Antioxidants in nutrition of horses
Antioxidants in physiology and nutrition of exercising horses
Endogenous antioxidants and those provided by the diet work as part of a coordinated and balanced team, to help counteract the potential deleterious effects of free radicals and reactive oxygen species (ROS) on tissues and body fluids. Free radicals are chemical species that have an unpaired electron, making them very ‘reactive’ (Table 1). This extreme reactivity, which underlies their ability to interact with and ultimately damage tissues, is driven by a desire to acquire another electron therefore becoming paired and more stable. Some radicals do not have an unpaired electron and are comparatively less reactive, although still capable of damage. ROS, as the name suggests, are oxygen containing species that may or may not have an unpaired electron but are nevertheless reactive and potentially damaging.
Free radicals and ROS are formed as by-products of normal metabolism and are capable of damaging DNA, lipids, proteins and carbohydrates (Evans and Halliwell, 2001). The superoxide radical (•O2 -), for example is formed during normal mitochondrial respiration, and hydrogen peroxide (H2O2) can be formed during the degradation of neurotransmitters such as dopamine in the brain.
Table 1. Reactive oxygen and related species.

Adapted from Noguchi and Niki, 1999 However, not all production of these ‘reactive’ species occurs accidentally, as the body can and does use these substances for its own benefit. For example, nitric oxide (NO•) functions as a regulator of vascular tone and as a messenger in the central nervous system. Immune function is also dependent on the targeted use of these reactive species by activated neutrophils and macrophages in killing invading bacteria and fungi. So free radicals and ROS can be useful, but only when produced in controlled amounts and in the ‘right place at the right time’. Uncontrolled production of certain reactive species occurs in several diseases in man, for example, rheumatoid arthritis or inflammatory bowel disease (Evans and Halliwell, 2001).
Under normal conditions the deleterious effects of ROS are counteracted by the body’s antioxidant defences, which are contributed to through dietary intake of key nutrients. Antioxidants are effective in a number of ways:
- Preventive antioxidants: These suppress the formation of free radicals; for example catalase and glutathione peroxidase, two antioxidant enzymes, decompose hydrogen peroxide, preventing the formation of oxygen radicals.
- Free radical scavengers: These antioxidants confer stability to the ‘reactive’ species by donation of an electron and become oxidised themselves to form a more stable radical. For example α-tocopherol (vitamin E) scavenges peroxyl radicals and is converted to a tocopherol radical. To illustrate the interaction between antioxidants, these tocopherol radicals can then be ‘re-activated’ by ascorbic acid, which in turn forms an ascorbate radical in the process.
- Sequestration of metal by chelation: Metals such as iron and copper can propagate the formation of more reactive radicals and so are kept bound to transport proteins such as transferrin or ceruloplasmin, as this renders them less available to contribute to radical formation.
- Quenching of active oxygen species: Antioxidants can convert active oxygen species to more stable forms, for example, carotenoids and vitamin E stabilise singlet oxygen radicals, forming less reactive hydrogen peroxide.
Table 2. Endogenous and diet-derived components of the antioxidant system.
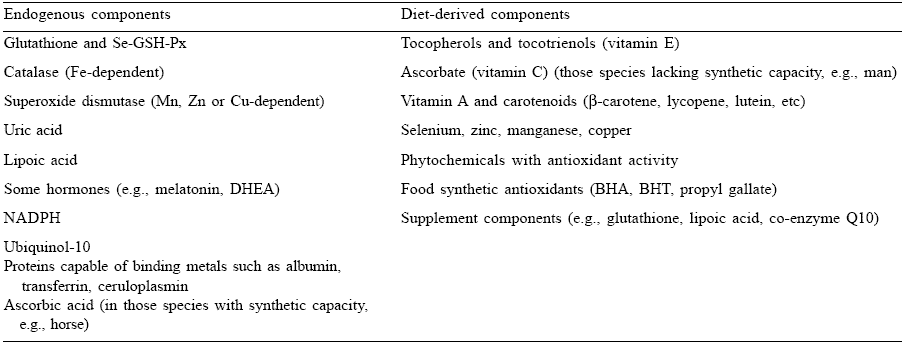
To enlarge, click here
Papas, 1999b
Table 3. Collaboration between the major components of the antioxidant system.
To enlarge, click here
Antioxidants are also positioned in specific cellular and tissue locations and so offer comprehensive and coordinated defence against ROS and free radicals. Glutathione peroxidase, a selenium-dependent antioxidant enzyme, is located in the mitochondria and cytosol and removes hydrogen peroxide and organic hydroperoxides. Vitamin E, which is a fat soluble vitamin, functions as a membrane-bound antioxidant, trapping lipid peroxyl free radicles produced from unsaturated fatty acids under conditions of ‘oxidative stress’. The orientation of vitamin E within cell membranes appears to be critical to its functionality. Vitamin E is an amphipathic molecule in that it has a structure that incorporates hydrophobic (water-hating) and hydrophilic (water-loving) parts. This characteristic facilitates its orientation and retention within the lipid bilayer of cell membranes, probably giving it an ideal position for free radicle scavanging and thus protection of cell membranes. Vitamin C is located in the aqueous phase of cells, where it contributes to radical scavenging and is also thought to regenerate vitamin E, through its own oxidation from ascorbic acid to dehydroascorbic acid. Glutathione, which is found in all cells, works in coordination with GSH-Px as a multifunctional antioxidant.
