shrimp health
Nutritional and management strategies to promote shrimp health
The possibilities of applying vaccination to successfully control diseases in shrimp farming appear remote because shrimp seem to possess an immune system that is not capable of adaptive immunity. Antibiotics show little effectiveness in controlling most disease problems. Furthermore, antibiotic use is banned in many countries because of Europe’s low tolerance of antibiotic residues in foods. Consequently, there is no relief in sight for eliminating or significantly reducing disease problems in the shrimp farming industry. However, the industry is learning how to manage the disease problems. Nutrients that enhance and products that activate the immune system form an important part in disease management of shrimp. The objective of this paper is to provide an overview of shrimp health management with particular emphasis on the nutritional and stimulatory aspects of the shrimp immune system.
Shrimp diseases
Farmed shrimp are infected by a range of disease agents including bacteria, viruses, fungi and protozoa. This overview focuses mainly on viral and bacterial diseases that have had a significant impact on the shrimp farming industry. For comprehensive information on disease problems in shrimp farming, please refer to Lightner (1996). There are a number of viruses that infect shrimp, but not all of them cause fatal diseases. Infectious hypodermal and hematopoietic necrosis virus (IHHNV) has been observed in most commercially farmed shrimp species. It appears to be harmless in some species such as the Asian tiger shrimp, Penaeus monodon, but malicious in others causing mortality and growth retardation. There are a number of other viruses such as the monodon baculovirus (MBV), hepatopancreatic parvo-like virus (HPV), and baculovirus penaei (BP) that damage the cells of the hepatopancreas and make the shrimp susceptible to other disease agents. It is believed that infection by these viruses causes a reduction in growth rates.
As noted earlier, the three viruses that cause acutely fatal diseases in shrimp farming are the white spot syndrome virus (WSSV), yellow head virus (YHV) and Taura syndrome virus (TSV). All three viruses can cause extensive mortality within a few days of the first clinical signs of the disease. As discussed below, the severity of a viral disease typically subsides in about two years after the first incidence of the given disease. This apparently indicates some type of an adaptive response to the disease agent. However, the viruses are never completely eliminated. They resurface periodically, particularly at times of stress, to cause large-scale mortalities. Furthermore, growth retardation often coincides with viral infections resulting in economic losses.
YELLOW HEAD VIRUS
Yellow head virus was first reported in Thailand in 1991. A related virus called Gill-Associated Virus (GAV) was reported from Australia in 1996. Yellow head virus caused severe disease outbreaks in Thailand until 1994. The disease typically occurs in juveniles or sub-adults. A spurt in feed consumption followed by loss in appetite, lethargy and erratic swimming are the gross signs first observed. Pale yellow coloration of the gills and cephalothorax is often noted. Mortalities start within a few days and can reach as high as 100% in 3-5 days after the gross signs are observed. Sporadic disease outbreaks still occur, mainly in Asia, but the mortalities are less severe than in the past.
WHITE SPOT SYNDROME VIRUS
White spot syndrome virus was first reported in Japan in 1993, although it might have originated in China. This virus has caused the most damage to the shrimp farming industry. It spread to almost all shrimp farming countries of Asia in a span of three years. It was reported in the United States in 1995, and spread to Central and South American countries in a span of four years. Almost all shrimp species have been affected. Further, most crustaceans can be infected with the virus and become carriers. The characteristic feature of WSSV infection is the presence of white spots or patches under the carapace, although this may not be present in all diseased shrimp. Soon after showing general signs of ill-health such as reduced feed intake and erratic swimming, mortalities occur. Mortality up to 100% may occur within seven days after the first sign of problems. The infection may occur at any stage in the life cycle of the shrimp. Stressful conditions such as sudden changes in environmental conditions, particularly lowered temperatures, trigger disease. Frequent WSSV disease outbreaks still occur worldwide, but there are more and more cases of shrimp populations escaping severe mortality in spite of WSSV infections.
TAURA SYNDROME VIRUS
Taura syndrome was reported first in 1992 in Ecuador. Presence of TSV was reported in 1995. TSV spread throughout the Pacific coast of Central and South America and mainly affected the Pacifc White Shrimp, P. vannamei. Distinguishable gross signs of TSV are pale reddish coloration of the body, red tail fans, necrosis of the cuticular epithelium, and soft shells. Mortality during molting is common. Sometimes, the shrimp are affected only transitionally: gross signs of the disease may occur, but the shrimp may behave and feed normally. While TSV still occurs, the catastrophic losses suffered in the early years of TSV infection are less common now.
