Development of a duplex Fluorescent Microsphere Immunoassay (FMIA) for the detection of antibody responses to influenza A and newcastle disease viruses
Published: March 12, 2015
By: Mathieu Pinettea, c, , Juan C. Rodriguez-lecompte a, b, , John Pasick c, , Davor Ojkic d, , Marsha Leith c, , Matthew Suderman c, , Yohannes Berhane a, c, (a Department of Animal Science, University of Manitoba, Winnipeg, Manitoba R3T 2 N2, Canada; b Atlantic Veterinary College, University of Prince Edward Island, Charlottetown, Prince Edward Island C1A 4P3, Canada; c National Centre for Foreign Animal Diseases, Winnipeg, Manitoba, Canada; d Animal Health Laboratory, University of Guelph, Guelph, Ontario, C
Journal of Immunological Methods 405 (2014) 167–177. http://dx.doi.org/10.1016/j.jim.2014.02.004 0022-1759 Crown Copyright © 2014 Published by Elsevier B.V. Contents lists available at ScienceDirect Journal of Immunological Methods journal homepage: www.elsevier.com/locate/jim Open access under CC BY-NC-ND license. Open access under CC BY-NC-ND license
Abstract
Highly pathogenic avian influenza virus (HPAI) and virulent forms of avian paramyxovirus-1 (APMV-1) cause serious illnesses in domestic poultry, both of which are reportable to the World Organization of Animal Health (OIE). The clinical presentation of avian influenza (AI) and APMV-1 infections are difficult to differentiate, emphasizing the importance of rapid and sensitive serologic assays that are able to distinguish them. Currently, a variety of serological assays are used for the serologic diagnosis of both diseases, but these assays are not used in multiplex formats. In this study, development of a duplex fluorescent microsphere immunoassay (FMIA) based on Luminex xMAP Technology is described. The assay employs MagPlex magnetic microspheres that are covalently coated with recombinant avian influenza virus nucleoprotein and APMV-1 nucleocapsid antigens produced in a baculovirus insect cell expression system. The assay is able to detect AIV antibodies against all existing hemagglutinin (H1–H16) subtypes and simultaneously detect antibodies against APMV-1. In the process of this assay development different bead coupling conditions were compared. The assay has the capability of detecting serum antibodies from chickens and turkeys and optimization was accomplished by using 2462 chicken and 446 turkey field and experimental sera and had a comparable detection capability with currently used assays in the laboratory. Assay threshold values were calculated with Receiver Operating Characteristic Analysis (ROC) in non-parametric analysis due to a highly skewed data distribution; this analysis resulted in AIV nucleoprotein relative diagnostic sensitivity and specificity of 99.7%, and 97.3% respectively. The APMV-1 nucleocapsid relative diagnostic sensitivity and specificity were 95.4%, and 98.5% respectively.
1. Introduction
Recent outbreaks of avian influenza (AI) and avian paramyxovirus-1 (APMV-1) infections in domestic poultry in Canada, Mexico, China, India, Vietnam, Colombia, Russia, Italy, Japan, Korea, parts of the Middle East, and parts of Africa, have led to the destruction of a large number of flocks with a total economic impact in the millions of dollars (Mutinelli et al., 2003; Pasick et al., 2012; Pawar et al., 2012; Wang et al., 2008; Watson et al., 2009). From 2006 to 2009, the OIE with its 176 member countries, reported that APMV-1 caused outbreaks in 56 countries while HPAI caused outbreaks in 28 countries (World Livestock Disease Atlas, 2011). The highly contagious nature and similar clinical characteristics of both APMV-1 and AI infections emphasize the importance of being able to perform rapid and accurate serologic diagnosis that can determine which disease is causing the outbreak (Alexander and Senne, 2003; Leijon et al., 2011).
Avian influenza viruses belong to the influenza A genus of the Orthomyxoviridae family and have a genome that is made up of 8 single-stranded RNA segments of negative sense that code for 11 proteins. They are classified into serological subtypes based on their viral surface glycoproteins–hemagglutinin (H1 to H16) and neuraminidase (N1 to N9). Based on their potential to cause disease in chickens, AIV are divided into low pathogenic (LPAI) and highly pathogenic (HPAI) viruses. Low pathogenic AIV causes a localized mild respiratory or enteric disease, while few viruses belonging to H5 and H7 subtypes are considered highly pathogenic AIV and cause systemic disease of domestic poultry with up to 100% mortality. Low pathogenic AI and HPAI H5 and H7 are both reportable to the OIE.
Newcastle disease virus (NDV) also known as APMV-1 belongs to the family of Paramyxoviridae, genus Avulavirus. The virus has a single stranded RNA genome of negative sense containing 6 genes that code for 9 structural and non-structural proteins. The OIE defines velogenic ND as a virus with an intracerebral pathogenicity index of N0.7 for day-old chicks, or with a polybasic cleavage site in the F protein. Newcastle disease can also be caused by lentogenic strains that cause sub-clinical to mild clinical disease; mesogenic – causing non-fatal respiratory disease; and velogenic – most pathogenic and often fatal (Aldous and Alexander, 2001; Kapczynski et al., 2013; Seal et al., 2002).
Currently, a variety of serological assays are being used for the detection of AIV and APMV-1 antibodies in domestic poultry to monitor the antibody response due to infection, or for the purpose of monitoring vaccine efficacy. Although different commercially available ELISA kits are used for detecting APMV-1 antibodies in domestic poultry, the HI assay continues to be the “gold standard” for detecting APMV-1 antibodies. For the detection of AIV antibodies, the agar gel immunodiffusion (AGID) and hemagglutination inhibition (HI) test remain as the “gold standard” methods approved by the OIE. The AGID test detects antibodies against the highly conserved nucleoprotein (NP) and matrix (M1) proteins. However, the assay is less sensitive and is prone to producing higher rates of false negatives (Snyder et al., 1985; Watson et al., 2009; Zhou et al., 1998). Unlike the AGID test, the HI assay specifically detects hemagglutinin subtype specific antibodies using H1–H16 antigens, however it has several drawbacks: it is labor intensive, difficult to automate as results are read visually, it requires subtype specific antigens that are difficult to produce under ordinary lab conditions, the antigens are hard to standardize as they need to be calibrated for each assay, and the assay depends on fresh red blood cells (RBC) from chickens that can be difficult to acquire (Comin et al., 2013; Upadhyay et al., 2009; Zhou et al., 1998). In addition, the reference antigen used in the HI assay needs to be updated on a yearly basis with respect to the currently circulating viruses, otherwise this may impair the sensitivity of the assay dramatically.
Currently, different laboratories use the HI assay in combination with an ELISA to check the immune status of vaccinated flocks (Desvaux et al., 2012; Toffan et al., 2010). Despite their robustness and higher sensitivity, ELISAs don't have multiplexing capabilities, and for each individual analyte to be detected a separate test must be carried out making the use of ELISAs time consuming and laborious. In the current study, the development of a duplex florescent microsphere immunoassay (FMIA) based on the xMAP technology for simultaneous detection and differentiation of antibody response to AIV and APMV-1 infections is described. The duplex FMIA is based on AIV nucleoprotein (AIV-NP) and APMV-1 nucleocapsid protein (APMV-1-NC) that are the most abundant proteins in AIV and APMV-1 respectively. Both AIV-NP and APMV-1-NC proteins are highly conserved and immunogenic, which therefore makes them suitable candidates for the detection of virus-specific antibodies and for serologic diagnosis of each individual disease.