The body’s antioxidant system depends on the supply of a number of its nutrient components, including vitamins and trace minerals, from the diet. Whilst some of these nutrients, such as vitamin C and E, have direct antioxidant properties, others, such as selenium or copper, are required as cofactors for the activity of key antioxidant enzymes. There are also a number of natural components of plants, cereals and pulses, which will be discussed below that exhibit antioxidant properties. Major dietary antioxidants
VITAMINS
Vitamin E
The term ‘vitamin E’ refers to a group of compounds known as tocopherols, with the most biologically active form being α-tocopherol. Vegetable oils are the richest source of natural vitamin E and the level and type of tocopherol varies between different oil sources (Table 4). These natural tocopherols largely act as internal antioxidants to prevent oxidation of unsaturated fatty acids that make up the constituent triglyceride and hence prevent rancidity of the oil. However, they may also have antioxidant properties in vivo. The level of vitamin E in an oil appears to be positively correlated to its polyunsaturated fatty acid (PUFA) content, specifically the combined amounts of linoleic and α-linolenic fatty acids. Horses appear to absorb natural forms of vitamin E to a much greater extent than synthetic versions (Gansen et al., 1995). When the absorption of different sources of vitamin E were compared in thoroughbred yearlings, higher plasma concentrations were reported with the d-forms compared to equivalent amounts of the dl-form (Wooden and Papas, 1991).
Table 4. Typical vitamin E content of fats and oils (mg/100 g oil). Total tocopherol is the sum of α-tocopherol and γ-tocopherol.

Dupont et al., 1990
Previous studies in human subjects suggested that the requirement for vitamin E increased with increasing dietary PUFA content (Wardlaw, 1999). Supplementation of the equine diet with oil may also therefore bring with it an increased requirement for antioxidant provision including vitamin E. The increased fat oxidation that occurs during submaximal exercise following fat supplementation (Dunnett et al., 2002) is likely due to increased production of peroxyl free radicles, hence the need for antioxidant support . This increased requirement for vitamin E may in part be satisfied by the natural tocopherol content of the oil concerned, but can also require specific antioxidant supplementation.
Both human and animal studies suggest that the requirement for vitamin E is estimated to be 0.6 mg of α-tocopherol per g of linoleic acid, which equates to an additional 36 mg of vitamin E per 100 g of vegetable oil added. For most animal species, 3 mg of α-tocopherol is recommended for each 1 g of ω- 3 polyunsaturated fatty acid (Vitec, 1987). For soya or corn oil, this would equate to 180 mg per 100 g of oil. However, both soya and corn oil have an endogenous tocopherol content of about 60 mg/100 g, leaving a net requirement for 120 mg/100 g of oil or 108 mg vitamin E per 100 ml of oil. Care should, however, be taken to ensure that vitamin E levels are adequate to account for the shelf life of the product, since endogenous tocopherol levels may be substantially reduced during storage without additional antioxidant support.
Horses fed a diet supplemented with 6.4% soyabean oil and containing current NRC recommended levels of vitamin E (80 IU/kg DM) showed no significant change in the ratio of α- tocopherol to total lipid (serum cholesterol + triglyceride) compared to an isoenergetic control diet (Siciliano and Wood, 1993). This may indicate that either the current requirements for vitamin E were high enough to cope with the level of addition of PUFA-containing oil or that the endogenous tocopherol content of the oil itself diminished the need for further supplementation. However, the work of McMeniman and Hintz (1992) suggests that further supplementation may be required to maintain normal antioxidant balance. They reported no reduction in plasma or muscle vitamin E status in ponies fed a diet where 10% of the total digestible energy was provided in the form of corn oil. However, in the same study plasma thiobarbituric acid reactive substances (TBARs) and breath pentane, which give an indication of oxidative stress, were increased during exercise, especially in those horses with a relatively low plasma vitamin E concentration.
Human studies have suggested a link between vitamin E status and health. Epidemiological studies and short-term intervention studies propose that vitamin E, mostly in the form α-tocopherol, may reduce the risk of heart disease, some forms of cancer and cataracts amongst other diseases, as well as reducing exercise induced muscle damage. These benefits do not appear to have resulted from correction of a vitamin E deficiency, but follow hyper-supplementation of vitamin E at levels reaching 1-30 times the recommended daily allowance (Papas, 1999b).