VIBRIOSIS
Infection by Vibrio spp. is the most common bacterial disease problem in shrimp culture. Vibrio spp. are ubiquitous and naturally present in most aquatic ecosystems. Infections occur when shrimp are stressed or unhealthy. Infections may also occur as a result of high concentrations of Vibrio spp. in the culture system. Some species and strains, particularly V. harveyi, are more infectious than others. Shell lesions, black coloration of gills and discoloration of shells occur as a result of vibriosis. Severe mortalities may follow acute infections. Chronic infections may result in erratic swimming behavior, abnormal coloration, external fouling and less severe, but sustained mortalities.
Shrimp immune system
An understanding of shrimp immune defense is just beginning to emerge. Bachère (2000) provided an excellent summary of the defense systems used by shrimp in the event of infection. However, as Flegel (2001) pointed out, most of our knowledge of shrimp immunity is based on studies of bacterial and fungal infections. The author further argued that shrimp response to viral infections is different from that of bacterial infections.
Shrimp have an open circulatory system and there is no separation between the circulatory and lymphatic systems. The fluid within this open system is called hemolymph. Hemocytes are cells present in the hemolymph. These cells play a major role in the immune response of shrimp to bacterial and fungal infections. The shrimp’s immune system identifies invasion by means of specific cell wall components of the invading organisms.
Peptidoglycans and lipopolysaccharides on bacterial cell walls and betaglucans on fungal cell walls are the molecules that are readily identified by specific molecules in the hemolymph. Once such a pattern recognition molecule binds to its specific molecular counterpart on the foreign body, a number of hemocyte-mediated responses follow to clear the invading organisms. These responses include agglutination, phagocytosis, and production of free radicals and antimicrobial compounds. A number of studies have shown that the shrimp immune system can be stimulated with purified peptidoglycans, lipopolysaccharides or betaglucans to achieve generalized immune protection against bacterial infections. Live or inactivated bacteria or yeast may also offer protection against bacterial infections. These aspects will be discussed later in this article.
It is generally believed that invertebrates are incapable of adaptive immune response. This is based on the fact that inducible humoral compounds such as immunoglobulins, T cell receptors, the Major Histocompatibility Complex and memory T-cells are not present in invertebrates. However, citing the presence of adhesion molecules belonging to the super family of immunoglobulins and a few other observations, Arala-Chaves and Sequeira (2000) postulated that invertebrates might possess a unique form of adaptive immune response.
As discussed earlier, the severity of a viral disease typically subsides in about two years after the first incidence of the given disease. Meticulously documenting the history of past viral epizootics worldwide, Flegel (2001) noted that all major viral disease problems resulted in catastrophic crop losses in the first two years, but mortality became less severe afterwards. The virus was never eliminated, but persistent infections occurred. In many cases, the infected shrimp did not show any sign of disease and survived to harvest. The apparent protection from the disease was applicable only for those shrimp previously exposed to the given virus. Shrimp populations previously unexposed to the virus suffered severe mortality. There was no crossprotection, i.e. shrimp populations that enjoyed a degree of protection against a certain virus due to previous exposure were susceptible to disease due to another virus to which they had not been exposed. All these observations point to the possibility that a specific and memory-based viral defense mechanism exists in shrimp.
Pointing out that viral infections do not result in hemocyte aggregation and subsequent generalized cell-based immune responses, Flegel (1997) suggested that shrimp die from viral infections due to large-scale occurrence of programmed cell death (apoptosis). He further proposed that prior exposure to a virus resulted in specific memory that suppressed apoptosis upon subsequent infections (Flegel, 2001). This has been termed as the ‘viral accommodation’ theory.
If specific and memory-based viral defense mechanisms are present in shrimp as speculated by Arala-Chaves and Sequeira (2000) and Flegel (2001), possibilities of developing compounds to prevent mortalities due to viral infections exist. Note that repeated infections still occur in shrimp previously exposed to a virus, but the infected shrimp do not succumb to diseases due to the defense mechanism. The infections impose an energy cost on the shrimp. The generally declining performance of P. monodon stocks in Asia is now attributed to the persistence of one or more viruses in the stocks. Furthermore, the protection offered by previous viral exposure is limited. Stressful situations, particularly low dissolved oxygen, low temperature or a sudden change in pH, increase the vulnerability of shrimp to WSSV or YHV even now.
