Introduction
Animal domestication has been important in the development of human societies and today milk and dairy products are included in dietary recommendations by public health organizations around the world. Sustainability in agriculture depends in substantial part on the productivity of plant and animal agriculture. Productivity, also referred to as productive efficiency, is defined as the relationship between food production and resource inputs, and over the last century we have made remarkable improvements. Using the U.S. as an example, American farmers have gone from producing enough food to feed 15 people a century ago, to today where an average farmer produces sufficient food to feed over 150 people (Bauman and Capper, 2011). In the case of animal agriculture, the application of scientific principles in the U.S. dairy industry began following in the 1940s and annual milk yield per cow has increased from about 2100 kg in 1944 to over 10,000 kg today. Improvements in genetics and management, especially nutritional management, are the basis for these gains. In lactating cows, productive efficiency has been historically expressed as feed resource inputs per unit of milk output, and this is a measure of feed efficiency. Provision of feed represents a major cost in dairy production, and feed efficiency has improved over 400% during this interval. Thus, the dairy industry has made remarkable gains in the efficiency of resource use, and an equally impressive reduction in the environmental impact of producing milk (Capper et al., 2009; Capper and Bauman, 2013).
The dairy cow has a remarkable ability to coordinate her biological processes to support mammary gland use of nutrients for milk synthesis, a coordination that must involve all nutrient classes and include most, perhaps all, organs and physiological processes (Bauman and Currie, 1980). While a linear increase in milk yield per cow has occurred over the last century, at some point gains in milk yield will reach a plateau where genetic selection and management practices have optimized the biological processes essential for milk synthesis. But where is this plateau? In the Northeast we have many herds that annually average over 13,000 kg per cow. The top Jersey cow, Mainstream Barkley Jubilee, calved at 4 yr 8 mo and produced 25,270 kg of milk (4.6% fat, 3.2% protein). The Holstein record is held by a Wisconsin cow EverGreen View My 1326 with an annual production of 32,805 kg milk (3.9% fat, 3.0% protein); Evergreen had to have an average daily net energy intake which was over 6.5x maintenance. Finally, Guinness World Records recently recognized a Canadian cow, Gillette E Smurf, as the lifetime record holder; Smurf’s production of 217,350 kg milk in 10 lactations represents an amazing average of 38.9 kg of milk for every day of life. These few examples indicate dairy cows are capable of extraordinary milk yields. Indeed, there must be a plateau, but that plateau is not evident and increases in productive efficiency will clearly continue in the future.
The following sections identify the biological processes that are associated with the remarkable gains in productive efficiency that have occurred over the last century and discuss concepts of regulation in lactating dairy cows. Genetics and management are keys to these gains and this leads to a discussion of new applications of genomics, including biotechnology and recombinant bovine somatotropin, the recent focus on residual feed intake to examine the genomics of feed efficiency, and recognition of nutrigenomics and the role nutrients can play in regulating metabolism.
Biological Basis for Improvements in Productive Efficiency
Sources of Variation – Cycle of Life
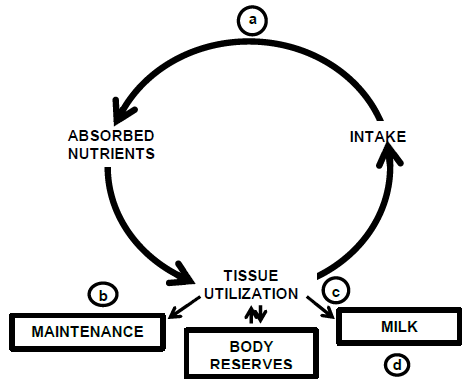
Productive efficiency can also be considered in biological terms. The biology of a dairy cow involves a series of chemical reactions in which food is transformed and used to support body tissues and activity. Feed is consumed and following rumen fermentation and digestive processes, nutrients are absorbed. These nutrients are then utilized by body tissues and this in turn plays a key role in regulating feed intake. Some nutrients are used to meet the maintenance requirement while other nutrients are used to maintain body reserves. Depending on physiological state, a major portion of the nutrients are utilized for productive functions such as milk synthesis (lactation) or fetal development (pregnancy). These biological processes comprise the “Cycle of Life” (Figure 1) and they provide the framework to consider the basis for the historic gains in productive efficiency (see review by Bauman et al., 1985).