Vitamin C
Vitamin C (ascorbic acid) is a weak acid and a potent antioxidant and hence reducing agent. In chemical terms, this means that ascorbic acid easily gives up electrons to provide stability to reactive species such as ROS. The term vitamin C actually refers to a number of related compounds that have the biological activity of L-ascorbic acid. Vitamin C is essentially a scavenger of ROS (e.g., hydroxyl radicals), as well as antioxidant-derived radicals (i.e., produced when antioxidants such as α-tocopherol or uric acid carry out their antioxidant function, themselves becoming oxidised). In other words, along with a direct antioxidant function, ascorbic acid is also able to recycle or regenerate other components of the antioxidant system, particularly vitamin E, and in doing so becomes oxidised itself allowing reduction and hence regeneration of the antioxidant concerned.
Unlike humans, but like most other vertebrates, the horse under normal circumstances has no dietary requirement for ascorbic acid, as it is synthesised in the liver ultimately from glucose. However, research suggests that there may be cause for supplementation in times of stress or where disease is present. In this situation, the body’s ability to synthesise vitamin C may not be sufficient to satisfy the increased demand in response to, for example, age-related conditions, environmental irritants or pollutants or disease, especially when combined with a poor quality diet.
In support of this concept, Hoffman et al. (1995) reported that foals exhibiting high levels of stress during weaning due to confinement in stalls exhibit lower than normal plasma ascorbate levels. Furthermore, vitamin C supplementation (20 g per day) has been reported to improve antibody response to vaccine in aged horses, particularly those with Cushing’s disease, and may benefit those with chronic infections (Ralston, 1999).
Although there are no reports of vitamin C toxicity in horses (NRC, 1989), there remains some controversy over the relative merits of hypersupplementation. When present in excess, ascorbic acid may have pro-oxidant properties (i.e., the ability to promote the formation of free radicals), especially in the presence of an excess of iron that is not bound to transferrin (Halliwell, 1994).
Absorption of vitamin C is reported to occur by passive diffusion in the ileum of the horse (Lewis, 1995). However, in humans absorption is described to occur via a sodium-dependent active transport mechanism (Rumsey et al., 1999). Uptake of vitamin C in the horse appears to be affected by both the level and form in which it is supplied. Although plasma levels of vitamin C were elevated by feeding 20 g of ascorbic acid daily, they were not affected by a single 20 g administration (Snow and Frigg, 1987). Absorption of ascorbic acid appears to be poor, and other forms of vitamin C such as ascorbyl palmitate are more bioavailable (Deaton et al., 2003; Snow and Frigg, 1989; Snow and Frigg, 1990). There also appears to be significant variation in bioavailability of vitamin C between individuals (Deaton et al., 2003; Snow and Frigg, 1989).
Commercial preparations of vitamin C are now frequently obtained as salts of ascorbic acid, commonly as sodium or calcium ascorbate, traditionally prepared from ascorbic acid and sodium or calcium carbonate by controlled precipitation in dilute acetone or alcohol (Goodman, 2003). There are now, however, forms of vitamin C available that use a new process for the manufacture of calcium ascorbate. This process, in contrast to other methods, does not use solvents such as alcohol or acetone and instead, the entire process is carried out in water. These preparations contain non-acidic mineral ascorbates and other ascorbic acid metabolites, which are reputed to improve absorption. This, together with the more neutral pH and buffering power of ascorbate salts compared with ascorbic acid, is frequently cited as a superior attribute of these ascorbate products. Other vitamin C products claiming to offer improved stability during feed manufacture include those composed of a mixture of the phosphate esters of L-ascorbic acid. Bioavailability of this latter form of vitamin C in horses is marginally greater than ascorbic acid but reduced in comparison to ascorbyl palmitate (Deaton et al., 2003).
TRACE MINERALS
There are a number of trace minerals provided by the diet that have an essential role in supporting the function of endogenous antioxidant enzymes. For example, the trace minerals copper, manganese, zinc, iron and selenium are all co-factors for specific antioxidant enzymes namely, superoxide dismutase (SOD-Cu, Mn, Zn), catalase (Fe), and glutathione peroxidase (GSH-Px) (Se).
Although these trace minerals are therefore important dietary constituents, it is important to note that they can also act as pro-oxidants (promote the formation of free radicals). This pro-oxidant activity is due to their ability to convert from one oxidation state to another and in doing so either accept or donate electrons, which can result in the formation of more potent ROS, as in the example below. However, their pro-oxidant activity is largely suppressed as these trace minerals are mainly transported in the body bound to proteins such as haemoglobin, transferrin and ceruloplasmin.
Fe2+ + H2O2 Fe3+ + HO• Fenton reaction
Selenium
Selenium is the trace mineral that has been studied in the horse in greatest detail with respect to its antioxidant function. It is a component of a number of enzyme-based antioxidants, which are selenoproteins such as the GSH-Px family of enzymes, giving dietary selenium an indirect antioxidant role. The antioxidant role of this enzyme group is to reduce hydroperoxides and lipid peroxides, becoming oxidised themselves in the process. There have been five tissue-specific GSH-Px enzymes identified in mammals (see Surai, this volume).