Health management in shrimp culture
It is clear from the above discussion that the prospects for prevention or treatment of viral infections are dim. Reliable methods to prevent infections from becoming fatal disease outbreaks or retarding growth do not exist. Therefore, the first step in health management of shrimp is enforcement of stringent biosecurity measures that prevent the entry of pathogens into the culture system. Other important components of shrimp health management include minimization of stressful events and use of compounds that boost and/or stimulate the shrimp immune system. Essentially, these components relate to the three elements, namely host, pathogen and environment, in the classic epidemiological triad proposed by Snieszko (Figure 1). For a comprehensive review of concepts and practices related to shrimp health management, please see Fegan and Clifford (2001). Following are key recommended practices for effective shrimp health management.
1. DISINFECTION OF POND AND WATER PRIOR TO STOCKING
Disinfecting pond bottoms after every culture cycle is required to eradicate pathogens and carriers of pathogens. The pond should be completely drained and thoroughly dried or treated with oxidizing agents. Organic waste accumulated on the pond bottom should also be removed.
Water used to fill ponds also requires disinfection. First, the water should be filtered through a series of screens. The last screen should have 250 micron or smaller size mesh openings. The screening is meant to filter out carriers of pathogens. The water is then treated with chlorine to kill pathogens and/ or carriers that might have slipped through the filters. Since viruses do not survive outside of a host for more than a few days, the water should be allowed to stand for 5-7 days before stocking.
2. STOCKING HEALTHY LARVAE
It is very important that the larvae stocked in ponds are free of viruses. Withyachumnarnkul (1999) showed that ponds stocked with shrimp carrying WSSV are 50 times more likely lose a crop compared to those stocked with shrimp free of WSSV. Therefore, larvae should be screened for known viral diseases before purchase. General health of the larvae should also be ascertained prior to stocking.
Specific Pathogen Free (SPF) shrimp stocks are available in some countries. These stocks go through a set of stringent screening, isolation and biosecurity protocols to ensure that the larvae are free of one or more specific viruses. Wider availability and use of these stocks will greatly reduce the likelihood of diseases in the future.
3. EXCLUSION OF CARRIERS
Other crustaceans likely to occur in shrimp culture areas, particularly crabs, are prone to viral infections. These organisms may infect healthy shrimp. Predatory and scavenging birds living in shrimp culture areas have always been considered prime suspects in the spread of WSSV. Efforts to completely eliminate crabs entering shrimp ponds or bird predation have not succeeded to a large extent because of practical implementation issues.
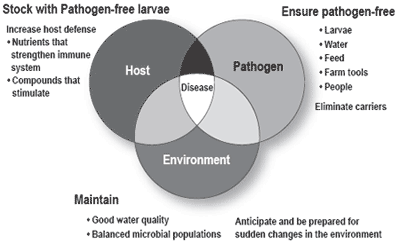
Figure 1. Snieszko’s ‘epidemiological triad’ and its relevance to shrimp health management.
However, it is possible to eliminate carriers during the pond preparation stage by use of chlorine or insecticides.
4. ENVIRONMENTAL MANAGEMENT TO MINIMIZE STRESS
Sudden changes in water quality trigger viral diseases in shrimp. Particularly critical are dissolved oxygen, temperature, salinity and pH. High mortalities due to viral disease are associated with heavy rainfall in many countries.
Maintaining pH at stable levels (preferably at 8- 9) with the use of lime has been shown to reduce disease incidence in some Asian farms. Fegan and Clifford (2001) recommended a minimum dissolved oxygen level of 4 ppm in shrimp ponds. Vidal et al. (2001) clearly established the relationship between WSSV incidence and water temperature. In their study, 100% mortality occurred in WSSV-infected shrimp maintained at 25oC, but less than 20% mortality in shrimp maintained at 32oC. When some of the surviving shrimp were transferred from 32oC to 25oC, 100% mortality occurred. The authors speculated that WSSV replication slowed down at the higher temperature. Field experiments conducted in Ecuador indicated that simple greenhouse systems built over earthen ponds maintained water temperatures 3-4oC above water temperatures observed in adjacent open ponds (Sonnenholzner and Rodriguez, 2002). Shrimp grown in ponds covered with greenhouses yielded significantly higher growth and survival than those in open ponds. WSSV infections were not detected in the former, while the latter suffered mortalities due to WSSV.