2. Materials and methods
2.1. Cloning of APMV-1 nucleocapsid and AIV nucleoprotein genes
Avian Influenza nucleoprotein from A/Duck/AB/AFLB-C16/ 2008 (H7N7) and APMV-1-NC from APMV-1 vaccine (La'Sota) were selected for the development of the assay. The AIV-NP gene was amplified using universal AIV-NP primers as described previously (Hoffmann et al., 2001) and cloned into PCR4 TOPO cloning vector (Invitrogen, Ontario, CA). The coding region of AIV-NP was re-amplified using forward primers: ACG CGC CCG CTA TGG CGT CTC AAG GCG CCA A and reverse primers: CGG TCT GAG TTA ATT GTC ATA CTC CTC TGC ATT. The full nucleocapsid gene from APMV-1, La'Sota strain was amplified using primers: ATG TCT TCC GTA TTT GAT GA (forward) and TCA ATA CCC CCA GTC GGT GT (reverse). The amplified AIV-NP and APMV-1-NC PCR products were sub-cloned in-frame into the Not 1 and Xba 1 restriction enzyme sites of pAB-bee-FH transfer vector which included an 8X HIS tag for later protein purification (AB Vector, California). Correct frame and sequence of the gene in the transfer vector (pAB-bee-FH–APMV-1-NC or AIV-NP) were confirmed by sequencing.
2.2. Transfection and generation of recombinant baculovirus
The APMV-1-NC or AIV-NP genes that were cloned in the transfer vector were co-transfected into Spodofera frugiperda (Sf-9) cells with ProFold-ER1 linearized baculovirus DNA (AB Vector, California, USA). All procedures for the production of recombinant proteins, including plaque purification, production of high titer stocks and optimization of protein expression were performed according to the manufacturer's protocols (AB Vector, California, USA). The APMV-1-NC or AIV-NP recombinant proteins from the cell pellets and media supernatants were purified by batch procedure using Ni-NTA agarose (Qiagen, Maryland, USA) according to manufacturer's instructions.
2.3. Coupling of proteins to Luminex MagPlex Microspheres
Coupling of proteins to Luminex MagPlex Microspheres was carried out according to the manufacturer's instructions (Luminex Corporation, Texas, USA). Prior to coupling, the recombinant proteins were desalted using micro-biospin 6 columns (Bio-Rad, California, USA) according to the manufacturer's instructions to exchange the buffer from imidazole to PBS, and quantified using Pierce BCA Protein Quantification Kit (Thermo Scientific, Maryland, USA). Recombinant APMV-1-NC protein were coupled to MagPlex region #012 microspheres and AIV-NP were coupled to region #015 (Luminex Corporation, Texas, USA). Twelve micrograms (μg) of each recombinant protein were coupled to a fixed amount of beads (1.25 × 106 ) per 1× reaction.
2.4. Duplex FMIA
Coupled APMV-1-NC and AIV-NP beads at concentration of 2000 beads/well were combined and blocked for 2 h in 5% (v/v) normal donkey serum (Jackson ImmunoResearch Laboratories Inc., Pennsylvania, USA) in PBS. Twenty five microliters of bead suspension and 25 μl of test serum diluted 1:500 in 1% (v/v) normal donkey serum in PBS were added to each well. The plate was left at room temperature in the dark on a shaking incubator for 1 h. A wash program consisting of 3 washes with 200 μl of PBS-tween 20 in magnetic plate washer was completed (Bio-Tek, 405 Select TS), and the plates were incubated with 50 μl of 2 μg/ml biotinylated donkey anti-chicken immunoglobulin (Jackson ImmunoResearch Laboratories, Inc., Pennsylvania, USA) for 30 min at room temperature. Plates were washed again before the final addition of 75 μl of 10 mg/ml streptavidin-phycoerythrin conjugate (Jackson ImmunoResearch Laboratories Inc., Pennsylvania, USA) and incubation at room temperature for 15 min. Following the final wash, each well of beads were re-suspended in 125 μl of wash buffer and analyzed with a BioPlex 100 instrument running Bioplex Manager 6.1 software (Bio-Rad, California, USA). The results were expressed as arbitrary units of median fluorescence intensity (MFI).
2.4.1. Luminex reference curve
A 12-step reference curve was developed for use with the assay as a method of variability monitoring between consecutive runs by comparing the changes in fluorescent signal of the reference curve, and using this data to calculate a correction factor which was applied to all data sets. The references were prepared by mixing AIV and APMV-1 hyper-immune sera of known titers and diluted 2-fold in negative chicken serum to produce a 12-step curve. These reference samples were tested for the presence of AIV and APMV-1 antibodies using the IDEXX ELISA kits and hemagglutination inhibition assays. The hemagglutination inhibition titers of both the APMV-1 and AI sera that were used for making the standards were 512 and 128 consecutively.
2.4.2. Luminex internal controls
Region 18 and 19 MagPlex beads were used for internal controls for the assay. Highly purified chicken IgY (Jackson ImmunoResearch Laboratories, Inc., Pennsylvania, USA) and donkey anti chicken IgY (Jackson ImmunoResearch Laboratories, Inc., Pennsylvania, USA) 0.1 μg/ml concentrations each were coupled to these respective bead regions as described above, and included in the assay for the purpose of verifying the addition of secondary detection antibody and test serum respectively.
2.5. Hemagglutination inhibition assay
Hemagglutination inhibition (HI) assay was performed according to the methods described in the OIE manual (OIE Manual, 2008). For testing the serial bleeds from experimentally inoculated chickens, 4 HA units of homologous virus was combined with 2-fold serial dilutions of each serum sample. Chicken red blood cells (0.5% v/v suspension) were then added and HI endpoints read. Serum HI titre N1:4 was considered positive.
2.6. IDEXX ELISA
Commercial IDEXX AIV and APMV-1 ELISA kits were used to determine presence of AIV or APMV-1 antibodies in poultry sera. Depending on the source of the serum, the indirect APMV-1 antibody test kits for turkey and chicken sera were used (IDEXX Laboratories, Maine, USA). The competitive MultiS-Screen ELISA Kit was used for AIV antibody detection in turkey and chicken sera. Both assays were performed according to the manufacturer's protocols. Results were quantified by reading plates at 650 nm with SpectraMax Plus microplate reader (Bio-Tek Instruments Inc., Vermont, USA) using SoftMax Software (Bio-Tek Instruments, Vermont, USA).
2.7. Competitive Enzyme-linked Immunosorbent Assay (cELISA)
Competitive ELISA (cELISA) was performed as described by Yang et al., 2008. Microtiter plates coated with 1 μg of recombinant baculovirus AIV-NP protein in carbonate buffer (pH 9.6) at 100 μl/well and plates were incubated 4 °C overnight. After washing, equal volumes (50 μl) of diluted test sera (1:5) and hybridoma culture supernatants containing AIV-NP monoclonal antibody (1:200) were added to the plates and incubated at 37 °C for 1 h with agitation. Then, HRP conjugated anti-mouse IgG (Jackson ImmunoResearch Laboratories Inc., Pennsylvania, USA) was added and incubated for 1 h at 37 °C with subsequent washing. Lastly, the enzyme substrate, 3,3',5,5'-Tetramethylbenzidine (TMB, Sigma-Aldrich, Missouri, USA), was added and color development was stopped after 15 min with 50 μl/well of 2.0 M sulfuric acid. The OD was determined at 450 nm on an automated plate reader (SpectraMax Plus, Molecular Devices, California, USA).