The selenium content of vegetable feed ingredients is largely dependent on the underlying selenium content of the soil on which the particular crop is grown. There are some regions of the world, including parts of New Zealand, the western and eastern parts of the US and Scotland in the UK, where the selenium content of the soil is low (Frape, 1998). In the UK the low selenium content of the soil is reflected in the reported decline in blood selenium concentration in humans in recent years (Rayman, 1997). However, equally there are regions in the world with high selenium soils where selenium toxicity has been described in horses, with clinical signs including loss of mane and tail hair and sloughing of hooves (McLaughlin and Cullen; 1986, Schryver, 1990). The maximum tolerable level of selenium is reported to be 2 mg/kg of diet (NRC, 1989), whilst the toxic level for horses, according to Schryver (1990) may vary from 5-40 mg/kg in the diet.
In forages, cereal grains and oilseed meals, selenium is present in organic form, largely as selenomethionine but also as selenocystine and selenocysteine (Schrauzer, 2000), which can be synthesised by microorganisms and plants, but not by higher animals and man. Historically, horse feed has been supplemented with selenium in the form of sodium selenite. However, it appears that the form in which selenium is ingested can influence its behaviour and activity within the body. Pagan et al. (1999) measured selenium utilisation in two diets containing 0.41 mg/kg selenium, either in the form of sodium selenite or as a selenium-rich yeast (Sel- PlexTM, Alltech Inc). The apparent digestibility of yeast selenium (mainly in the form of selenomethionine) was increased above that of selenite (57% vs 51%), and selenium retention (plasma increase minus urinary and faecal losses) was again greater for the yeast compared to the inorganic source.
Whilst the National Research Council recommends a minimum intake of selenium of 0.1 mg/kg diet dry matter (NRC, 1989), other researchers recommend higher levels (0.1–0.3 mg/ kg) (Pagan, 1999). However, the optimum dietary intake needed to support antioxidant function during times of stress or disease remains unresolved. A decrease in blood glutathione concentration reported during a 45-day training study suggests that repetitive exercise increases pressure upon the antioxidant system as a result of ROS produced (Shelle et al., 1985). However, despite this reduction in glutathione, the activity of erythrocyte GSH-Px was reported to increase during training in the same study, which may represent up-regulation in response to the exercise stimulus. Dietary selenium availability is likely to influence this exercise-induced increase in GSH-Px activity, since the increase in GSH-Px in this study was greater when these horses were supplemented with an additional 2.5 mg Se/day (Shelle et al., 1985). However, this response will depend on the basal intake of selenium before supplementation, but suggests that adequate selenium intake becomes more important during training.
OTHER DIETARY ANTIOXIDANTS: PHYTOCHEMICALS
There are a large number of phytochemicals not recognised as having a nutritive value that are believed to have antioxidant properties. Most feed ingredients will contain some of these phytochemicals, but fruits, vegetables and herbs are regarded as the richest sources. The major classes of phytochemicals with potential for antioxidant activity are listed in Table 5.
Table 5. Major classes of phytochemicals with antioxidant activity.

Papas, 1999a
Oxidative stress
Antioxidant status represents the balance between the antioxidant system and the pro-oxidants (including free radicals and ROS) in the body. This balance is dynamic, and in humans is tipped slightly in favour of oxidation, which allows for normal metabolism (Evans and Halliwell, 2001). The body has adapted to this slight favouring of oxidation by developing repair mechanisms for damage created. In addition, this situation induces production of endogenous antioxidants and so essentially offers a type of feedback arrangement (Evans and Halliwell, 2001). However, when this balance is seriously tipped in favour of oxidation, oxidative stress has occurred. Oxidative stress may be due to an increased production of free radicals or ROS, which can occur as a result of disease or infection, or as a consequence of exercise, where an increased demand for ATP increases throughput through the aerobic energy-generating pathways and hence electron transport chain. Oxidative stress may be exacerbated when dietary antioxidant intake is reduced, or where intake of trace mineral co-factors is insufficient to facilitate antioxidant support. Although micronutrient deficiencies may make animals more susceptible to oxidative stress, this may only become significant during infection or disease.
Oxidative stress can result in net damage to tissues and other body chemicals and has been implicated in a number of disease states in man including atherosclerosis, cancer, rheumatoid arthritis, inflammatory bowel disease and asthma (Halliwell, 1994). In addition, exercise-induced oxidative stress is believed to contribute to poor performance and even exercise intolerance, mediated via the action of ROS on muscle, contributing to muscle fatigue and damage (Blumberg and Halpner, 1999). Oxygen uptake in exercising horses may increase 30-fold compared to resting rates (Butler et al., 1993). This increased oxygen profusion in muscle is proposed to indirectly lead to localised muscle fibre damage due to the resultant production of ROS and their subsequent disruption of muscle fibre membranes. Oxidative stress has been demonstrated to occur during exercise in the horse, but its presence and degree seems to be dependent on both the exercise intensity and duration as well other ‘stress’ factors such as temperature and humidity (Mills et al., 1996). Endurance exercise is characterised as having a high reliance on aerobic energy-generating pathways, which is likely to increase the exerciseinduced generation of ROS. Hargreaves et al.