One of the major impacts of viral diseases on shrimp pond management has been in the area of water exchange. Ten years ago, frequent water exchanges were considered essential in semiintensive and intensive shrimp farming so that metabolic wastes accumulating in pond water could be flushed out. It was common practice to replace as much as 100% of the water on a daily basis during the last few weeks of a crop cycle. The incoming water, rarely treated except by filtration, also brought in viruses and their carriers. As a result, water exchange is reduced, eliminated or managed better now. Complete elimination of water exchange has lowered the incidence of diseases in many farms worldwide. Use of intensive aeration and water circulation is necessary if water exchange is completely eliminated in semi-intensive and intensive culture systems. Complete elimination of water exchange is not possible in extremely hot climates that have little precipitation. Due to water evaporation, pond salinity in such climates increases beyond tolerable limits (> 40 ppt) if there is no water exchange. It is recommended that water used for exchange is adequately treated to reduce the risk of viral transmission.
Stocking density determines the level of management required to produce a successful crop. Systems stocked intensively (>25 animals/m2) require a high level of water quality and stress and animal health management. When such intensive management is not logistically or economically feasible, farmers tend to reduce stocking densities. Whether such reductions result in lower disease incidence is not known. On the other hand, efforts have succeeded in developing zero-exchange pond systems that are stocked at high densities (125-140 animals/m2) and are disease free (Browdy et al., 2001). Obviously, management of stressful situations rather than lowering stocking density per se is more important in shrimp health management.
The role of feeds in shrimp health management
Feed and feed management play a critical role in shrimp health management at a number of levels. First, feeds should be free of disease-causing organisms. The shrimp culture industry widely uses pelleted feeds. The high temperature (>90ºC) used in the pelleting process is likely to eliminate most viral particles. The risk of viral transmission through feeds is therefore considered minimal. Second, feeding should be managed in ponds such that wastage and consequent organic loading are minimized. Frequent feeding is recommended as this minimizes wastage and ensures continuous availability of fresh feed to the animals. Feed pellets lose their integrity in water if left for more than few hours and contribute to wastage and resulting water quality problems. Third, and the most significant contribution of feeds in shrimp health management, is through the delivery of nutrients and products (Table 1) that enhance and stimulate the host defense system.
Table 1. Health products of promising potential that can be used in shrimp feeds.

To enlarge the image, click here
NUTRIENTS THAT ENHANCE SHRIMP IMMUNE SYSTEM
Immune response imposes energy cost in animals. It results in changes in nutrient partitioning and directs more nutrients to the immune system (Humphrey et al., 2002). Therefore, nutrient requirements for optimum health status of an animal are expected to be higher than those for growth. Apart from this, certain nutrients are specifically implicated in the enhancement of immunity in many animals. These include some vitamins, trace minerals, ω-3 fatty acids, phospholipids, carotenoid pigments and nucleotides.
Merchie et al. (1998) found that increasing vitamin C level from 100 to 3400 mg/kg diet resulted in a concomitant drop in mortality of post-larval P. monondon subjected to osmotic shock. They also reported that high vitamin C or astaxanthin levels resulted in an increased resistance to salinity shock. Chien et al. (2002) reported that dietary supplementation with 80 mg/kg astaxanthin improved survival of P. monondon exposed to toxic levels of ammonia. The positive effect of vitamin C and astaxanthin on shrimp immunity is probably related to their antioxidant properties. In addition, vitamin C also plays a role in wound healing. Lavens and Sorgeloos (2000) noted that feeding post-larvae shrimp with Artemia nauplii (brine shrimp) that have been enriched with highly unsaturated fatty acids improved their ability to survive salinity shock. Coutteau et al. (2000) showed that phosphotidyl choline included in the diets of P. japonicus and P. vannamei at 1.5% significantly improved stress resistance in the animals. Both ω-3 and ω-6 fatty acids are precursors of eicosanoids that are potent mediators of inflammatory response in higher animals.
Phospholipids play a major role in cell membrane integrity, a significant factor in the first line of defense against antigens in all organisms. Experience with other animal species, including fish, shows that vitamin E, selenium and nucleotides may have a beneficial effect on the shrimp immune system. Vitamin E and selenium are highly effective antioxidants known to affect immune defense systems of vertebrates including fish. Vitamin E scavenges free radicals generated during the early stages of lipid peroxidation in cell membranes, while selenium is a component of glutathione peroxidase, which reduces the level of lipid peroxidation inside the cells. Given the facts that almost all highly unsaturated fatty acids are required for optimum growth of shrimp, and that these fatty acids are extremely prone to oxidation, dietary provision of vitamin E and selenium becomes all the more important.