Digestion and Nutrient Absorption. The chemical and physical characteristics of feedstuffs have major effects on digestibility, and this forms the basis for current extension recommendations. Likewise, effects of feeding level, diet composition and other dietary factors markedly alter digestibility and nutrient absorption (Huhtanen et al., 2009; Nousiainen et al., 2009). In contrast, most studies with lactating cows suggest little animal variation in apparent digestibility under standardized conditions (see review by Veerkamp and Emmans, 1995). Furthermore, genetic merit and digestibility were independent (Grieve et al., 1976; Custodio et al., 1983) and genetically diverse lines of dairy cows differed little in digestibility, especially when intake was similar (Davey et al., 1983; Belyea and Adams, 1990; Gordon et al., 1995; Ferris et al., 1999). Thus, management practices can markedly affect digestibility and nutrient absorption, but genetic advancements in this area have been of minor importance in the historic improvements in productive efficiency (Table 1).
Maintenance Requirement. The maintenance requirement can be markedly affected by diet, management and the environment. Dietary fiber content, grazing conditions, overcrowding, and heat stress serve as examples (see reviews by Agnew and Yan, 2000; Collier et al., 2005). However, relatively few studies have evaluated the variation associated with the maintenance requirement of dairy cows when compared under standardized conditions. van Es (1961) estimated that among animal coefficient of variation in maintenance requirement was 5 to 10% across 237 energy balance studies with dairy cows and steers. Flatt et al. (1969) also found remarkably good agreement in estimates of maintenance requirements of dairy cows among studies. Consistent with this Davey et al. (1983) concluded that the maintenance requirement was not influenced by genetic merit for milk production in Friesians. Thus, the maintenance requirement appears to be a minor source of variation among animals and changes in maintenance requirement have contributed little to the historic gains in productive efficiency (Table 1).
Utilization of Metabolizable Energy for Milk Production. The partial efficiency (PE) of metabolizable energy (ME) utilization varies according to use (maintenance, lactation, growth, fattening or pregnancy) and type of diet (Garrett and Johnson, 1983; Agnew and Yan, 2000). This variation undoubtedly relates to differences in the pattern of absorbed nutrients and the specific products being formed. However, under typical conditions in nutrient supply and milk composition there appears to be little difference among cows in the PE of milk synthesis (Table 1). Indeed, studies comparing low- and high-milk yielding genetic lines of dairy cows found no differences in the PE for milk synthesis (Belyea and Adams, 1990; Gordon et al., 1995; Ferris et al., 1999; Yan et al., 2006). The biochemical pathways for synthesis of major milk components involve a series of enzymatic reactions, each having a specific energetic efficiency. Thus, similarities in PE estimates are expected because all cows use the same biochemical pathways and enzyme reactions for the synthesis of milk components.
Nutrient Partitioning. The major source of historic gains in productive efficiency is nutrient partitioning (Table 1; Bauman et al., 1985). High-yielding cows direct a greater portion of absorbed nutrients to the mammary glands for milk synthesis and associated with this is a greater voluntary intake. Low-yielding cows have a lower voluntary intake: if they do consume more feed they use it for excessive body fat accretion rather than milk synthesis. Thus, selection for high milk production results in dairy cows that not only utilize more nutrients for milk synthesis but also have a higher intake to support a greater milk yield (Bauman et al., 1985; Agnew and Yan, 2000; Yan et al., 2006).
Productive efficiency is a key component of sustainability because a greater milk yield per cow results in less resource use per unit of milk, a concept often referred to as “dilution of maintenance” (Bauman et al., 1985). Dilution of maintenance is usually considered in terms of feed resources per unit of milk, but it applies more broadly to all of the costs of producing milk including renewable and non-renewable resources as well as the costs for facilities and labor.
Concepts of Regulation
The regulation of nutrient use for productive functions is a key biological principle in animal production with the overall goal of maintaining the animal’s well-being regardless of the physiological or environmental challenges that are encountered. The use of nutrients by the mammary gland represents a tremendous demand such that total maternal metabolism must be coordinated to support the demands for milk synthesis. From a conceptual basis, this involves two types of regulation, homeostasis and homeorhesis (Bauman and Currie, 1980; Bauman, 2010).