2.8. Serum samples
2462 serum samples that were collected from SPF chickens, broilers chickens, layers and breeder chickens and an additional 446 turkey serum samples that were collected from turkey poults and breeders were used in the validation of the assay. All sera were pre-screened by a combination of assays including HI, in-house developed cELISA, and IDEXX ELISA to determine the presence or absence of detectable antibodies.
2.9. Reference AIV and APMV-1 antisera
Reference antisera against all existing 16 hemagglutinin subtypes of AI viruses and avian paramyxovirus (APMV) types 1–9 (Table 1) were produced by inoculating six-week-old white leghorn specific pathogen free (SPF) chickens that were obtained from the animal care unit of the Ottawa Laboratory Fallowfield (OLF), Canadian Food Inspection Agency (CFIA). The OLF SPF stock was developed from Charles River/SPAFAS birds (Storrs, Connecticut, USA). Negative SPF chicken sera used in this study were also collected from the same flock.
2.10. Field serum samples
Field serum samples used in this study were collected from broilers, turkey and chicken breeder flocks; some of which were vaccinated for APMV-1 using a combination of live and inactivated APMV-1 vaccines. Some serum samples were collected from turkeys that were vaccinated with inactivated H3N4 AIV vaccine (Berhane et al., 2012). Chicken field serum samples used in this study were donated to us by the Animal Health Laboratory (AHL), Guelph, Ontario or from archived serum database of NCFAD.
2.11. Production of experimental serum samples
During the production of experimental serum samples, all animal care and handling procedures were done according to the Canadian Council on Animal Care guidelines and as outlined in protocols approved by the institute.
Table 1 Reference strains of avian influenza (H1 to H16 subtypes) and avian paramyxovirus serotype 1–9 (excluding serotype 5) were used for immunizing specific pathogen free chickens for the production of reference antisera that was used for the evaluation of the avian influenza and avian paramyxovirus type 1 duplex FMIA.
2.11.1. Chicken serum
For the production of anti-APMV-1 serum samples, one group of chickens (n = 20) were inoculated intranasally and ocularly with 1 ml of inoculum containing 104 egg infective dose 50 [EID50] of La'Sota APMV-1 vaccine strain. For the production of AIV serum samples, a second group of chickens (n = 20) were also inoculated intranasally and ocularly with 1 ml of inoculum containing 104 plaque forming units (PFU) of low pathogenic influenza virus A/Anhuii/1/2013 (H7N9). Serum samples were collected at 0, 4, 7, 14, 21 and 28 days after infection (dpi). The H7N9 group of chickens was kept in enhanced level 3 animal cubicles
2.11.2. Turkey serum
For experimental turkey serum production, 8 to 10 week old influenza free turkey poults were bought from a commercial supplier in Manitoba. The turkeys were bled and pre-tested for the presence of influenza A group specific antibodies (NP) using cELISA and confirmed negative. The first group of turkeys (n = 10) were inoculated intranasally and ocularly with A/turkey/ON/1963 (H6N8), and the second group (n = 10) with A/turkey/MN/12877/1285/81 (H9N2). The turkeys were inoculated with 106 egg infectious does50 (EID50)/ml of virus inoculated intranasally and ocularly. Serum samples were collected at 0, 7, 14, 21 and 28 days after infection (dpi).
2.12. Data analysis
All statistics were done using the Microsoft Excel add on “XLSTAT-Life”. Basic frequency versus signal histograms were generated to visualize the distribution of the data. Receiver Operating Characteristic analysis (ROC) was used to determine accurate threshold values for the assay
3. Results
3.1. Recombinant protein production and purification
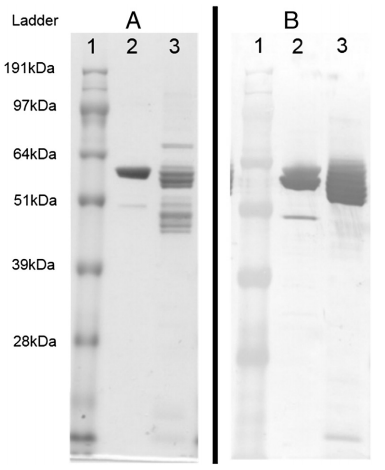
The AIV-NP and APMV-1-NC cloned into pAB-bee-FH transfer vectors were co-transfected with linearized ProFold™-ER1 baculovirus genomic vector into SF9 cells. The presence of Aequorea victoria green-fluorescent protein (GFP) in ProFold™-ER1 allowed for the convenient monitoring of recombinant baculovirus infection. Expression of both recombinant proteins (AIV-NP and APMV-1-NC) in the cell culture supernatants and cell pellets were monitored by running the denatured protein product in 10% Bis-Tris precast gels (Invitrogen) followed by Coomassie blue staining and Western blot analysis. The APMV-1-NC or AIV-NP recombinant proteins from the cell pellets and media supernatants were purified using Ni-NTA agarose and the purity was confirmed by SDS-PAGE followed by Coomassie Blue Staining (Fig. 1A) and the authenticity of the purified recombinant proteins was confirmed Western blot analysis (Fig. 1B).
3.2. Bead coupling dose response and protocol optimization
The quantity of protein used in each microsphere coupling reaction was based on recommendations within the Luminex literature to range from 5 μg to 12 μg of purified protein per 1X coupling reaction (1.25 × 106 beads per 100 μl reactions). Five confirmed negative serum and 6 confirmed positive sera were used to test the different quantities of protein bound to the beads. Doses of 6 μg, 12 μg, 23 μg, and 45 μg of each recombinant protein were tested in each bead coupling reaction. For both AIV-NP and APMV-1-NC proteins, coupling with 12 μg of protein per reaction gave the maximum signal-to-noise ratio (Fig. 2). The lowest background to noise ratio was achieved when the beads were blocked for 2 h in 5% (v/v) normal donkey serum in PBS and the test sera diluted at 1:500 in PBS solution containing 1% (v/v) normal donkey serum.
Fig. 1. Analysis of recombinant avian influenza nucleoprotein (2A and 2B) and avian paramyxovirus-1 nucleocapsid protein (3A and 3B) that were expressed using the baculovirus expression system were successfully purified and underwent sodium dodecyl sulfate–polyacrylamide gel electrophoresis followed by Coomassie Blue staining (Fig. 1A) and western immunoblot (Fig. 1B) using anti his antibody. Equivalent volumes of each protein (5 μl) were used for both Coomassie and western immunoblot analysis.