Nucleotides are increasingly being perceived as semi-essential nutrients in human and animal nutrition. Although endogenous synthesis of nucleotides occurs in tissues, it is an energetically expensive process. Furthermore, cells involved in immune response do not synthesize nucleotides. Nucleotides from dietary sources are preferably used at times of rapid growth or physiological stress. Pointing out that natural food organisms and ingredients rich in nucleic acids such as bacteria, fish solubles, meat solubles and yeast extracts have growth promoting effects in shrimp, Devresse (2000) proposed that nucleotides are perhaps critical in shrimp health management. Health aspects of nucleotides have not been studied in shrimp. In trout and salmon, Burrels et al. (2001) have shown that nucleotide-supplementation caused greater resistance to microbial infections.
IMMUNOSTIMULANTS
Immunostimulants are compounds that stimulate the non-specific defense mechanisms in organisms. In shrimp, non-self recognition is the key to immune system stimulation, and this is mediated by pattern recognition proteins that identify and bind with specific molecules on the invading organisms.
Specific cell wall carbohydrates of bacteria and fungi have been identified as molecules that are identified by the pattern recognition proteins. These molecules are widely used as immunostimulants. Newman and Bullis (2001) provided an excellent review of studies that have evaluated immunostimulants in shrimp. The three most common immunostimulants that have been tested in shrimp are lipopolysaccharide (LPS), glucans and peptidoglycan (PG). LPS and PG are bacterial cell wall carbohydrates, while glucans are a set of polymeric glucose molecules found on fungal cell walls.
Lipopolysaccharide is a cell wall component of Gram-negative bacteria and consists of lipids and carbohydrates. Dead Vibrio cell suspensions are the most common form of LPS applications in experiments. About 20 studies have tested LPS in various species of shrimp. The results show that LPS exposure generally increases resistance to bacterial infections. Some tests show improved antiviral activity, better growth rates, increased resistance to stress and higher survival. The preparations are heat stable and can be administered via feed, or as immersion baths.
Peptidoglycan is a cell wall component of Grampositive bacteria. A couple of studies have found it to be effective in increasing shrimp resistance to viral infections. The most promising and conclusive study was that of Itami et al. (1998). The authors found that peptidoglycan derived from Bifidobacterium thermophilum fed at the rate of 0.2 mg/kg bodyweight/day offered protection against WSSV in P. japonicus.
Glucans are found on the cell walls of fungi. Glucans have been tested in shrimp in more than 10 studies. Glucans are also the most widely used immunostimulant in the shrimp farming industry. Nevertheless, the results are equivocal. While some studies show that they are effective in increasing shrimp resistance to pathogens, others show no efficacy. Some studies also show that there are adverse effects of glucan use on growth, survival and disease resistance. Scholz et al. (1999) reported that P. vannamei fed ß-glucan showed poorer survival and resistance to infection by a virulent strain of Vibrio harveyi.
Newman and Bullis (2001) pointed out that glucans derived from the mycelia of a mushroom, Schizophyllum commune, were much more effective in antibacterial activity than glucans derived from a baker’s yeast, Saccharomyces cerevisiae. Raa (2000) suggested that ß-1,3/1,6 glucan is the molecule that is recognized by the immune system and that its isolation from yeast must be done carefully in order to maintain its sidebranches intact, and thereby preserve its biological activity. Glucans are heat-labile and break down at temperatures above 130ºC. Crustaceans are suspected to have the ability to digest glucans, which may further explain the ineffectiveness of glucans as immunostimulants.
Mannan oligosaccharides, another component of yeast cell wall, have recently gained attention as immunostimulants in shrimp culture. In other animal species, mannan oligosaccharides have been established as molecules that bind with some bacterial species that have a specific receptor to the molecules. This binding prevents the bacterial species from colonizing the intestinal cell wall of animals. It is suggested that the molecules also adsorb potentially immunosuppressive mycotoxins. In the above two respects, mannan oligosaccharides are not strictly immunostimulants. However, evidence is mounting that the molecules may have a immunostimulatory role in other animal species (Cotter et al., 2002). Investigation of the effect of mannan oligosaccharides on shrimp health is required.
Fucoidan, a sulfated polysaccharide, has shown promise as a viral inhibitor in shrimp. Takahashi et al. (1998) reported that dietary administration of fucoidan derived from the brown alga, Cladosiphon okamuranus, reduced mortality due to WSSV. Purified fucoidan is too expensive to justify its use in shrimp feeds, however the use of crude extracts of brown algae is a possibility. In Japan, shrimp farmers already use a combination of peptidoglycan and an algal extract to protect shrimp against another form of WSSV.
