium perfringens of grape seed, mimosa and quebracho tannin extracts was 10, 10 and 5-10 mg ml-1, respectively. Toda et al. 26 found inhibition zones of 4-8 mm and < 4 mm against Salmonella Typhimurium TSA-2121 with aqueous extracts of 200 mg ml-1 of black and green tea. However, the work described here achieved comparable inhibition at much lower concentrations (3 mg ml-1). This may suggest that the preparation method of tannin-rich extracts might be another important factor for ensuring antimicrobial activity.
Table 4. Inhibition zone for commercial tannins and tannin fractions measured after 24 hours by disc diffusion assay.
Apart from factors that determine the in vitro activity of tannins a number of other mechanisms are likely to contribute to their in vivo activities in animal health. Although some bacteria can develop tolerance mechanisms against tannins by secreting extracellular polysaccharides (EPS) 27, it may not be necessary to achieve complete inhibition of pathogenic Salmonella in order for tannins to produce beneficial health effects. Tannins that prevent adhesion, cause bacterial aggregation, damage bacterial membranes or delay bacterial growth may prove sufficiently effective, as their action may provide enough time to eliminate pathogenic bacteria and for the host to develop its immune response 16, 28, 29. The inclusion of low levels of quebracho and chestnut tannins in the diets of black rhinoceros and steers significantly decreased, respectively, the number of Enterobacteriaceae spp. 30 and E. coli 31 in the faeces.
Our study demonstrated for the first time that condensed, hydrolysable and flavanol-3-gallate tannins had significant effects against a pathogenic Salmonella strain and may therefore provide alternatives to antibiotics. The inhibitory spectrum depended on the particular type of tannins and achieved up to 1.28 log10 reduction after 24 hours. Sumach gallotannins appear to have the best potential as an effective feed additive.
Acknowledgements
We thank Silvateam (San Michele Mondovi, Italy) for providing the commercial tannin products used in the experiments.
References
1. Haslam, E. 1981. Vegetable tannins. In Stumpf, P. K. and Conn, E. E. (eds). The Biochemistry of Plants. Vol. 7, Academic Press, New York, pp. 527-556.
2. Khanbabaee K. and Van Ree, T. 2001. Tannins: Classification and definition. Nat. Prod. Rep. 18:641-649.
3. Cowan, M. 1999. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 12:564-582.
4. Mueller-Harvey, I. 2006. Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 86:2010-2037.
5. Uekusa, Y., Kamihira, M. and Nakayama, T. 2007. Dynamic behavior of tea catechins interacting with lipid membranes as determined by NMR spectroscopy. J. Agric. Food Chem. 55:9986-9992.
6. Mueller-Harvey, I. 2001. Analysis of hydrolysable tannins. Anim. Feed Sci. Technol. 91:3-20.
7. Schofield, P., Mbugua, D. M. and Pell, A. N. 2001. Analysis of condensed tannins: A review. Ani. Feed Sci. Techn. 91:21-40.
8. Scalbert, A. 1991. Antimicrobial properties of tannins. Phytochem. 30:3875-3883.
9. Min, B. R., Barry, T. N., Attwood, G. T. and McNabb, W. C. 2003. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol. 106:3- 19.
10. Niezen, J. H., Charleston, W. A. G., Robertson, H. A., Shelton, D., Waghorn, G. C. and Green, R. 2002. The effect of feeding sulla (Hedysarum coronarium) or lucerne (Medicago sativa) on lamb parasite burdens and development of immunity to gastrointestinal nematodes. Vet. Parasitol. 105:229-245.
11. Paolini, V., Fouraste, I. and Hoste, H. 2004. In vitro effects of three woody plant and sainfoin extracts on 3rd stage larvae and adult worms of three gastrointestinal nematodes. Parasitology 129:69-77.
12. Van den Bogaard, A. E. and Stobberingh, E. E. 2000. Epidemiology of resistance to antibiotics. Links between animals and humans. Int. J. Antimicrobial Agents 14:327-335.
13. White, D. G., Zhao, S., Sudler, R., Ayers, S., Friedman, S., Chen, S., McDermott, P. F., McDermott, S., Wagner, D. D. and Meng, J., 2001. The isolation of antibiotic-resistant Salmonella from retail ground meats. N. Engl. J. Med. 345:1147-1154.
14. Stewart, J. L., Mould, F. and Mueller-Harvey, I. 2000. The effect of drying treatment on the fodder quality and tannin content of two provenances of Calliandra calothyrsus Meissner. J. Sci. Food Agric. 80:1461-1468.
15. Puupponen-Pimiä, R. N. L., Meier, C., Kähkönen, M., Heinonen, M., Hopia, A. and Oksman-Caldentey, K. M. 2001. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 90:494-507.
16. Funatogawa, K. H. S., Shimomura H., Yoshida T., Hatano T., Ito, H. and Hirai, Y. 2004. Antibacterial activity of hydrolyzable tannins derived from medicinal plants against Helicobacter pylori. Microbiol. Immunol. 48:251-261.
17. Cross, D. E., Brown, R., Mueller-Harvey, I., Mackey, B. M. and Kelly, A. F. 2006. The antimicrobial effect of tanniniferous plant extracts in vitro against Campylobacter spp., in assays involving both solid and liquid growth media. World Poultry Science Association, Annual Conference, York, British Poultry Abstract 2:24-25.
18. Sivakumaran, S. M. A., Meagher, L. P., Kolb, B., Foo, L. Y., Lane, G. A., Attwood, G. A., Fraser, K. and Tavendale, M. 2004. Variation in antimicrobial action of proanthocyanidins from Dorycnium rectum against rumen bacteria. Phytochemistry 65:2485-97.