3.3. Assay development
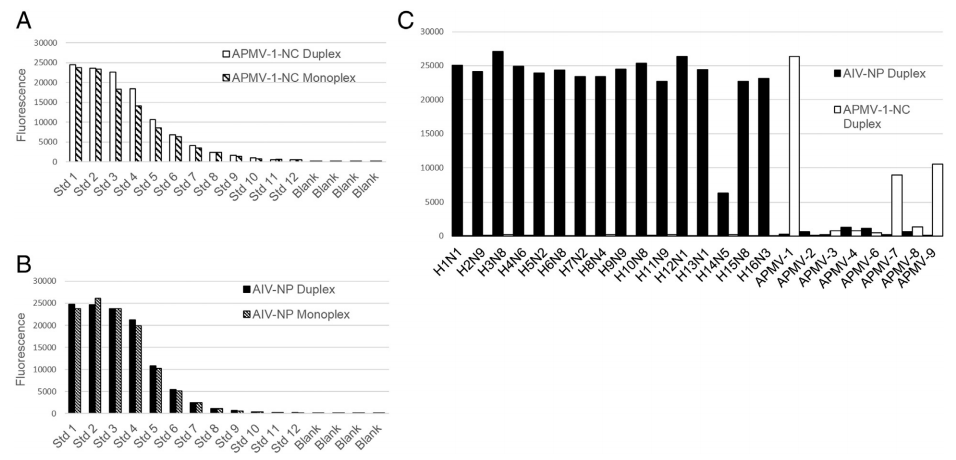
The reference curve developed for monitoring assay variability between consecutive runs was also used to demonstrate that the assay was not affected when detecting antibody with both beads simultaneously. Monoplex and duplex FMIA results for both AIV-NP and APMV-1-NC were analyzed using the non-parametric Wilcoxon, Mann–Whitney comparison test with 5% significance level (XLSTAT 2013). Neither result suggested to reject the null hypothesis (H0: The difference between the samples is equal to 0), so the monoplex and duplex FMIA results were not found to be significantly different (AIV-NP p-value 0.865; APMV-1-NC p-value 0.706). The AIV-NP coated beads were able to detect antibodies developed against all existing hemagglutinin subtypes (H1 to H16) while showing no cross reactivity with antisera developed against APMV 1, 2, 3, 4, 6, 7, 8, and 9 serotypes (Fig. 3).
The APMV-1-NC beads did not cross react with any of the H1 to H16 subtypes of influenza, and were able to detect antibodies to APMV-1 very strongly. The APMV-1-NC cross-reacted weakly with anti-sera developed to APMV-7 and APMV-9 (Fig. 3). The antibody coupled microspheres (chicken IgY and donkey anti-chicken IgY) were used as internal assay controls to confirm the addition of critical reagents. The presence of donkey anti-chicken IgY coupled microspheres ensured that test serum has been added. The presence of chicken IgY coupled microspheres, ensured that the secondary detection antibody has been added. The presence of these internal controls helped elucidate false negatives caused by user error. MFI values greater than 8000 for each internal control microsphere indicate successful reagent addition.
Fig. 2. The optimization of optimal amount of recombinant protein required for coupling to the Luminex MagPlex Microspheres (Luminex Corporation, Austin, Texas, USA) was assessed by testing 4 different concentrations of recombinant avian influenza nucleoprotein at 5.6 μg (A), 11.2 μg (B), 22.4 μg (C) and 44.8 μg (D) with 5 known negative and 6 known positive chicken sera. Beads that were coupled with 11.2 μg of recombinant avian influenza nucleoprotein showed the most favorable sample/negative ratio and also displayed the lowest overall background signal.
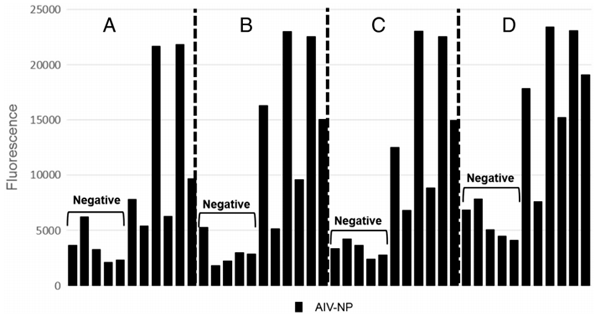
Fig. 3. Fig. 3A & B. Comparison of results of the FMIA using recombinant avian influenza nucleoprotein and avian paramyxovirus type 1 nucleocapsid coated beads when tested in monoplex and duplex formats with the Luminex reference sera. Fig. 3A describes avian paramyxovirus type 1 FMIA in monoplex and duplex formats and Fig. 3B — avian influenza monoplex and duplex FMIA. There was no significant difference in fluorescent mean values (MFI) when the assays were run in duplex or monoplex formats. C. The duplex fluorescent microsphere immunoassay for serologic diagnosis of avian influenza and avian paramyxovirus type 1 infection was tested using a panel of reference antisera against all the existing 16 hemagglutinin subtypes (H1 to H16) and avian paramyxoviruses type 1–9 (excluding type-5). The Luminex beads coated with avian influenza nucleoprotein were able to detect antibodies against all 16 hemagglutinin subtypes and beads coated with avian paramyxovirus-1 nucleocapsid protein were able to detect reference antisera against type 1 with minimal cross reactivity to type 7 and 9.
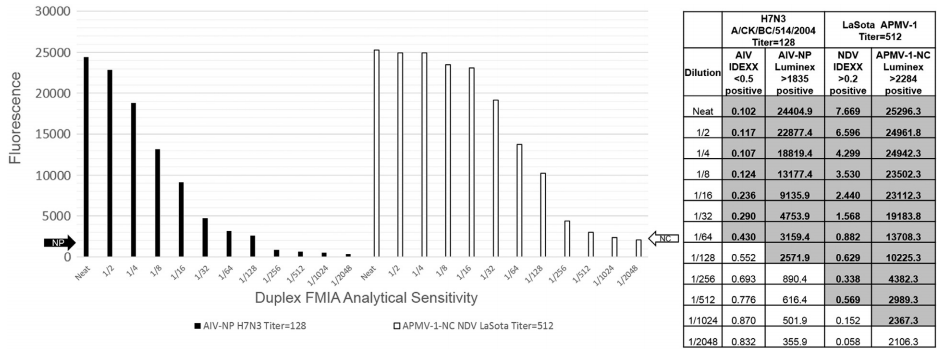
3.4. Duplex FMIA analytical sensitivity
In order to determine the analytical sensitivity of the assay, AIV and APMV-1 reference antisera that were developed in chickens were diluted 2-fold in negative SPF chickens serum (Sigma Aldrich, USA) in a 12-step dilution series. The APMV-1 reference antiserum was developed in chickens that were inoculated with La'Sota NDV vaccine strain. The hemagglutination inhibition titer of this antiserum to homologous antigen was 512. The AIV reference antiserum was produced in chickens that were inoculated with A/Chicken/ BC/514/2004 (H7N3) and HI titer of this antiserum to homologous antigen was 256. These 12 serial dilutions OF AIV and APMV-1 antisera were tested using the AIV-NP and APMV-1-NC duplex FMIA. The AIV-NP beads were able to detect antibodies in serum at dilutions as low as 1/128, and APMV-1-NC beads could detect antibodies at dilutions as low as 1/1024 before falling below the assay background (Fig. 4).
Fig. 4. A 12 step 2-fold serial dilutions of reference AI and APMV-1 sera were conducted to determine the analytical sensitivity of the duplex FMIA. The AIV-NP beads were able to detect dilution of antisera at 1/128, and APMV-1-NC beads were able to detect down to 1/1024.