Although immunostimulants are widely used in the shrimp culture industry at present, there are several unresolved issues concerning their field application. The primary concern is related to dosage: how much to apply and how long should application continue? Some believe that long-term and continuous exposure to immunostimulants exerts an energy cost on the animal and may even weaken or desensitize the immune system. Administration of immunostimulants during or after an infection may also be deleterious. It is widely accepted that immunostimulants exert their protective effect only on a short-term basis because they elicit only non-specific defense mechanisms. So, farmers need to understand how frequently the products need to be used. Finally, the interactive effects between two types of immunostimulants need to be understood as well. For example, beneficial effects of combining lipopolysaccharides and glucans have been noted by some (see Newman and Bullis, 2001).
PROBIOTICS
Several probiotic products are used in shrimp farming, particularly in Asia. Unlike in terrestrial animal farming, probiotic application in aquaculture extends beyond maintaining the intestinal microbial balance of host animals (Gatesoupe, 1999). Maintaining optimal microbial balance and eliminating pathogenic bacteria in the pond is one of the stated applications. Bioremediation of organic wastes produced by the animal in the pond water, and thereby enabling good water quality is another major application. In fact, a majority of probiotic products in aquaculture are intended for application in water, not through feed. It is assumed that the probiotic microbes added in water eventually enter the animal’s intestine. While probiotic bacteria added in the culture media has conferred disease resistance to shrimp (for example, Uma et al., 1999), it is not clear whether the benefit is due to intestinal colonization. On the other hand, dietary delivery of probiotic organisms has shown positive effect on shrimp health in a number of studies (Rengpipat et al., 1998; Uma et al., 1999; Scholz et al., 1999; Rengpipat et al., 2000).
Probiotic bacteria that have been shown effective in shrimp health include the following groups: Vibrio (particularly, V. alginolyticus) and various Bacillus and Lactobacillus species and strains. Rengpipat et al. (1998) showed that even the lyophilized form of bacteria (Bacillus) can be effective. The effect that has been demonstrated in almost all studies is the control of infection by the pathogenic bacteria V. harveyi. In addition, increase in phagocytosis, melanization activity and antibacterial activities has also been demonstrated (Rengpipat et al., 2000). The mechanisms suggested for the beneficial effects of probiotic bacteria in improving shrimp health are: (1) competitive exclusion of pathogenic bacteria in the gut; (2) cell wall components of probiotic bacteria stimulating the innate immune system of shrimp; and (3) enzymes and antibacterial compounds produced by the probiotic bacteria being detrimental to the pathogenic bacteria.
Scholz et al. (1999) showed that yeasts (S. cerevisiae and Phaffia rhodozyma) enhanced shrimp resistance to vibriosis. It is surprising that only limited attention has been paid to yeasts as probiotic organisms in shrimp. Their cell wall is a rich source of glucans and mannans that stimulate the immune system. They are rich in nucleotides, vitamins and trace minerals that are essential nutrients for optimum immune system function. Yeasts certainly deserve more attention in the context of shrimp health management. As noted above, probiotic organisms have been shown to be effective only for pathogenic bacterial control. Efforts are underway to identify bacteria with antiviral effects (Horowitz and Horowitz, 2001). Certain cyanobacteria and Pseudomonas strains with antiviral activity have been identified and are being tested.
Horowitz and Horowitz (2001) advocate caution in the application of probiotics. Continued exposure to probiotics may lower the sensitivity of pathogenic bacteria to the antibiotic compounds produced by the probiotic organism and may even lead to the development of resistant strains. Some of the antibiotic residues found in shrimp may actually originate from the probiotic organisms. Given the stringent regulations on antibiotic residues in food by major shrimp-consuming countries, this speculation is worth further investigation. There is also a risk that the otherwise benign probiotic organisms may become pathogenic due to genetic exchange between microbes. Alternatively, genes that provide the probiotic organism’s superior survival skills may transfer to the pathogenic bacteria making the latter even more resistant to destruction.
Conclusions
Shrimp diseases are the single most constraining factor on the profitability and progress of shrimp farming worldwide. Prevention of diseases is a major challenge. Shrimp have an innate immune system that responds to infections, but their adaptive immune system is inadequate to handle most infections. Particularly poor is the ability of shrimp to fight viral infections. Antibiotics do not work against most infections, and their use is banned in many countries anyway. As a result, emphasis in shrimp health management is on biosecurity – exclusion of infectious organisms and their carriers in the culture system. Environmental management to minimize stress also plays a major role in disease prevention. Developing a fool-proof biosecurity system and managing shrimp culture systems without any stress are not realistic goals. Improving the host defense capabilities should therefore be part of any practical shrimp health management program.