Homeostatic controls operate on an acute basis so that different tissues and organs are “working cooperatively” to maintain physiological equilibrium. There are many well-established examples of the multiple compensatory mechanisms functioning to maintain physiological equilibrium despite challenges from the external environment. One important example in dairy cows is the maintenance of circulating glucose. Glucose supply is critical for many tissues so over the short term the reciprocal actions of insulin and glucagon ensure the proper balance in glucose supply and for use in milk synthesis and other processes (Bauman and Currie, 1980).
Homeorhesis was defined as the “orchestrated changes for priorities of a physiological state” (Bauman and Currie, 1980). This was first applied to lactation and pregnancy, but the general concept has been extended to include different physiological states, nutritional and environmental situations, and even pathological conditions (Collier et al., 2005; Bauman, 2010). Key features of homeorhetic control are: 1) its chronic nature, hours and days vs. seconds and minutes required for most examples of homeostatic regulation; 2) its simultaneous influence on multiple tissues and systems that results in an overall coordinated response, and; 3) its mediation through altered response to homeostatic signals (Bauman, 2010). Bovine somatotropin is arguable the best characterized example of a homeorhetic control, and its regulation of nutrient partitioning has a remarkable impact on productive efficiency (Bauman, 1999).
Application of Genomics to Nutrition
Genomic Developments
In April 2009, SCIENCE heralded new developments in animal agriculture with its issue cover and two articles announcing the sequencing of the bovine genome (The Bovine Genome Sequencing and Analysis Consortium et al., 2009; The Bovine HapMap Consortium, 2009). The bovine genome is similar in size to the genomes of humans and other mammals, containing about 3 billion base pairs and ~ 22,000 genes. These genes code for the specific proteins involved in the cycle of life discussed earlier. Thus, sequencing of the bovine genome and the development of new genomic tools offer an opportunity to better understand the biology of dairy cattle and provide the framework to identify the genetic basis for animal differences in productive efficiency that can be applied to improve performance and animal well-being by genetic selection or management intervention (Garrick et al., 2012).
Single nucleotide polymorphisms (SNPs) represent nucleotide base changes within the DNA sequence. Recently, USDA-ARS scientists genotyped over 5000 Holstein cows and bulls; 38,000 SNP markers were identified which they related to selection traits such as milk production and longevity (VanRaden et al., 2009; Cole et al., 2009). SNPs will undoubtedly be associated with the greater productivity efficiency observed for high producing dairy cows. However, it seems likely that small differences in variation may exist at many sites rather than few sites capturing large portions of the variation in productive efficiency among animals.
Each gene codes for a particular sequence of amino acids that constitute a specific protein. A SNP can result in a different amino acid in the protein sequence and this may affect the biological function of the protein. At this point in time there has been only limited success in identifying specific SNPs that correspond to functional differences the productive efficiency (feed efficiency) of dairy cows, but this area should offer exciting applications in the future.
There are three areas where the developments in genomics are already affecting our understanding related to nutrition, feed efficiency and the biology of milk production. These are 1) the application of recombinant DNA technology to produce bovine somatotropin, 2) the use of residual feed intake to identify cows with superior genetics for feed efficiency, and 3) the application of nutrigenomics to understand and trouble shoot problems in milk fat synthesis. Discoveries in biotechnology, specifically recombinant DNA technologies, were key to the development and commercialization of recombinant bovine somatotropin (rbST) as a management tool for dairy producers; to date more than 35 million U.S. dairy cows have received rbST supplements over its 19 years of commercial use. rbST is a homeorhetic control that dramatically effects nutrient partitioning resulting in an increase in milk yield of 4 to 5 kg/day. Mechanisms by which rbST orchestrates metabolism to improve feed efficiency and reduce the carbon footprint of milk production have been reviewed elsewhere (Bauman, 1999; Capper and Bauman, 2013) so will not be discussed further. The other two areas represent more recent developments using genomics, and they will be briefly discussed in the following sections.