3.5. Statistical normalization of results
The standard curve results were compiled for every run of the FMIA, providing a summary of how the assay performed over time. Statistical ANOVA testing was carried out across the individual reference controls and revealed a minor difference between runs. This prompted the normalization of the overall data to correct for assay variation. Averages of the MFI values of each standard were calculated across all runs and the calculated averages of the standards were used to generate an overall correction factor to determine the correct assay values. When values of the standards were higher or lower than the average, the entire standard curve usually reflected this shift either wholly above, or wholly below the average. For example, if standard 1 read a greater value than normal, the rest of the reference samples in that run also read greater values than normal. This allowed us to create a separate correction factor for each run of the assay and apply it uniformly to either raise or lower the observed values to normalize the values of the standard curve. The corrected assay values were then used for statistical analysis to determine the assay cut-off
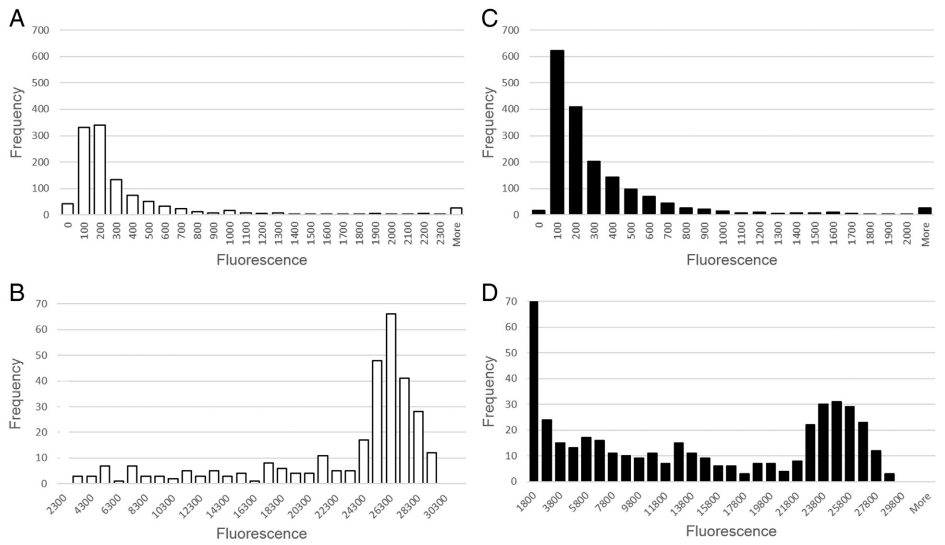
3.6. Receiver operating characteristic analysis for threshold determination
Receiver operating characteristic (ROC) analysis was used for assay threshold determination because of the inability to transform the data to fit a normal distribution even after attempted log10 and fourth root transformations (Fig. 5). The ROC analysis was used to calculate the threshold value determined by the ratio of true positives to false negatives. The median fluorescence intensity (MFI) at which the ratio of false positives and false negatives are lowest determines the optimal threshold. The IDEXX ELISA results were used as a guide for “true positive” and “true negative” status of the test serum undergoing analysis as the assay has been commercially validated. The diagnostic specificity of the APMV-1 IDEXX ELISA was 100% (n = 40 SPF sera), and the assay showed sensitivity of 91.5% in relation to HI results (n = 47) (IDEXX APMV-1 ELISA Validation Report 2010). The AIV MultiS-Screen ELISA has a diagnostic specificity of 99.7% (n = 5007), and diagnostic sensitivity that matched the HI (IDEXX AIV ELISA Validation Report). In the events where the IDEXX ELISA and the HI results were in disagreement, “true positivity or negativity” was assigned according to the HI as it is the current gold standard and also the assay that the IDEXX was validated against.
The AIV antibody response detection threshold of the duplex FMIA was calculated to be 1835 median fluorescent units with an associated diagnostic sensitivity of 99.7% and diagnostic specificity of 97.3% relative to the IDEXX ELISA. This threshold value yielded 332 relative true positives, 56 false positives, 1 false negative, and 1987 true negatives.
The avian paramyxovirus antibody response detection threshold of the duplex FMIA was calculated to be 2284 median fluorescent units (MFU) with an associated diagnostic sensitivity of 95.4% and diagnostic specificity of 98.5% relative to the IDEXX ELISA. This threshold yielded 313 true positives, 18 false positives, 15 false negatives, and 1149 true negatives.
All values were statistically significant within a 95% confidence interval (XLSTAT 2013).
Fig. 5. Figures describe the combined frequency distribution of all chicken and turkey serum data for threshold determination, separately analyzing positive and negative samples. A) Avian paramyxovirus 1 negative serum samples; B) Avian paramyxovirus type 1 positive serum samples; C) Avian influenza negative serum samples; D) Avian influenza positive serum samples.
3.7. Monitoring the kinetics of antibody response using the duplex FMIA
3.7.1. Chickens
The duplex FMIA was used to monitor the kinetics of antibody responses in 2 groups of chickens that were experimentally infected with influenza and APMV-1 La'Sota strain. Serum samples that were collected at 0, 4, 7, 14, 21 and 28 days after infection (dpi) were tested using the duplex FMIA. In addition, the serum samples were also tested for the presence of APMV-1 antibodies using the indirect IDEXX APMV-1 ELISA kit and HI assay. In addition to the FMIA, for detecting AIV antibodies the IDEXX AIV Multiscreen ELISA Kit was also used. All pre-bleed samples from chickens of both groups tested negative for AIV and APMV-1 antibodies using all test methods.
Chickens that were infected with APMV-1 started to develop antibody responses at 7 dpi and reached peak MFU by 14 dpi. The results were comparable to APMV-1 IDEXX ELISA results, but the duplex FMIA was more sensitive than HI assay. Results of this study are summarized in Fig. 6A.
Serum samples collected from the 2nd group of chickens that were infected with A/Anhuii/1/2013 (H7N9) were tested on the duplex FMIA. Infected chickens started to develop antibody responses starting at 4 dpi based on the IDEXX ELISA and duplex FMIA results. By 7 dpi, all samples tested positive on the IDEXX ELISA and duplex FMIA. However, there were only 3 samples that were positive on the HI assay. At 14 dpi, all samples tested positive on the IDEXX ELISA and on the duplex FMIA, however only 50% of samples had anti H7 hemagglutinin antibodies. By 21 dpi, all assays were detecting a positive immune response from all test serum. Results of this study are summarized in Fig. 6B.
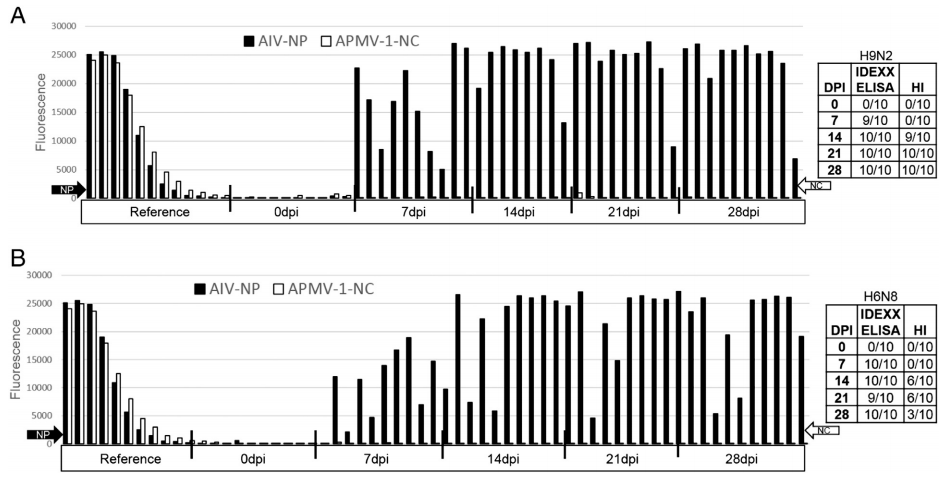
3.7.2. Turkeys
The duplex FMIA assay was also used to detect the kinetics of antibody responses in turkey poults that were infected with LPAI viruses. For this purpose, turkeys in the 1st group were inoculated with A/turkey/ON/1963 (H6N8), and the 2nd group with A/turkey/MN/12877/1285/81 (H9N2). Pre-bleed serum samples collected from these turkeys tested negative for APMV-1 and AIV antibodies using the FMIA or IDEXX ELISA tests for both antibodies. Based on the IDEXX ELISA and FMIA results, almost 100% of the turkeys in both groups elicited humoral immune response against AIV starting at 7 dpi and reached peak responses by 14 dpi. According to the HI assay results, anti-hemagglutinin antibodies were detected in the 90% of turkeys in the H9N2 group by 14 dpi and all turkeys seroconverted by 21 dpi and remained at higher level until the end of the study at 28 dpi. Sixty percent of turkeys that were inoculated with the H6N8 virus developed anti hemagglutinin antibodies at 14 and 21 dpi. Results of this study are summarized in Fig. 7.