(2002) proposed that the antioxidant defences of endurance horses are severely tested during a race and that the degree of oxidative stress developed may be related to subsequent muscle damage. They reported evidence of associated changes in antioxidant status and markers of muscle cell leakage during two endurance races of 80 and 160 km distance. Antioxidant status was characterised by a reduction in the plasma vitamin C and glutathione concentration, while circulating levels of vitamin E were maintained, presumably as a result of further release from adipose tissue stores concomitant with the mobilisation of free fatty acids from adipose tissue triglycerides. The concentration of vitamin E is also likely to have been buffered by its regeneration by vitamin C. However, despite the reported decline in the levels of individual antioxidants, they were only weakly correlated to observed increases in aspartamine transaminase (AST) and creatine kinase (CK), two markers of muscle cell damage (r = 0.36-0.67; P<0.03) (Hargreaves et al., 2002).
In addition, a similar study by Marlin et al. (2002) reported that despite quite marked variation in individual antioxidant concentration pre-race, there was no clear relationship with subsequent increases in either AST or CK. These authors suggest that the relationship may be more complicated and requires comparisons to be made between markers of muscle damage and an index of total antioxidant capacity rather than with individual components. In agreement with Hargreaves et al. (2002), plasma vitamin E levels were maintained during this 140 km endurance race, as were the levels of ascorbic acid, which did not decline until 16 hrs of recovery.
The relationship between oxidative stress and exercise-induced muscle damage is complex, as it will be affected by degree of oxidative stress developed, which is affected by factors such as individual variation in antioxidant status, dietary intake of antioxidants and intensity and duration of exercise. Marlin et al. (2002) concluded that although the group of horses studied showed evidence of lipid peroxidation as indicated by an increase in plasma TBARS, as well as changes in some antioxidants, the results were not indicative of marked oxidative stress. Additionally, it was difficult to assess the effect of diet as details of the basal rations were unknown.
OXIDATIVE STRESS AND DISEASES OF CONSEQUENCE TO HORSES IN TRAINING
Oxidative stress reputedly plays a role in the aetiology of many diseases in man as described earlier. Likewise, in the horse the relevance of oxidative stress in a number of conditions is also now being recognised. An association has been proposed between oxidative stress and exerciseinduced pulmonary haemorrhage (EIPH) (Derksen, 1997) and muscle myopathy (Perkins et al., 1998) as well as joint disease (Dimock et al., 2000) and recurrent airway obstruction (known as COPD or heaves) (Kirschvink et al., 2002). The oxygen-rich environment in the lungs leaves this tissue particularly susceptible to damage from ROS, and strong antioxidant defences led by vitamin C and glutathione are vital. Vitamin C level in lung fluids is reduced in horses with airway inflammation (Deaton et al., 2003).
In man, several studies have suggested that supplementation with individual antioxidants or indeed cocktails can ameliorate oxidative stress during exercise by increasing the body’s antioxidant capacity and thus its ability to manage ROS (Blumberg and Halpner, 1999). The effect of antioxidant supplementation in horses has been investigated, but often with inconclusive results. Intravenous administration of 5 g of ascorbate to thoroughbreds prior to racing has been reported to nullify the oxidative stress response to maximal exercise resulting in a reduction in markers of oxidative stress (White et al., 2001). Whilst most of the supplementation studies in horses have concentrated on administration of single antioxidants such as vitamin C or E, there could be a rationale for use of mixtures or ‘cocktails’ as they may have a synergistic effect in vivo. Deaton et al. (2002) investigated the effect of supplementation with a cocktail of antioxidants consisting of a mixture of natural plant-based antioxidants, together with vitamins C and E as well as selenium. Using a crossover design, horses in this study were fed a basal control diet, supplemented with either the antioxidant cocktail or placebo for four weeks. Total intake of vitamins C and E and selenium in the supplemented diet was 458 IU/kg, 525 mg/kg and 0.31 mg/kg of feed, respectively. The basal diet in contrast provided 204 IU of vitamin E and and 0.14 mg of selenium per kg of feed. The results of this study indicated a significant increase (40%) in the plasma concentration of reduced ascorbic acid and α-tocopherol, both at rest and during subsequent exercise, following antioxidant supplementation compared to the placebo. However, the effect of this supplementation on oxidative stress could not be assessed since the exercise tests failed to elicit any significant oxidative stress in either the antioxidant- or placebo-supplemented horses despite the exercise being intense (90% VO2 max).
The quantity of vitamin E and selenium provided by the basal diet in this study significantly exceeded the NRC minimum recommendations for horses in hard work (NRC, 1989). In this situation, it seems likely that the endogenous antioxidant capacity was able to suppress the development of significant oxidative stress during exercise. The additional benefit of feeding an antioxidant-based supplement, where requirements are met by the basal diet, may be limited in normal horses. However, supplementation may become more important when intake from the diet is insufficient, or where the production of ROS is greater, for example, as the result of disease or where exercise intensity or duration is increased. In the absence of vitamin and mineralised feed or supplements, the horse’s traditional diet, although rich in β-carotene, a chain breaking antioxidant, may lack sufficient quantities of other antioxidants such as vitamin C and E, as well as some of the trace minerals required by specific antioxidant enzymes.