Residual Feed Intake
Feed efficiency is defined and measured in several different ways. One of these is residual feed intake (RFI), first purposed by Koch et al. (1963) as a measure of feed efficiency in growing cattle. In beef cattle, the cited advantage of RFI is that it is phenotypically independent of the production traits used to estimate feed intake (Herd and Arthur, 2009; Berry and Crowley, 2012). The RFI for growing cattle is calculated as the difference between an animal’s actual feed intake and its expected feed intake, which was determined by regression of dry matter intake against mean body weight and growth rate (Williams et al., 2011). Recently, several groups have estimated RFI values to evaluate the feed efficiency of lactating dairy cows (Coleman et al., 2010; Connor et al., 2012; VandeHaar et al., 2012). Prediction of feed intake for lactating cows varies, but generally involves a regression that adjusts for parity, body size, changes in body weight and body condition score, and production of milk and milk components.
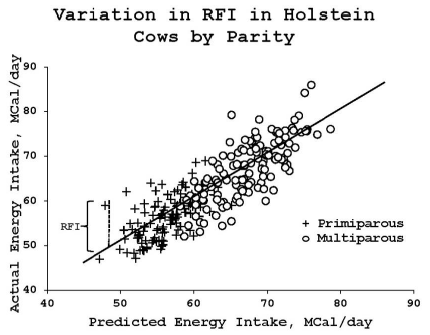
An example of the variation in RFI for a group of Holstein cows in early lactation is shown in Figure 2. Cows that lie below the unity line have a greater feed efficiency because their actual feed intake is less than the predicted feed intake as derived from the regression adjustments. A potential advantage of RFI is the possibility of identifying dairy cows that inherently have a high feed efficiency, but do not exhibit the negative effects on health risk or well-being that are often associated with high milk production (Connor et al., 2012). Thus, RFI could be a tool to identify more efficient animals and derive associations between SNPs and RFI for use in a selection program. However, in a review on genetics of feed efficiency of dairy and beef cattle Berry and Crowley (2013) cautioned that it was a misconception to assume the unexplained intake (RFI) represents true feed efficiency; they joined with Robinson (2005) and pointed out that the residual component can be due, in part, to random noise, such as measurement and prediction error, or due to inaccurate recording, feed losses, or bias in the regression coefficients for the respective regressors.
As defined earlier, productivity or productive efficiency in dairy cows is calculated as milk per unit of resource input, so it also represents a measure of feed efficiency. How does the earlier discussion on the “cycle of life” and sources of variation in productive efficiency relate to the variation depicted by RFI values? The calculation of RFI is designed to remove animal differences in nutrient partitioning via the adjustments related to individual animal differences in milk yield and body weight and body condition. Thus, animal differences in nutrient partitioning, the area identified in the preceding section as is not a component of the RFI evaluation. However, RFI would include animal differences in digestibility and nutrient absorption, maintenance requirement and the partial efficiency of nutrient use for milk synthesis. Indeed, in selection based on RFI, cows identified as having the best feed efficiency (as reflected by a lower RFI) can be at any level of milk production. Thus, the impressive gains in feed efficiency that are a consequence of achieving higher milk yields and the “dilution of maintenance” in gains in feed efficiency are not components of RFI. The application of RFI to lactating dairy cows is a recent concept and thus far there is a paucity of data to evaluate this approach. However, the failure of RFI to capture the cow differences in milk yield and the dilution of maintenance represents an important limitation; as discussed earlier; nutrient partitioning represents the major source for historical gains in productive efficiency and the major source of variation among today’s top herds and genetically superior cows (Table 1). Of additional interest is to evaluate the extent to which RFI is repeatable; variables used in the regression adjustments are all associated with some degree of error so a demonstration that the rank order of RFI among cows is repeatable in successive lactations would provide solid support to its potential for use in selection systems.
Nutrigenomics
Nutrigenomics is an umbrella term that refers to the impact of dietary components on physiological process by altering gene expression, epigenetic effects, proteins or metabolites (Bauman et al., 2011). Nutrition research has characteristically centered on identifying essential nutrients, establishing dietary requirements, and developing feeding systems to meet these requirements. Thus, past nutrition research has focused on designing diets that supply a quantity and pattern of nutrients to provide nourishment and allow for normal body function and health maintenance. Nutrigenomics represents a new dimension in nutrition research; results provide clear evidence that in addition to a nutritive role, nutrients can also regulate the expression of genes thereby affecting metabolic pathways and homeostatic control, including diet-environment interactions. Nutrigenomics has been most vigorously studied in humans where it is hypothesized that the diversity in human genotypes provides differences that may allow an optimization of diets on an individual basis to maintain health and prevent chronic diseases (Müller and Kersten, 2003).