Fig. 6. 6A). Kinetics of antibody response in chickens that were immunized with NDV La'Sota vaccine strain using the duplex FMIA, NDV IDEXX ELISA kit (IDEXX Laboratories, Maine, USA) and hemagglutination inhibition assay using homologous La'Sota antigen. According the FMIA data, 100% of chickens were able to sero-convert by 7 dpi according to the duplex assay and IDEXX ELISA. 6B) Kinetics of antibody response in chickens that were infected with LPAI A/Anhuii/1/2013 (H7N9) using the duplex FMIA, avian influenza Multiscreen ELISA Kit (IDEXX Laboratories, Maine, USA) and hemagglutination inhibition assay with homologous antigen. 100% of chickens were able to sero-convert by 7 dpi according to the duplex assay and the IDEXX ELISA.
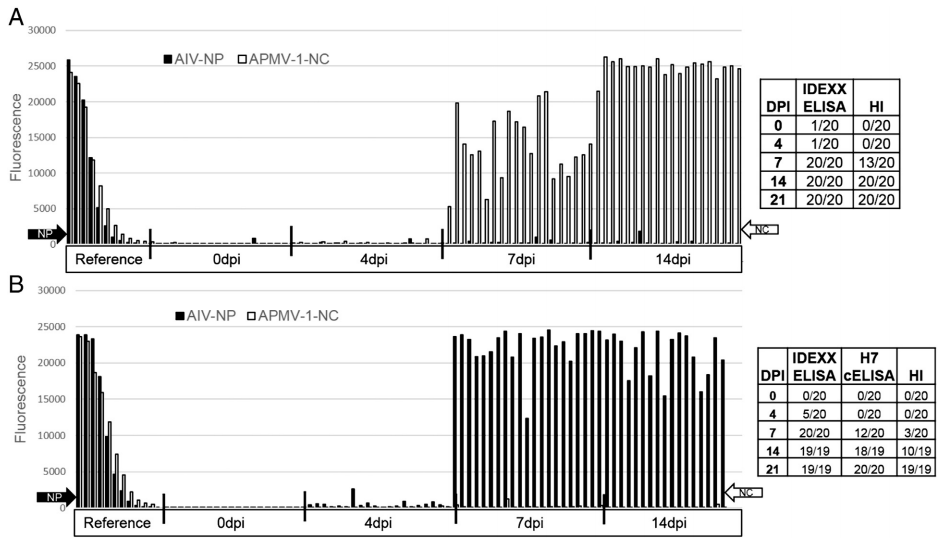
Fig. 7. Kinetics of antibody response in turkeys that were infected with either LPAI A/turkey/MN/12877/1285/81 (H9N2) (7A) or A/turkey/ON/1963 (H6N8) (7B) using the duplex FMIA, avian influenza Multiscreen ELISA Kit (IDEXX Laboratories, Maine, USA) and hemagglutination inhibition assay using corresponding homologous antigens. Almost 100% of turkeys were able to sero-convert by 7 dpi according to the duplex assay and the IDEXX ELISA.
4. Discussion
The most commonly used serological methods of diagnosis to detect antibody response to APMV-1 and AIV infections in domestic poultry are by ELISA and HI assay; in addition the AGID test is used for detecting AIV antibodies. Although AGID and HI are gold standards, both assays are labor intensive and are less suited for high-throughput screening. The current project reports the development of multiplex immunoassay with the ability to simultaneously detect antibody responses to AIV and APMV-1 infections using the Luminex xMAP Technology.
The application of Luminex technology for the detection of antibody responses to multiple pathogens in a single assay format has been demonstrated previously (Anderson et al., 2011; Bergmann et al., 2003; Chen et al., 2009; Clavijo et al., 2006; Dias et al., 2005; Lupiani et al., 2010; Martins, 2002). Most of these developed assays used either recombinantly expressed proteins, or purified, or semi-purified antigens coupled to the beads. In the current study, expression of the AIV-NP and APMV-1-NC proteins were by using the baculovirus expression system. Producing the recombinant antigens in the baculovirus expression system has significant advantages including accurate glycosylation, speed and volume of protein expression, as well as proper folding (Invitrogen, Ontario, CA). Expressing the recombinant proteins fused to a His-tag had additional advantages in attaining a very clean protein product that helped minimize the occurrence of possible cross-reactions, lowered unwanted background signal in the assay, and also increased confidence of the results.
Many veterinary diagnostic laboratories currently use the IDEXX ELISA or other similar ELISAs for serologic screening of both AIV and APMV-1 disease infections in domestic poultry. The performance of the duplex FMIA was compared to the widely used IDEXX ELISA kits for both infections. In addition, for detecting AIV antibodies an in-house developed cELISA was used which employed identical capture antigen that was used in the FMIA (data not shown). Based on the comparative data presented in this study, the duplex FMIA assay performance was almost identical to the IDEXX ELISA kits for both diseases. Although the IDEXX ELISA kits or other commercially available ELISA kits allow for the processing of a very large number of samples in a relatively short period of time, they lack the multiplexing capabilities, and individual tests have to be carried out for each analyte to be detected. Therefore, ELISA testing becomes laborious and time consuming. The duplex FMIA reported here has a number of advantages as it is able to detect antibody responses to AIV and APMV-1 infection using a single serum dilution and also could easily be automated and used for processing of large amounts of serum submissions. The assay's potential to detect multiple targets in the same platform could offer substantial benefits for routine serological testing through reduced staff and reagent costs, and simplified laboratory operation.
As described previously by Martins (2002), the duplex FMIA assay also incorporates an additional two microspheres as internal controls that allow for the establishment of quality control parameters to ensure that each individual reaction has received the correct reagents. This allows the monitoring of whether the correct concentration of each individual reagent such as test sera, conjugate, and substrate has been added. The secondary antibody addition control ranged in from 9000 to 12,000 MFI on average. MFI values below 8000 were considered untrustworthy and repeated. The control for proper serum addition ranged from 6000 to 10,000 MFI on average. MFI values below 6000 were considered unreliable and were retested. Internal controls were tested on plates where serum or secondary antibody were intentionally left out or added incorrectly, and the controls successfully identified all problematic wells. Internal controls could also be utilized to increase precision and accuracy by monitoring instrument fluctuations, allowing intra- and inter-assay normalization (Martins, 2002).
The performance of the developed duplex FMIA was also compared with IDEXX ELISA kits in studying the kinetics of immune responses in chickens and turkeys that were infected with different subtypes of LPAI viruses. Based on these studies, in most infected animals, AIV antibodies could be detected as early as 4–7 dpi; reaching their peak by 14 dpi and remaining at a high level until 28 dpi. The results of duplex FMIA were almost identical to the IDEXX ELISA kit values for most of the groups infected with different subtypes of AIV. In addition, the performance of the assay was almost identical to the commercial ELISA platform when studying the kinetics of antibody responses in chickens immunized with the La'Sota APMV-1 vaccine. Antibodies to ND-NC could be detected as early as 4–7 dpi and remained at peak levels until 28 dpi. This study demonstrates the usefulness of the assay in monitoring antibody response in chickens or turkeys that were exposed to APMV-1 and AIV. The assay was also useful in monitoring antibodies in field serum samples collected from turkey breeders immunized with inactivated AIV and APMV-1.