Fresh grass ensures a relatively high intake of β- carotene, as do some other traditional forages such as alfalfa. However, the level of carotenoid substances in conserved forage is likely to be much lower as long-term stability is affected by exposure to light and is dependent on the surface area exposed (Boileau et al., 1999). The carotenoids are a group of over 600 naturally occurring plant pigments, that contribute to the yellow, orange and red colours seen in many fruits, vegetables and forages. β-carotene, the most well known carotenoid, is also a precursor of vitamin A (retinol). The carotenoids are described as good chain-breaking antioxidants (Boileau et al., 1999). Free radicals generated within the body can react with PUFA, then causing a chain reaction of radical formation capable of destroying many lipid molecules in a short period of time. Chain-breaking antioxidants are capable of reducing unstable radical species by donating hydrogen to confer stability. Carotenoids are also able capable of stabilizing singlet oxygen species thus reducing their damaging effects (Boileau et al., 1999).
Soya, which is another ingredient widely used in the manufacture of horse feeds, is a rich source of compounds known as flavones, particularly diadzein and genistein. These compounds have a structural similarity to oestrogen and are thought to have antioxidant properties (Brouns, 2000).
Rice bran has gained in popularity amongst horse owners and trainers in recent years due to its high oil and low starch content. However, this ingredient may also have other benefits as it contains many beneficial natural antioxidants including tocopherols, tocotrienols, carotenoids and oryzanols (Jariwalla, 2001). γ-Oryzanol is a nutritionally important constituent of rice bran oil and is normally present at a level of about 1-3% (Lloyd et al., 2000). However, some of the γ-oryzanol is lost during processing and commercially available rice bran meal products quote levels of about 0.25% or 245 mg/100 g. γ-Oryzanol is in fact not a single compound, but a mixture of ferulic acid esters of triterpene alcohols and plant sterols. These components of the γ-oryzanol have a number of reputed effects including antioxidant activity (Jariwalla, 2001).
There are also many other novel raw materials available for use in horse feed such as grapeseed extract, citrus and vegetable extracts and traditional herbs and spices with reputed antioxidant properties. However, in many cases these antioxidant properties have been conferred in an in vitro situation, and their effects cannot always be extrapolated in vivo (Halliwell, 2002). A high intake of fruit and vegetables is proven to have a protective effect against some diseases involving the cardiovascular and respiratory systems and has even been cited to offer protection against certain forms of cancer in man. However, studies in which an attempt has been made to isolate the components responsible in the food concerned and then supplement them outside of the fruit and vegetable matrix have largely failed to produce consistent results. This suggests that the interaction between different components of feed ingredients in the natural state is complex.
Acknowledgments
Thanks to Dr David Marlin, Head of Physiology at The Animal Health Trust, Newmarket UK for help in the preparation of this review.
References
- Blumberg, J.B. and A.D. Halpner. 1999. Antioxidant status and function: Relationship to aging and exercise. In: Antioxidant Status, Diet, Nutrition and Health (A.M. Papas, ed). CRC Press, Boca Raton, pp. 251-275.
- Boileau, T.W.M., A.C. Moore and J.W. Erdman. 1999. Carotenoids and vitamin A. In: Antioxidant Status, Diet, Nutrition and Health (A.M. Papas, ed). CRC Press, Boca Raton, pp. 89-106.
- Brouns, F. 2000. Soya isoflavones: a new and promising ingredient for the health food sector. Agro Food Industry, Nov/Dec p. 12-17.
- Butler, P.J., A.J. Woakes, K. Smale, C.A. Roberts, C.J. Hillidge, D.H. Snow and D.J. Marlin. 1993. Respiratory and cardiovascular adjustments during exercise of increasing intensity and during recovery in thoroughbred racehorses. J. Exp. Biol 179:159-180.
- Deaton, C.M., D.J. Marlin, C.A. Roberts, N. Smith, P.A. Harris, F.J. Kelly and R.C. Schroter. 2002. Antioxidant supplementation and pulmonary function at rest and exercise. Equine Vet. J. 34:58- 65.
- Deaton, C.M., D.J. Marlin, N.C. Smith, C.A. Roberts, P.A. Harris, F.J. Kelly and R.C. Schroter. 2003. Pulmonary bioavailability of ascorbic acid in an ascorbate-synthesising species, the horse. Free Radical Res. 37:461-461.
- Derksen, F.J. 1997. Oxidant injury and nitric oxide: a role in exercise-induced pulmonary haemorrhage. Vet. J. 153:119-121.
- Dimock, A.N., P.D. Siciliano and C.W. McIlwraith. 2000. Evidence supporting an increased presence of reactive oxygen species in the diseased equine joint. Equine Vet. J. 32:439-443.