Enhancing the immune system through nutrients and immunostimulants is important in improving host defense capabilities in shrimp. Probiotics also play an effective role in improving defense capabilities. Immune-enhancing nutrients that have been identified in shrimp are vitamin C, ω-3 fatty acids, phosphotidyl choline and astaxanthin. Other nutrients with potential immune-enhancing properties are nucleotides, vitamin E and selenium. Compounds that stimulate immune response in shrimp are mainly derived from the cell walls of single cell organisms. Lipopolysaccharides and glucans have been found to be effective against bacterial infections, although the effect of glucans has not been consistent. Peptidoglycan and fucoidan have been found to be effective against viral infections, but studies on these molecules are limited. Furthermore, they may not be cost-effective. Mannan oligosaccharides that have been found to be effective in improving health status of other animal species need to be investigated in shrimp. Dietary delivery of some probiotic bacteria and yeasts increases resistance of shrimp to bacterial infections.
Although a much better understanding of shrimp diseases, the immune system and the effects of nutrients, immunostimulants and probiotics has been achieved in the past five years, there is much more to learn. At the fundamental level, there is a strong need to develop predictable indices of immune response and a standardized disease challenge protocol. It will make interpreting results of independent studies on effects of a given nutrient or product more reliable. At the application level, there is an array of needs ranging from empirical testing of products to optimizing application (doseresponse, application protocols, etc.). There is also a need to test nutrients, molecules and microbes that have not been investigated adequately in the past. Antioxidants, nucleotides, mannan oligosaccharides, yeasts and antiviral bacteria deserve the most attention in the future.
Author’s note on taxonomy
The genus Penaeus was revised a few years ago. However, this revision has not been universally accepted. This author has used the unrevised generic names throughout the article except in the list of references where they appear as reported in the original source.
References
Author: A. VICTOR SURESHArala-Chaves, M. and T. Sequeira. 2000. Is there any kind of adaptive immunity in invertebrates? Aquaculture 191:247-258.
Bachère, E. 2000. Shrimp immunity and disease control. Aquaculture 191:3-11.
Browdy, C.L., D. Bratvold, A.D. Stokes and R.P. McIntosh. 2001. Perspectives on the application of closed shrimp culture systems. In: The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture 2001 (C.L. Browdy and D.E. Jory, eds.). The World Aquaculture Society, Baton Rouge, LA, USA, pp. 20-34.
Burrels, C., P.D. Williams and P.F. Forno. 2001. Dietary nucleotides: a novel supplement in fish feeds. I. Effects on resistance to disease in salmonids. Aquaculture 199:159-169.
Chien, Y-H., C-H. Pan and B. Hunter. 2002. The resistance to ammonia stress by Penaeus monodon juvenile fed diets supplemented with astaxanthin. In: Book of Abstracts. World Aquaculture 2002. The World Aquaculture Society, Baton Rouge, LA, USA, p. 134.
Cotter, P.F., A.E. Sefton and M.S. Lilburn. 2002. Manipulating the immune system of layers and breeders: novel applications of mannan oligosaccharides. In: Nutritional Biotechnology in the Feed and Food Industries. Proceedings of Alltech’s Eighteenth Annual Symposium. (T.P. Lyons and K.A. Jacques, eds). Nottingham University Press, Nottingham, England, pp. 21- 27.
Coutteau, P., E.K.M. Kontara and P. Sorgeloos. 2000. Nutritional value of phosphatidyl choline purified from soybean and marine fish roe in the diet of postlarval Penaeus japonicus Bate and Penaeus vannamei Boone. Aquaculture 181:331- 345.
Devresse, B. 2000. Nucleotides – a key nutrient for the immune system of shrimp? Aqua Feed International 3(2):35-39.
Fegan, D.F. and H.C. Clifford. 2001. Health management for viral diseases in shrimp farms. In: The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture 2001 (C.L. Browdy and D.E. Jory, eds). The World Aquaculture Society, Baton Rouge, LA, USA, pp. 168-198.
Flegel, T.W. 1997. Major viral diseases of the black tiger prawn (Penaeus monodon) in Thailand. World J. Microb. and Biotech. 12:433-442.
Flegel, T.W. 2001. The shrimp response to viral pathogens. In: The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture 2001 (C.L. Browdy and D.E. Jory, eds). The World Aquaculture Society, Baton Rouge, LA, USA. pp. 254-278.
Gatesoupe, F.J. 1999. The use of probiotics in aquaculture. Aquaculture 180:147-165.