The stability of AIV-NP and APMV-1-NC coupled beads stored at 4 °C was tested for a period of 4 months. The reference curve was used as a benchmark to track the sensitivity of the assay for this period of time, and four month old protein coupled beads have yet to show a drop in fluorescence values (data not shown).
In the duplex FMIA assay, there was limited cross reactivity between the APMV-1-NC antigen with APMV-7 and APMV-9 antisera. There is a possibility that this could be due to the presence of some conserved immunogenic antigenic sites on the nucleocapsid protein between these viruses and APMV-1 serotypes. Traditionally, APMVs are grouped into different serotypes (APMV-1 to AMPV-9) based on the HI test (Alexander, 1988), however, antibodies to prototype viruses representative of the nine currently defined APMV serotypes are already known to cross react with other APMV serotypes and with mammalian paramyxoviruses (Alexander and Chettle, 1978; Alexander and Collins, 1981; Kessler et al., 1979; Lipkind et al., 1982; Shortridge et al., 1980; Tumova et al., 1979). It is possible the APMV-1-NC may be too conserved among APMV serotypes to provide enough resolution for the exact classification.
A number of Luminex based assays have been developed for serologic diagnosis of avian diseases including AIV (Chowdhury et al., 2009; Drummond et al., 2008; Giavedoni, 2005; van Gageldonk et al., 2008; de Jager et al., 2005). To our knowledge this is the first study where a duplex FMIA was developed for serologic diagnosis and differentiation of two important poultry diseases. The time saving value of the FMIA even makes it a feasible option for high-throughput surveillance testing (Seideman and Peritt, 2002).
Receiver Operating Characteristic analysis (ROC) is a statistical non-parametric function that was used to determine the assay threshold values by analyzing the trade-off between true positive results and false positive results (Fawcett, 2006). Overall performance of the assay is consistent, and is based on a large data set. It is important to include a set of reference standards to allow for independent threshold determination without the need to normalize results. However, if the assay were to only be used with chicken and turkey serum from the same geographical region, once a suitable threshold value has been determined, there is no need to undergo the complex statistical analysis again. In its current state, the assay could be effectively used for surveillance of AIV and APMV-1 in poultry flocks. New work is under way to incorporate two additional bead sets which will allow for the specific differentiation of AIV infections of H5 or H7 subtypes. These are the notifiable forms of AIV and also have the capacity to become highly pathogenic strains.
Acknowledgments
The project was funded by CFIA (WIN-A-1304), Manitoba Egg Producers and MAFRI (Manitoba Agriculture, Food and Rural Initiatives). The authors would like to acknowledge Dr. Kathleen Hooper McGreevy and Animal Health Laboratory (AHL), Guelph, ON for providing the field sera used in the studies, Dr. Kurtis Swelka and his animal care staff for helping out with animal experiments, Dr. Andre Broes (Bio-Vet) for his input in assay development, and Dr. Gary Crow for his statistical analysis advice. CSF/Avian staff and Kate Hole for their help in some aspects of the assay validation.
References
Aldous, E.W., Alexander, D.J., 2001. Detection and differentiation of Newcastle disease virus (avian paramyxovirus type 1). Avian Pathol. 30, 117.
Alexander, D.J., 1988. Newcastle disease virus — an avian paramyxovirus. In: Alexander, D.J. (Ed.), Newcastle Disease. Kluwer Academic Publishers, Boston, MA, p. 11.
Alexander, D.J., Chettle, N.J., 1978. Relationship of parakeet/Netherlands/ 449/75 virus to other avian paramyxoviruses. Res. Vet. Sci. 25 (1), 105.
Alexander, D.J., Collins, M.S., 1981. The structural polypeptides of avian paramyxoviruses. Arch. Virol. 67 (4), 309. Alexander, D.J., Senne, D.A., 2003. Newcastle disease, other avian paramyxoviruses and pneumovirus infections: Newcastle disease, other avian paramyxoviruses, In: Saif, Y.M. (Ed.), Diseases of Poultry, 7th edn. Iowa State University Press, Ames, p. 63.
Anderson, S., Wakeley, P., Wibberley, G., Webster, K., Sawyer, J., 2011. Development and evaluation of a Luminex multiplex serology assay to detect antibodies to bovine herpes virus 1, parainfluenza 3 virus, bovine viral diarrhoea virus, and bovine respiratory syncytial virus, with comparison to existing ELISA detection methods. J. Immunol. Methods 366, 79.
Bergmann, I.E., Neitzert, E., Malirat, V., Ortiz, S., Colling, A., Sanchez, C., Correa Melo, E., 2003. Rapid serological profiling by enzyme-linked immunosorbent assay and its use as an epidemiological indicator of foot-and-mouth disease viral activity. Arch. Virol. 148, 891.
Berhane, Y., Kehler, H., Handel, K., Hisanaga, T., Xu, W., Ojkic, D., Pasick, J., 2012. Molecular and antigenic characterization of reassortant H3N2 viruses from turkeys with a unique constellation of pandemic H1N1 internal genes. PLoS One 7 (3), e32858.
Chen, T., Lee, F., Lin, Y., Pan, C., Shih, C., Lee, M., Tsai, H., 2009. Development of a Luminex assay for the detection of swine antibodies to non-structural proteins of foot-and-mouth disease virus. J. Immunol. Methods 396, 87–95.
Chowdhury, F., Williams, A., Johnson, P., 2009. Validation and comparison of two multiplex technologies, Luminex® and Mesoscale Discovery, for human cytokine profiling. J. Immunol. Methods 340, 55.
Clavijo, A., Hole, K., Li, M., Collingnon, B., 2006. Simultaneous detection of antibodies to foot-and-mouth disease non-structural proteins 3ABC, 3D, 3A and 3B by a multiplexed Luminex assay to differentiate infected from vaccinated cattle. Vaccine 24, 1693.
Comin, A., Toft, N., Stegeman, A., Klinkenberg, D., Marangon, S., 2013. Serological diagnosis of avian influenza in poultry: is the haemagglutination inhibition test really the ‘gold standard’? Influenza Other Respi. Viruses 7, 257.
de Jager, W., Prakken, B.J., Bijlsma, J.W.J., Kuis, W., Rijkers, G.T., 2005. Improved multiplex immunoassay performance in human plasma and synovial fluid following removal of interfering heterophilic antibodies. J. Immunol. Methods 300, 124.
Desvaux, S., Garcia, J.M., Nguyen, T.D., Reid, S.A., Bui, N.A., Roger, F., Fenwick, S., Peiris, J.S., Ellis, T., 2012. Evaluation of serological tests for H5N1 avian influenza on field samples from domestic poultry populations in Vietnam: consequences for surveillance. Vet. Microbiol. 156, 277.
Dias, D., Van Doren, J., Schlottmann, S., Kelly, S., Puchalski, D., Ruiz, W., Boerckel, P., Kessler, J., Antonello, J.M., Green, T., Brown, M., Smith, J., Chirmule, N., Barr, E., Jansen, K.U., Esser, M.T., 2005. Optimization and validation of a multiplexed Luminex assay to quantify antibodies to neutralizing epitopes on human papillomaviruses 6, 11, 16, and 18. Clin. Diagn. Lab. Immunol. 12, 959.