- Dunnett, C.E., D.J. Marlin and R.C. Harris. 2002. Effect of dietary lipid on response to exercise: relationship to metabolic adaptation. Equine Vet. J. 34:75-80.
- Dupont, J., P.J. White, M.P. Carpenter, E.J. Schaefer, S.N. Meydani, C.E. Elson, M. Woods, and S.L. Gorbach. 1990. Food uses and health effects of corn oil. J. Amer. Coll. Nutr. 9:438- 470.
- Evans, P. and B. Halliwell. 2001. Micronutrients: oxidant/antioxidant status. Br. J. Nutr. 85:S67-S74.
- Frape, D. 1998. The roles of major minerals and trace elements. In: Equine Nutrition and Feeding. Blackwell Science, Oxford, pp. 43-71.
- Gansen, S., A. Lindener and A. Wagener. 1995. Influence of a supplementation with natural and synthetic vitamin E on serum α-tocopherol content and v4 of Thoroughbred horses. Proc. of the 14th Equine Nutr. Physiol. Soc. Symp., p. 68.
- Goodman, S. 2003. Vitamin C: The master nutrient. www.positivehealth.com.
- Halliwell, B. 1994. Free radicals and antioxidants: A personal view. Nutr. Rev. 52:253-265.
- Halliwell, B. 2002. Latest developments in antioxidant research. Vitafoods International Conference. Vitafoods, Geneva, p. 2.
- Hargreaves, B.J., D.S. Kronfeld, J.N. Waldron, M.A. Lopes, L.S. Gay, K.E. Saker, W.L. Cooper, D.J. Sklan and P.A. Harris. 2002. Antioxidant status of horses during two 80-km endurance races. J. Nutr. 132:1781S-1783S.
- Hoffman, R.M., D.S. Kronfeld, J.L. Holland and K.M. Greiwe-Crandell. 1995. Preweaning diet and stall weaning method influences on stress response in foals. J. Anim. Sci. 73:2922-2930.
- Jariwalla, R.J. 2001. Rice-bran products: phytonutrients with potential applications in preventive and clinical medicine. Drugs and Exper. Clinic. Res. 27:17-26.
- Kirschvink, N., T. Art, B.D. Moffarts, N. Smith, D. Marlin, C. Roberts and P. Lekeux. 2002. Relationship between markers of blood oxidant status and physiological variables in healthy and heaves-affected horses after exercise. Equine Vet. J. 34:159-164.
- Lewis, L.D. 1995. Vitamins and minerals for horses. Equine Clinical Nutrition - Feeding and Care. Williams and Wilkins, Baltimore, pp. 25-88.
- Lloyd, B.J., T.J. Siebenmorgan and K.W. Beers. 2000. Effects of commercial processing on antioxidants in rice bran. Cereal Chem. 77:551- 555.
- Marlin, D.J., K. Fenn, N. Smith, C.D. Deaton, C.A. Roberts, P.A. Harris, C. Dunster and F.J. Kelly. 2002. Changes in circulatory antioxidant status in horses during prolonged exercise. J. Nutr. 132:1622S-1627S.
- McLaughlin, J.G. and J. Cullen. 1986. Clinical cases of chronic selenosis in horses. Irish Vet. J. 40:136- 138.
- McMeniman, N.P. and H.F. Hintz. 1992. Effect of vitamin E status on lipid peroxidation in exercised horses. Equine Vet. J. 24:482.
- Mills, P.C., N.C. Smith, I. Casas, P. Harris, R.C. Harris and D.J. Marlin. 1996. Effects of exercise intensity and environmental stress on indices of oxidative stress and iron homeostasis during exercise in the horse. Europ. J. Appl. Phy. 74:60- 66.
- Noguchi, N. and E. Niki. 1999. Chemistry of active oxygen species and antioxidants. In: Antioxidant Status, Diet, Nutrition and Health (A.M. Papas, ed). CRC Press, Boca Raton, pp. 3-20.
- NRC. 1989. Nutrient Requirements of Horses, 5th revised edition. National Academy Press, Washington DC.
- Pagan, J.D. 1999. Micromineral requirement in horses. In: Advances in Equine Nutrition II (J.D. Pagan and R.J. Geor, eds). Nottingham University Press, Kentucky, USA, pp. 317-327.
- Pagan, J.D., M.A.P. Kennedy, T. Currier and K.E. Hoekstra. 1999. Effects of selenium source on selenium digestibility and retention in exercised Thoroughbreds. Proceedings of the 6th Equine Nutrition and Physiology Symposium, North Carolina, pp. 135-140.
- Papas, A.M. 1999a. Diet and antioxidant status. In: Antioxidant Status, Diet, Nutrition and Health (A.M. Papas, ed). CRC Press, Boca Raton, pp. 89-106.
- Papas, A.M. 1999b. Vitamin E: Tocopherols and tocotrienols. In: Antioxidant Status, Diet, Nutrition and Health (A.M. Papas, ed). CRC Press, Boca Raton, pp. 189-210.