Among the best characterized examples of nutrigenomics is diet-induced milk fat depression (MFD) in dairy cows (Bauman et al., 2011). While diet-induced MFD continues to be a challenge for commercial dairies, in the context of this paper it serves to illustrate how absorbed nutrients can effect gene expression and alter metabolic processes. The problem of diet-induced MFD perplexed producers, consultants and scientists for over a half-century, and many theories and practices were purposed and found inadequate (Harvatine et al., 2009). A key development in understanding MFD was the recognition that it involved an interaction between rumen fermentation and mammary fatty acid (FA) synthesis. Specifically, certain dietary conditions shift rumen fermentation and the bacterial pathways for the biohydrogenation of dietary polyunsaturated FAs. As a result unique conjugated FAs are produced as biohydrogenation intermediates and some of these are potent inhibitors of milk fat synthesis. To date three conjugated linoleic acid (CLA) isomers have been identify as bioactive FAs that inhibit milk fat synthesis – trans-10, cis-12 CLA, cis-10, trans-12 CLA and trans-9, cis-11 CLA. This inhibition involves a coordinated down regulation of gene expression for key enzymes in the synthesis of milk fat. This has been best studied for trans-10, cis-12 CLA and the cellular mechanism involves the SREBP1 transcription factor family. The genes for key enzymes in FA synthesis have a base sequence in their DNA code that is referred to as a SRE element. The SRE element binds the active fragment of SREBP1 thereby reducing gene expression (Bauman et al., 2011). Table 2 lists the specific lipogenic genes that are coordinately down-regulated during diet-induced MFD and CLA-induced MFD, and the DNA sequence for all of these genes has a SRE element in the promoter region.
In the case of MFD, an understanding of how unique rumen biohydrogenation intermediates are able to regulate milk fat synthesis is already being applied in the formulation of diets to minimize the problem and for troubleshooting MFD when problems are encountered on commercial dairy farms. In particular this example provides insight to the potential importance of nutrient-gene interactions and the area of nutrigenomics. Clearly, improving our understanding of the role that specific nutrients play in the regulation of gene expression and metabolism represents a developing area that should offer exciting new opportunities to improve productive efficiency of dairy cows.
Final Thoughts
Our challenge in animal agriculture is to understand the regulation of physiological processes and their role in animal productivity, well-being and disease prevention. Estimates are that about one-half to two-thirds of the historic gains in productive efficiency have come from genetic improvement and the remainder from advances in nutrition and management. Presentations at this and previous Symposiums in this series provide invaluable information on the latest developments in nutrition, management, and the biology of lactation. But it’s important to recognize that this is a package deal – sound nutrition and management practices are essential to fully actualize improvements in genetics and vice versa. While past gains have been impressive, recent developments in genomics and nutrition indicate that the opportunity for future gains in productive efficiency may be even greater.
References
Agnew, R.E., and T. Yan. 2000. Impact of recent research on energy feeding systems for dairy cattle. Livest. Prod. Sci. 66:197-215.
Bauman, D.E. 1999. Bovine somatotropin and lactation: from basic science to commercial application. Domes. Anim. Endo. 17:101-116.
Bauman, D.E. 2010. Homeorhesis: past, present and future. 2010 Elanco Science Symposium, pp 1-20.
Bauman, D.E., and W.B. Currie. 1980. Partitioning of nutrients during pregnancy and lactation: a review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 63:1514-1529.
Bauman, D.E., and J.L. Capper. 2011. Sustainability and dairy production: challenges and opportunities. Proc. Cornell Nutr. Conf., pp. 136-153.
Bauman, D.E., K.J. Harvatine, and A.L. Lock. 2011. Nutrigenomics, rumen-derived bioactive fatty acids and the regulation of milk fat synthesis. Annu. Rev. Nutr. 31:299-319.
Bauman, D.E., S.N. McCutcheon, W.D. Steinhour, P.J. Eppard, and S.J. Sechen. 1985. Sources of variation and prospects for improvement of productive efficiency in the dairy cow: a review. J. Anim. Sci. 60:583-592.
Belyea, R.L., and M.W. Adams. 1990. Energy and nitrogen utilization of high versus low producing dairy cows. J. Dairy Sci. 73:1023-1030.