Horowitz, A. and S. Horowitz. 2001. Disease control in shrimp aquaculture from a microbial ecology perspective. In: The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture 2001. (C.L. Browdy and D.E. Jory, eds.). The World Aquaculture Society, Baton Rouge, LA, USA. pp. 199-218.
Humphrey, B.D., E.A. Koutsos and K.C. Klasing. 2002. Requirements and priorities of the immune system for nutrients. In: Nutritional Biotechnology in the Feed and Food Industries. Proceedings of Alltech’s 18th Annual Symposium (T.P. Lyons and K.A. Jacques, eds.). Nottingham University Press, Nottingham, UK, pp. 69-78.
Itami, T., M. Asano, K. Tokushige, K. Kubano, A. Nakawaga, N. Takeno, H. Nishimura, M. Maeda, M. Kondo and Y. Takahashi. 1998. Enhancement of disease resistance of Kuruma shrimp, Penaeus japonicus, after oral administration of peptidoglycan derived from Bifidobacterium thermophilum. Aquaculture 164:277-288.
Lavens, P. and P. Sorgeloos. 2000. Experiences on importance of diet for shrimp larval quality. Aquaculture 191:169-176.
Lightner, D.V. 1996. A handbook of pathology and diagnostic procedures for diseases of penaeid shrimp, The World Aquaculture Society, Baton Rouge, LA, USA.
Merchie, G., E. Kontara, P. Lavens, R. Robles, K. Kurmaly and P. Sorgeloos. 1998. Effect of vitamin C and astaxanthin on stress and disease resistance of postlarval tiger shrimp, Penaeus monodon (Fabricius). Aquaculture Research 29:579-585.
Newman, S.G. and R.A. Bullis. 2001. Immune mechanisms of shrimp: form, function and practical application. In: The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture 2001 (C.L. Browdy and D.E. Jory, eds.). The World Aqua. Soc., Baton Rouge, LA, USA, pp. 226- 237.
Raa, J. 2000. Beta-1,3/1,6-glucans: Modern science and ancient wisdom combined for the benefit of aquaculture. Aqua Feed International 3(2):32-34.
Rengpipat, S., W. Phianphak, S. Piyatiratitivorakul and P. Menasveta. 1998. Effects of a probiotic bacterium on black tiger shrimp Penaeus monodon survival and growth. Aquaculture 167:301-313.
Rengpipat, S., S. Rukpratanporn, S. Piyatiratitivorakul and P. Menasveta. 2000. Immunity enhancement in black tiger shrimp (Penaeus monodon) by a probiont bacterium (Bacillus S11). Aquaculture 191:271-288.
Scholz, U., G. Garcia Diaz, D. Ricque, L.E. Cruz Suarez, F. Vargas Albores and J. Latchford. 1999. Enhancement of vibriosis resistance in juvenile Penaeus vannamei by supplementation of diets with different yeast products. Aquaculture 176:271-283.
Sonnenholzner, S. and J. Rodriguez. 2002. Temperature and WSSV: CENAIM studies promising shrimp culture technique. Global Aqua. Advocate 5(4):55-57.
Takahashi, Y., K. Uehara, R. Watanabe, T. Okumura, T. Yamashita, H. Omura, T. Yomo, T. Kawano, A. Kanemitsu, H. Narasaka, N. Suzuki and T. Itami. 1998. Efficacy of oral administration of fucoidan, a sulfated polysaccharide, in controlling white spot syndrome in Kuruma shrimp in Japan. In: Advances in Shrimp Biotechnology (T.W. Flegel, ed). National Center for Genetic Engineering and Biotechnology, pp. 171-173.
Uma, A., T. Jawahar Abraham and V. Sundararaj. 1999. Effect of a probiotic bacterium, Lactobacillus plantarum on disease resistance of Penaeus indicus larvae. Indian J. Fisheries 46:367-373.
Vidal, O.M., C.B. Granja, F. Aranguren, J.A. Brock and M. Salazar. 2001. A profound effect of hypothermia on survival of Litopenaeus vannamei juveniles infected with white spot syndrome virus. J. World Aqua. Soc. 32:364-372.
Withyachumnarnkul, B. 1999. Results from black tiger shrimp (Penaeus monodon) culture ponds stocked with PL PCR positive or negative for white-spot syndrome virus (WSSV). Dis. Aquatic Organisms 39:21-27.
Aquaculture Feed Industry Consultant, Chennai, India

Can somebody help me? I need info about the using of PENERGETIC in shrimp, I need data info




.jpg&w=3840&q=75)