- Perkins, G., S.J. Valberg, J.M. Madigan, G.P. Carlson and S.L. Jones. 1998. Electrolyte disturbances in foals with severe rhabdomyolysis. J. Vet. Inter. Med. 12:173-177.
- Ralston, S.L. 1999. Management of geriatric horses. In: Advances in Equine Nutrition II (J.D. Pagan and R.J. Geor, eds). Nottingham University Press, Kentucky, USA, pp. 393-396.
- Rayman, M.P. 1997. Dietary selenium: time to act. Br. Med. J. 314:387-388.
- Rumsey, S.C., Y. Wang, and M. Levine. 1999. Vitamin C. In: Antioxidant Status, Diet, Nutrition and Health (A.M. Papas, ed). CRC Press, Boca Raton, pp. 160-187.
- Schrauzer, G.N. 2000. Selenomethionine: A review of its nutritional significance, metabolism and toxicity. J. Nutr. 130:1653-1656.
- Schryver, H.F. 1990. Mineral and vitamin intoxication in horses. Clinical Nutrition. W.B. Saunders Company, Philadelphia, PA, pp. 295-318.
- Shelle, J.E., W.D. Vanhuss, J.S. Rook and D.E. Ullrey. 1985. Relationship between selenium and vitamin E nutrition and exercise in horses. Proc. of the 9th Equine Nutrition and Physiology Symposium. EPNS, Michigan State University, pp. 104-109.
- Siciliano, P.D. and C.H. Wood. 1993. The effect of added dietary soybean oil on vitamin E status of the horse. Proc. 13th Equine Nutr. Physiol. Soc. Sym, p. 3.
- Snow, D.H. and M. Frigg. 1987. Plasma concentrations at monthly intervals of ascorbic acid, retinol, β-carotene and tocopherol in two Thoroughbred racing stables and the effects of supplementation. Proc. of the 10th Nutr. Phys. Symp., ENPS, Colorado State University, p. 617.
- Snow, D.H. and M. Frigg. 1989. Oral administration of different formulations of ascorbic acid to the horse. J. Equine Vet. Sci. 9:30-33.
- Snow, D.H. and M. Frigg. 1990. Bioavailability of ascorbic acid in horses. J. Vet. Pharm. and Ther. 13:393-403.
- Vitec. 1987. Omega 3 Fatty Acids. Roche Report, pp. A5, 1-5.
- Wardlaw, G.M. 1999. Perspectives in Nutrition, 4th Edition. WCB Mc Graw-Hill.
- White, A., M. Estrada, K. Walker, P. Wisnia, G. Filgueira , F. Valdes, O. Araneda, C. Behn and R. Martinez. 2001. Role of exercise and ascorbate on plasma antioxidant capacity in thoroughbred race horses. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 128:99-104.
- Wooden, G.R. and A.M. Papas. 1991. Utilisation of various forms of vitamin E by horses. Proc. 12th Equine Nutr. Physiol. Symp., p. 265.
Independent Equine Nutrition Ltd, Newmarket, UK

Heres related info on a antioxidant supplement specifically for nutrition of horses. a href[equal]http://www.sportforcellc.com/store/antioxidants-c-7/bioberry-potent-natural-anti-oxidant-formula-p-12 title[equal]Antioxidants for horsesBioBerry antioxidant supplement/a

I would like to thank Catherine Dunnett for her excellent article about antioxidants in physiology and nutrition of horses.
I wish to also bring across to the readers of this forum of the potential benefits of using oregano essential oil for its antioxidant properties in the equine species.
According to Joseph Mercola, whose findings were published in the Journal of Agricultural and Food Chemistry in November 2001, oregano was the herb with the highest antioxidant activity, with 3 to 20 times higher antioxidant activity than all the other herbs studied.
On a per gram fresh weight basis, oregano ranked even higher in antioxidant activity than fruits and vegetables which are known to be high in antioxidants. In comparison to the antioxidant activities of a few fruits and vegetables, oregano had 42 times more antioxidant activity than apples, 30 times more than potatoes, 12 times more than oranges and 4 times more than blueberries.
An example of an oregano essential oil product is Orego-Stim, produced by Meriden Animal Health Limited (UK). The phenolic compounds within such a product, carvacrol and thymol, are bioflavonoids that act as powerful antioxidants. These help combat free-radical damage, while protecting cells, lipids & vitamins such as A, C and E from the process of peroxidation.
An evaluation of antioxidant activities of different substances and the margin of deterioration of oxidation by UVA – VIS radiation showed that the antioxidant activity of oregano essential oil was higher than those of coriander and rosemary at various concentrations. In fact, it was even higher than ascorbic acid (vitamin C) and d-alpha tocopherol (vitamin E).
This shows that because carvacrol and thymol are more reactive, they will sacrifice themselves in the presence of a free radical in the peroxidation process, thus protecting the antioxidant vitamins and increasing their bioavailability as nutrients for the animals dietary use.