Berry, D.P., and J.J. Crowley. 2012. Residual intake and body weight gain: a new measure of efficiency in growing cattle. J. Anim. Sci. 90:109-115.
Berry, D.P., and J.J. Crowley. 2013. Genetics of feed efficiency in dairy and beef cattle. J. Anim. Sci. (published ahead of print January 23, 2013) doi:10.2527/jas.2012-5862.
Capper, J.L., and D.E. Bauman. 2013. The role of productivity in improving the environmental sustainability of ruminant production systems. Annu. Rev. Anim. Biosci. 1:9.1-9.21.
Capper, J.L., R.A. Cady, and D.E. Bauman. 2009. The environmental impact of dairy production: 1994 compared with 2007. J. Anim. Sci. 87:2160-2167.
Cole, J.B., P.M. VanRaden, J.R. O’Connell, C.P. Van Tassell, T.S. Sonstegard, R.D. Schnabel, J.F. Taylor, and G.R. Wiggans. 2009. Distribution and location of genetic effects for dairy traits. J. Dairy Sci. 92:2931-2946.
Coleman, J., D.P. Berry, K.M. Pierce, A. Brennan, and B. Horan. 2010. Dry matter intake and feed efficiency profiles of 3 genotypes of Holstein-Friesian within pasture-based systems of milk production. J. Dairy Sci. 93:4318-4331.
Collier, R.J., L.H. Baumgard, A.L. Lock, and D.E. Bauman. 2005. Physiological limitations, nutrient partitioning. Pages 351-378 in Yields of Farmed Species: Constraints and Opportunities in the 21st Century. R. Sylvester-Bradley and J. Wiseman eds. Nottingham University Press, Nottingham, UK.
Connor, E.E., J.L. Hutchison, and H.D. Norman. 2012. Estimating feed efficiency of lactating dairy cattle using residual feed intake. Pages 159-173 in Feed Efficiency in the Beef Industry. R.A Hill, ed. . John Wiley & Sons, Inc., Somerset, NJ.
Connor, E.E., J.L. Hutchinson, K.M. Olson, and H.D. Norman. 2012. Triennial Lactation Symposium: opportunities for improving milk production efficiency in dairy cattle. J. Anim. Sci. 90:1687-1694.
Custodio, A.A., R.W. Blake, P.F. Dahm, T.C. Cartwright, G.T. Schelling, and C.E. Coppock. 1983. Relationships between measures of feed efficiency and transmitting ability for milk of Holstein cows. J. Dairy Sci. 66:1937.
Davey, A.W.F., C. Grainger, D.D.S. Mackenzie, D.S. Flux, G.F. Wilson, I.M. Brookes, and C.W. Holmes. 1983. Nutritional and physiological studies of differences between Friesian cows of high or low genetic merit. Proc. New Zealand Soc. Anim. Prod. 43:67.
Ferris, C.P., F.J. Gordon, D.C. Patterson, M.G. Porter, T. Yan. 1999. The effect of genetic merit and concentrate proportion in the diet on nutrient utilisation by lactating dairy cows. J. Ag. Sci., Cambridge 132:483-490.
Flatt, W.P., P.W. Moe, and L.A. Moore. 1969. Influence of pregnancy and ration composition on energy utilization by dairy cows. Pages 123-138 in Energy Metabolism of Farm Animals. K.L. Blaxter, J. Kielanowski, and G. Thorbek eds. Assoc. Anim. Prod. Publ. No. 12. Oriel Press, Newcastle upon Tyne, England.
Garrett, W.N., and D.E. Johnson. 1983. Nutritional energetic of ruminants. J. Anim. Sci. 57(Suppl. 2):478.
Garrick, D.J., L.H. Baumgard, and H.L. Neilbergs. 2012. Invited Review: genomic analysis of data from physiological studies. J. Dairy Sci. 95:499-507.
Gordon, F.J., D.C. Patterson, T.Yan, M.G. Porter, C.S. Mayne, and E.F. Unsworth. 1995. The influence of genetic index for milk production on the response to complete diet feeding and the utilization of energy and nitrogen. Anim. Sci. 61:199-210.
Grieve, D.G., G.K. MacLeod, T.R. Batra, E.B. Burnside, and J.B. Stone. 1976. Relationship of feed intake and ration digestibility to estimated transmitting ability, body weight and efficiency in first lactation. J. Dairy Sci. 59:1312.