Drummond, J.E., Shaw, E.E., Antonello, J.M., Green, T., Page, G.J., Motley, C.O., Wilson, K.A., Finnefrock, A.C., Liang, X., Casimiro, D.R., 2008. Design and optimization of a multiplex anti-influenza peptide immunoassay. J. Immunol. Methods 334, 11.
Fawcett, T., 2006. An introduction to ROC analysis. Pattern Recogn. Lett. 861. Giavedoni, L.D., 2005. Simultaneous detection of multiple cytokines and chemokines from nonhuman primates using Luminex technology. J. Immunol. Methods 301, 89.
Hoffmann, E., Stech, J., Guan, Y., Webster, R.G., Perez, D.R., 2001. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 146, 2275.
Kapczynski, D.R., Afonso, C.L., Miller, P.J., 2013. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 41 (3), 447.
Kessler, N., Aymard, M., Calvet, A., 1979. Study of a new strain of paramyxoviruses isolated from wild ducks: antigenic and biological properties. J. Gen. Virol. 43, 273.
Leijon, M., Ullman, K., Thyselius, S., Zohari, S., Pedersen, J.C., Hanna, A., Mahmood, S., Banks, J., Slomka, M.J., Belak, S., 2011. Rapid PCR-based molecular pathotyping of H5 and H7 avian influenza viruses. J. Clin. Microbiol. 49, 3860.
Lipkind, M., Weisman, Y., Shihmanter, E., Shoham, D., 1982. Isolation of yucaipalike avian paramyxovirus from a wild mallard duck (Anas platyrhinchos) wintering in Israel. Vet. Rec. 110 (1), 15.
Lupiani, B., Mozisek, B., Mason, P.W., Lamichhane, C., Reddy, S.M., 2010. Simultaneous detection of avian influenza virus NP and H5 antibodies in chicken sera using a fluorescence microsphere immunoassay. Avian Dis. 54, 668.
Martins, T.B., 2002. Development of internal controls for the Luminex instrument as part of a multiplex seven-analyte viral respiratory antibody profile. Clin. Diagn. Lab. Immunol. 9, 41.
Mutinelli, F., Capua, I., Terregino, C., Cattoli, G., 2003. Clinical, gross, and microscopic findings in different avian species naturally infected during the H7N1 low- and high-pathogenicity avian influenza epidemics in Italy during 1999 and 2000. Avian Dis. 47, 844.
Pasick, J., Pedersen, J., Hernandez, M.S., 2012. Avian influenza in North America, 2009–2011. Avian Dis. 56, 845.
Pawar, S.D., Kale, S.D., Rawankar, A.S., Koratkar, S.S., Raut, C.G., Pande, S.A., Mullick, J., Mishra, A.C., 2012. Avian influenza surveillance reveals presence of low pathogenic avian influenza viruses in poultry during 2009–2011 in the West Bengal State, India. Virol. J. 9 (151-422X-9-151).
Seal, B.S., Crawford, J.M., Sellers, H.S., Locke, D.P., King, D.J., 2002. Nucleotide sequence analysis of the Newcastle disease virus nucleocapsid protein gene and phylogenetic relationships among the Paramyxoviridae. Virus Res. 83, 119.
Seideman, J., Peritt, D., 2002. A novel monoclonal antibody screening method using the Luminex-100™ microsphere system. J. Immunol. Methods 267, 165.
Shortridge, K.F., Alexander, D.J., Collins, M.S., 1980. Isolation and properties of viruses from poultry in Hong Kong which represent a new (sixth) distinct group of avian paramyxoviruses. Res. Vet. Sci. 255.
Snyder, D.B., Marquardt, W.W., Yancey, F.S., Savage, P.K., 1985. An enzymelinked immunosorbent assay for the detection of antibody against avian influenza virus. Avian Dis. 29, 136.
Toffan, A., Olivier, A., Mancin, M., Tuttoilmondo, V., Facco, D., Capua, I., Terregino, C., 2010. Evaluation of different serological tests for the detection of antibodies against highly pathogenic avian influenza in experimentally infected ostriches (Struthio camelus). Avian Pathol. 39, 11.
Tumova, B., Robinson, J.H., Easterday, B.C., 1979. A hitherto unreported paramyxovirus of turkeys. Res. Vet. Sci. 135.
Upadhyay, C., Ammayappan, A., Vakharia, V.N., 2009. Detection of NP, N3 and N7 antibodies to avian influenza virus by indirect ELISA using yeastexpressed antigens. Virol. J. 6 (158-422X-6-158).
van Gageldonk, P.G.M., van Schaijk, F.G., van der Klis, F.R., Berbers, G.A.M., 2008. Development and validation of a multiplex immunoassay for the simultaneous determination of serum antibodies to Bordetella pertussis, diphtheria and tetanus. J. Immunol. Methods 335, 79.
Wang, L., Pan, C., Severinghaus, L.L., Liu, L., Chen, C., Pu, C., Huang, D., Lir, J., Chin, S., Cheng, M., Lee, S., Want, C., 2008. Simultaneous detection and differentiation of Newcastle disease and avian influenza viruses using oligonucleotide microarrays. Vet. Microbiol. 127, 217.
Watson, D.S., Reddy, S.M., Brahmakshatriya, V., Lupiani, B., 2009. A multiplexed immunoassay for detection of antibodies against avian influenza virus. J. Immunol. Methods 340, 123.
World Livestock Disease Atlas, Bank, W., 2011. A Quantitative Analysis of Global Animal Health Data. (2006-2009). Washington, DC. http://www. oie.int/doc/ged/D11291.pdf. Accessed February 24, 2014. p35, 39.
World Organization for Animal Health, 2008. Manual of diagnostic tests and vaccines for terrestrial animals, Sixth edition. OIE, Paris, France. XLSTAT, Addinsoft, 2013. Data Analysis and Statistics with MS Excel. Addinsoft, NY, USA.
Yang, M., Berhane, Y., Salo, T., Li, M., Hole, K., Clavijo, A., 2008. Development and application of monoclonal antibodies against avian influenza virus nucleoprotein. J. Virol. Methods 147 (2), 265.
Zhou, E.M., Chan, M., Heckert, R.A., Riva, J., Cantin, M.F., 1998. Evaluation of a competitive ELISA for detection of antibodies against avian influenza virus nucleoprotein. Avian Dis. 42, 517.
Related topics
Authors:


Join to be able to comment.
Once you join Engormix, you will be able to participate in all content and forums.
* Required information
Would you like to discuss another topic? Create a new post to engage with experts in the community.
Create a post
University of Prince Edward Island
17 de marzo de 2015
I really appreciate your comments. Regards

17 de marzo de 2015
dear sir,
very nice article with new technology for differential diagnosis. Avian Influenza virus has gained importance very recently say 15years but ND virus is very old and are controlled through vaccination in poultry. ND can cause conjunctivitis in human being some time but AI virus there is no definite trend of zoonotic importance and any infection to human being said to create pandemic But there are reports widely penetrating every where the importance of AI virus.
what is the future of the poultry production on this globe when scientist have helped the state management to handle AI virus at the state cost. I do not see any good sign the way the issue is taken up every where on AI virus and perhaps it is fitness to eliminate chicken , turkey , ducks from commercial production when all are disturbed with apprehensions and suspicions. Let the migratory birds and wild birds be there on this planet.AI virus handling is getting expensive. Let us find out answer to such issue when international trade is now guided by such AI virus in poultry sector.
The article is very interesting thank you sir.







You may be interested in
.jpg&w=3840&q=75)