19. Kajiya, K., Hojo, H., Suzuki, M., Nanjo, F., Kumazawa, S. and Nakayama, T. 2004. Relationship between antibacterial activity of (+)-catechin derivatives and their interaction with a model membrane. J. Agric. Food Chem. 52:1514-1519.
20. Ikigai, H., Nakae, T., Hara, Y. and Shimamura, T. 1993. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta 1147:132- 136.
21. Delehanty, J. B., Johnson, B. J., Hickey, T. E., Pons, T. and Ligler, F. S. 2007. Binding and neutralization of lipopolysaccharides by plant proanthocyanidins. J. Nat. Products 70:1718-1724.
22. Johnson, B. J., Lin, B., Dinderman, M. A., Rubin, R. A., Malanoski, A. P. and Ligler, F. S. 2008. Impact of cranberry on Escherichia coli cellular surface characteristics. Biochem. Biophys. Res. Commun. 377:992-994.
23. Nasar-Abbas, S. M. and Halkman, A. K. 2004. Antimicrobial effect of water extract of sumac (Rhus coriaria L.) on the growth of some food borne bacteria including pathogens. Int. J. Food Microbiol. 97:63-9.
24. Min, B. R., Pinchak, W. E., Merkel, R., Walker, S., Tomita, G. and Anderson, R. C. 2008. Comparative antimicrobial activity of tannin extracts from perennial plants on mastitis pathogens. Scientific Research and Essay 3:66-73.
25. Shanmugavelu, S., Ruzickova, G., Zrustova, J. and Brooker, J. D. 2006. A fermentation assay to evaluate the effectiveness of antimicrobial agents on gut microflora. J. Microbiol. Meth. 67:93-101.
26. Toda, M., Okuda, S., Hiyoshi, R. and Shimamura, T. 1989. The bactericidal activity of tea and coffee. Appl. Microbiol. 8:123-125.
27. McSweeney, C. S., Palmer, B., Bunch, R. and Krause, D. O. 2001. Effect of the tropical forage Calliandra on microbial protein synthesis and ecology in the rumen. J. Appl. Microbiol. 90:78-88.
28. Mueller-Harvey, I., McAllan, A. B., Theodorou, M. K. and Beever, D. E. 1988. Phenolics in fibrous crop residues and plants and their effects on the digestion and utilisation of carbohydrates and proteins in ruminants. In Reed, J. D., Capper, B. S. and Neate, P. J. H. (eds). Plant Breeding and the Nutritive Value of Crop Residues. Proceedings of a workshop held at ILCA, Addis Ababa, Ethiopia, 7-10 December 1987. ILCA, Addis Ababa, pp. 97-132.
29. Foo, L. Y., Howell, A. B. and Vorsa, N. 2000. The structure of cranberry proanthocyanidins which inhibit adherence of urophatogenic Pfimbirated Escherichia coli in vitro. Phytochemistry 54:173-181.
30. Clauss, M., Wittenbrink, M. M., Castell, J. C., Kienzle, E., Dierenfeld, E. S., Flach, E. J., Macgregor, S. K., Hoppe, T., Hummel, J., Streich, W. J. and Hatt, J.-M. 2007. Quantification of enterobacteriaceae in faeces of captive black rhinoceros (Diceros bicornis) in relation to dietary tannin supplementation. J. Anim. Physiol. Anim. Nutr. 92:29- 34.
31. Min, B., Pinchak, W., Anderson, R. and Callaway, T. 2007. Effect of tannins on the in vitro growth of Escherichia coli O157:H7 and in vivo growth of generic Escherichia coli excreted from steers. J. Food Prot. 70:543-550.
32. Li, Y. and Song, G. 2004. Study of bacteriostasis of chestnut shell extract. Chem. Ind. Forest Prod. 24:61-64.
33. Takaishi, Y., Kondon, K., Shibata, H. and Higuti, T. 2004. ILSMRs (intensifier of β-lactam-susceptibility in methicillin-resistant Staphylococcus aureus) from tara [Caesalpinia spinosa (Molina) Kuntze]. Phytomedicine 13:209-212.
34. Özcan, M. 2003. Effect of sumach (Rhus coriaria L.) extracts on the oxidative stability of peanut oil. J. Medicinal Food 86:2010-2037.
35. Lauk, L., Caccamo, F., Speciale, A. M., Tempera, G., Ragusa, S. and Pantè, G. 1998. Anti-microbial activity of Rhus coriaria L. leaf extract. Phytotherapy Res. 12:S152-S153.
36. Hervas, G., Frutos, P., Javier Giraldez, F., Mantecon, A.R. and Alvarez Del Pino, M.C. 2003. Effect of different doses of quebracho tannins extract on rumen fermentation in ewes. Anim. Feed Sci. Technol. 109:65-78.
37. McSweeney, C. S., Krause, D. O., Smith, W. J. M., Brooker, J. D. and Palmer, B. 2005. Tolerance mechanisms of streptococci to hyrolysable and condensed tannins. Anim. Feed Sci. Technol. 121:59-75.
38. Kaur, K., Husheem, M., Arora, S., Härkönen, P. and Subodh, K. K. 2005. In vitro bioactivity-guided fractionation and characterization of polyphenolic inhibitory fractions from Acacia nilotica (L.). J. Ethnopharmacol. 99:353-360.
39. Eldeen, I. M. S. and Van Staden, J. 2007. Antimycobacterial activity of some trees used in South African traditional medicine. South African J. Bot. 73:248-251.