Harvatine, K.J., Y.R. Boisclair, and D.E. Bauman. 2009. Recent advances in the regulation of milk fat synthesis. Animal 3:40-54.
Herd, R.M., and P.F. Arthur. 2009. Physiological basis for residual feed intake. J. Anim. Sci. 87(E. Suppl.):E64-E71.
Huhtanen, P., M. Rinne, and J. Nousiainen. 2009. A meta-analysis of feed digestion in dairy cows. 2. The effects of feeding level and diet composition on digestibility. J. Dairy Sci. 92:5031-5042.
Koch, R.M., L.A. Swiger, D. Chambers, and K.E. Gregory. 1963. Efficiency of feed use in beef cattle. J. Anim. Sci. 22:486-494.
Müller, M., and S. Kersten. 2003. Nutrigenomics: goals and strategies. Nature Reviews. Genetics 1:315.
Nousiainen, J., M. Rinne, and P. Huhtanen. 2009. A meta-analysis of feed digestion in dairy cows. 1. The effects of forage and concentrate factors on total diet digestibility. J. Dairy Sci. 92:5019-5030.
Reid, J.T. 1974. Energy metabolism in the whole animal. Pages 113-151 in The control of metabolism. J.D. Sink, ed. The Pennsylvania State Univ. Press, University Park, PA.
Robinson, D.L. 2005. Accounting for bias in regression coeficients with example from feed efficiency. Livest. Prod. Sci. 95:155-166.
The Bovine Genome Sequencing and Analysis Consortium, C. E. Elski, R. L. Tellam, and K. C. Worley. 2009. The genome sequence of Taurine cattle: a window to ruminant biology and evolution. Science 324:522-528.
The Bovine HapMap Consortium. 2009. Genome-wide survey of SNP variation uncovers the genetic structure of cattle breeds. Science 324:528-532.
van Es, A.J.H. 1961. Between-animal variation in the amount of energy required for the maintenance of dairy cows. Ph.D. Dissertation. State Agricultural Univ., Wageningen, The Netherlands.
VandeHaar, M.J., D.M. Spurlock, L. Armentano, R. Tempelman, K. Weigel, and R. Veerkamp. 2012. Considerations in using residual feed intake to define feed efficiency in dairy cattle. J. Anim. Sci. 90(E. Suppl.1). Abstract T44.
VanRaden, P.M., C.P. Van Tassell, G.R. Wiggans, T.S. Sonstegard, R.D. Schnabel, J.F. Taylor, and F.S. Schenkel. 2009. Invited review: reliability of genomic predictions for North American Holstein bulls. J. Dairy Sci. 92:16-24.
Veerkamp, R.F., and G.C. Emmans. 1995. Sources of variation in energetic efficiency of dairy cows. Livest. Prod. Sci. 44:87-97.
Williams, Y.J., J.E. Pryce, C. Grainger, W.J. Wales, N. Linden, M. Porker, and B.J. Hayes. 2011. Variation in residual feed intake in Holstein-Friesian dairy heifers in southern Australia. J. Dairy Sci. 94:4715-4725.
Yan, T., C.S. Mayne, T.W.J. Keady, and R.E. Agnew. 2006. Effects of dairy cow genotype with two planes of nutrition on energy partitioning between milk and body tissue. J. Dairy Sci. 89:1031-1942.
Table 1. Sources of variation in productive efficiency (feed efficiency) in lactating dairy cows1
Table 2. Summary of SREBP1-regulated lipogenic genes in which mammary expression is coordinately reduced during milk fat depression1
Figure 1. Basis for animal differences in productive efficiency as illustrated by the Cycle of Life. Biological processes are labeled as: a) digestion and absorption, b) maintenance requirement, c) partial efficiency of milk synthesis, and d) nutrient partitioning for milk synthesis. Adapted from Bauman and Currie (1980).
Figure 2. Predicted energy intake vs actual intake of Holstein cows (254 lactations) during the first 90 days postpartum. Solid line represents equality between predicted and actual intake. The distance between the line and each data point is the residual feed intake (RFI) for that individual lactation. From Connor et al. (2012); reproduced with permission of John Wiley & Sons, Inc.
This paper was presented at the The 3rd International Symposium on Dairy Cow Nutrition and Milk Quality, Beijing, May 2013.